
Abstract
Background:
Males and females who consume cannabis can experience different mental health and cognitive problems. Neuroscientific theories of addiction postulate that dependence is underscored by neuroadaptations, but do not account for the contribution of distinct sexes. Further, there is little evidence for sex differences in the neurobiology of cannabis dependence as most neuroimaging studies have been conducted in largely male samples in which cannabis dependence, as opposed to use, is often not ascertained.
Methods:
We examined subregional hippocampus and amygdala volumetry in a sample of 206 people recruited from the ENIGMA Addiction Working Group. They included 59 people with cannabis dependence (17 females), 49 cannabis users without cannabis dependence (20 females), and 98 controls (33 females).
Results:
We found no group-by-sex effect on subregional volumetry. The left hippocampal cornu ammonis subfield 1 (CA1) volumes were lower in dependent cannabis users compared with non-dependent cannabis users (p<0.001, d=0.32) and with controls (p=0.022, d=0.18). Further, the left cornu ammonis subfield 3 (CA3) and left dentate gyrus volumes were lower in dependent versus non-dependent cannabis users but not versus controls (p=0.002, d=0.37, and p=0.002, d=0.31, respectively). All models controlled for age, intelligence quotient (IQ), alcohol and tobacco use, and intracranial volume. Amygdala volumetry was not affected by group or group-by-sex, but was smaller in females than males.
Conclusions:
Our findings suggest that the relationship between cannabis dependence and subregional volumetry was not moderated by sex. Specifically, dependent (rather than non-dependent) cannabis use may be associated with alterations in selected hippocampus subfields high in cannabinoid type 1 (CB1) receptors and implicated in addictive behavior. As these data are cross-sectional, it is plausible that differences predate cannabis dependence onset and contribute to the initiation of cannabis dependence. Longitudinal neuroimaging work is required to examine the time-course of the onset of subregional hippocampal alterations in cannabis dependence, and their progression as cannabis dependence exacerbates or recovers over time.
Introduction
Cannabis products are widely used globally with current estimates of ∼284 million users annually—a 26% increase over the previous decade. 1 An emerging public health concern with increasing availability of high potency cannabis is the risk for developing cannabis dependence or cannabis use disorder (CUD)—broadly defined as an inability to limit/control use despite use causing physical or psychological harm, 2 as per DSM-IV 3 and DSM-5, 4 respectively.
While the majority of “ever users” will experience little impact of their use on their daily functioning, 5 the United Nations World Drug Report 2020 estimates that approximately 9.9% of previous year users were daily or near daily users. Although numbers of daily or near daily users are not necessarily indicative of CUD, recent estimates suggest that around 1 in 10 regular users, and 1 in 3 daily users do meet the criteria for CUD. 6 Accordingly, there remains a lack of consensus regarding whether levels of cannabis use and presence of a CUD might differentially influence health outcomes. 7
Sex differences have been reported previously for several aspects of cannabis use, including risk of dependence. For example, males comprise around 75% of all cannabis users, 8 although the number of females who use cannabis has been increasing internationally.9–11 Further, while CUD is more prevalent in males (3.5%) than females (1.7%),12,13 females can experience a greater severity of cannabis dependence and withdrawal,12,14 undergo a more rapid progression from first cannabis use to cannabis dependence (∼13 years for females and ∼18 years for males), and have greater incidence of relapse than males.12,14–19 These differences may be (in part) ascribed to distinct neurobiological changes in males and females who use cannabis and experience problems with cannabis use.20,21
Various structural neuroimaging studies have reported lower hippocampal and amygdala volumes in cannabis users compared with controls. 22 Our 2019 meta-analysis of 12 studies/15 samples found small differences in total hippocampal volume. 23 More recent empirical work observed small effects on total hippocampal and subfield volumes, 24 and widespread alterations in hippocampal and amygdala subfields detected using multivariate morphometry statistics. 25 However, the presence of cannabis dependence/CUD was not a consistent inclusion criterion throughout the studies.
The hippocampus is involved in learning, memory, and stress,26,27 each of which has been reported to be altered in cannabis users;28–35 the amygdala may also have a role in cannabis craving and withdrawal.36–38 Thus, both brain regions have been proposed to be implicated in the development of cannabis dependence.
However, lower hippocampal and amygdala volumes are not always observed in the literature on cannabis users. 39 Inconsistent findings across studies could be driven by several methodological limitations. For example, confounders known to affect volumetry—such as, age, intracranial volume (ICV), alcohol and tobacco exposure, and intelligence quotient (IQ)—are often not accounted for and may make a distinct contribution to the variance in regional volumes in each study.
Despite sex differences in neuroanatomy, 40 and putative differences in rapidity of progression to cannabis dependence, most neuroimaging studies evaluate differing proportions of male/female participants, and often do not examine how sex interacts with cannabis use and neuroanatomy. 22 Thus, differences in numbers of males/females across studies may underscore some of the variations in results. In addition, the severity of cannabis dependence is not assessed in most samples examined to date.
This is important, as cannabis dependence, rather than non-dependent, recreational/occasional use, may be more likely associated with neuroadaptations within brain regions related to stress, reward, and learning/memory, as postulated within prominent neuroscientific theories of addiction. 37 This distinction has been observed in our prior work that showed lower hippocampal volumes in cannabis users with versus without dependence (small effects; Cohen's d's 0.28–0.39).41–43
Therefore, the neuroimaging evidence in cannabis users requires further examination of sex differences, and dependent versus non-dependent use. In addition, alongside difficulties with the accurate measurement of cannabis use indices, exposure has been seldom measured and examined in relation to volumetry,23,44,45 and correlational work is required to explore whether volumetric differences may be partially driven by chronicity of use.
The research on hippocampal and amygdala volumes to date has largely examined the volume of the whole hippocampus and amygdala and not their subregional neuroanatomy. Importantly, distinct hippocampus and amygdala subregions have a different distribution of CB1 receptors 46 and are implicated in distinct aspects of addictive behavior. For example, CB1 receptors are densely innervated within the molecular layer of the dentate gyrus, the strata pyramidale of the CA2 and cornu ammonis subfield 3 (CA3), the subiculum,46–48 and in the basal nucleus of the amygdala. 48
CB1 receptors may drive volumetric changes detected in cannabis users, as they are targeted by cannabis main psychoactive compound delta-9-tetrahydrocannabinol (THC) 49 and may therefore be vulnerable to repeated cannabinoid exposure. Further, emerging evidence shows altered neuroanatomy of selected hippocampal subregions in human cannabis users42,43,50,51 and in animals. 52
Selected hippocampal subfields and amygdala nuclei are implicated in specific aspects of addictive behavior. For instance, chronic stress associated with substance use may disrupt neurogenesis in the hippocampal dentate gyrus. 53 Meanwhile, cornu ammonis subfield 1 (CA1) and CA3 neurons are implicated in learning, memory, and stress,54–56 and the impairment of these cognitive domains has been observed in cannabis users.28,57 Further, the central amygdala is implicated in negative affect, stress, and withdrawal in substance dependence. 48
In sum, no study has yet concurrently examined how cannabis dependence status and sex affect subregional neuroanatomy within the hippocampus and amygdala in cannabis users, which might provide mechanistic insight into cannabis use and dependence.
We aimed to examine the interactive and independent roles of sex, and cannabis dependence on the subregional neuroanatomy of the hippocampus, and amygdala. Based on published literature in humans and animals, the hippocampal subfields selected were the dentate gyrus, CA1, CA3, and subiculum,42,43,46,47,50–52,58 and the amygdala subregions included the basolateral and central amygdala.46,48 All models included age, ICV, IQ, monthly standard drinks, and monthly cigarettes as covariates.
Based on our previous work,43,58 we hypothesized that there would be a main effect of cannabis user status whereby dependent cannabis users (relative to non-dependent cannabis users and controls) show volumetric reductions in selected hippocampal subfields (i.e., dentate gyrus, CA1, CA3 and subiculum) and amygdala nuclei (i.e., central and basolateral) high in CB1 receptors and implicated in addictive behavior.
We also hypothesized that there would be a sex-by-group interaction suggestive of volumetric alterations being moderated by sex. Further, we examined whether any regions showing group or group-by-sex effects were predicted by cannabis exposure parameters, above and beyond age, ICV, sex, monthly standard drinks, and monthly cigarettes.
Methods
This study was preregistered in an open access platform: https://osf.io/t8f9m Magnetic resonance imaging (MRI) and behavioral data were collected from seven research sites of the ENIGMA Addiction Working Group (http://enigma.ini.usc.edu/ongoing/enigma-addiction-working-group).
Participants
The original sample comprised 451 participants, who were screened based on our study exclusion criteria: current psychiatric comorbidities; no current regular use; missing data for any key variables (i.e., IQ and age); major MRI artefacts; lack of consistent data on substance use, such as cannabis (age of onset in years, cannabis quantity per month), standard drinks, and cigarettes per month; lack of assessment of cannabis dependence. All sites obtained approval from local ethics committees and obtained written informed consent from all participants at their local institution.
Measures
Demographic, substance use, and MRI acquisition/analysis tool characteristics were obtained from three research sites through multiple instruments (Supplementary Table S1 in the Supplementary Data). 59 We measured age, sex (male/female), IQ, and metrics for cannabis (i.e., age of cannabis use onset, current monthly cannabis dosage), alcohol (i.e., standard drinks/month, occasions of alcohol use, or drinking days), and tobacco use (i.e., cigarettes smoked or smoking occasions per month).
Monthly cannabis dosage (i.e., joints, bongs, grams) was converted to a standardized unit for comparisons (e.g., 1 g=12 cones, 1 paper joint=3 cones; as per https://cannabissupport.com.au/media/1593/timeline-followback.pdf). Criteria defining dependent/non-dependent cannabis use differed slightly by site. As performed in previous epidemiological research, 60 sites 1, 2, and 3 used the severity of dependence scale (SDS) total score of 4 or more out of 15. See Supplementary Table S1 in the Supplementary Data for further details.
Structural MRI data acquisition and processing
MRI data acquisition and processing
Structural T1-weighted brain MRI scans were analyzed with a standardized protocol (Supplementary Table S1 in the Supplementary Data). They were processed at the individual research sites using the fully automated neuroimaging segmentation algorithms61,62 within FreeSurfer version 5.3 (https://surfer.nmr.mgh.harvard.edu) and standardized protocols to harmonize analysis and quality control (http://enigma.ini.usc.edu/protocols/imaging-protocols).
Segmentation of the hippocampus and amygdala
Volumes for the hippocampal subfields and amygdala nuclei were obtained using the automated segmentation algorithm available in FreeSurfer 6.0.63,64 Importantly, FreeSurfer has been shown to have high test-retest reliability for automated small subregion segmentation both within and across different scanners. 65 We extracted volumes for the hippocampus and its subfields of interest: subiculum, CA1, CA3, GC-ML-DG (granule cell and molecular layer of the dentate gyrus); and for the total amygdala and its nuclei of interest: lateral, basal, and central nuclei.
The volumes for the hippocampus dentate gyrus and for the basolateral amygdala were obtained by summing up those of the smaller subregional outputs from FreeSurfer (i.e., hippocampus GC-ML-dentate gyrus and molecular layer; amygdala basal and lateral nuclei). Additional visual inspection was performed locally at Australian Catholic University (see Acknowledgments section) on all participants to ensure quality of the automated segmentation algorithm outputs.
Statistical analyses
Sample characteristics were used to fit linear mixed-effect models, to examine group (dependent cannabis users, non-dependent cannabis users, and controls), sex (females and males) and group-by-sex effects on demographics, substance use patterns, and ICV. To address the primary aim, we implemented linear mixed-effect models using group (dependent cannabis users, non-dependent cannabis users, and controls), sex (male, female), and group-by-sex as predictors, and subregional volumes as dependent variables.
All models were adjusted for age, IQ, ICV, monthly standard drinks, and monthly cigarettes. In all models, site was treated as a random factor and estimated as an intraclass correlation (ICC) to account for the systematic site-level variation in the dependent variables expected to occur from differences in scanners, protocols, and assessments.
Post hoc linear mixed models were estimated to examine how subregional volumes that were significantly affected by group were predicted by cannabis use parameters (i.e., age of onset of cannabis use, monthly cannabis dosage in cones). Subregional volumes were residualised for sex, age, IQ, ICV, monthly standard drinks, and monthly cigarettes before being entered in the model.
For all volumetric analyses, we used Cohen's d to estimate the size of significant group and group-by-sex effects, from estimated marginal means. Benjamini-Hochberg false discovery rate (FDR) correction 66 was performed via pooling all model parameters (i.e., sex, group, group×sex, age, IQ, alcohol, nicotine, ICV for all hippocampal or amygdala subregions) for simultaneous p-value adjustment. Statistical analyses were conducted using IBM SPSS Statistics for Mac Version 29.0.
Results
Sample demographics and substance use characteristics
Table 1 summarizes demographic and substance use characteristics for the sample of 206 participants, of whom 146 were male and 70 were female. Participants were recruited from three sites as part of the ENIGMA Addiction Working Group. They included 59 people with cannabis dependence (17 females), 49 cannabis users without cannabis dependence (20 females), and 98 controls (33 females).
Demographic, Substance Use Characteristics, and Brain Volumes, in Male and Female Controls and Cannabis Users
Values for age, education, IQ, ICV, StDr/monthly, Cig/monthly are mean (SD). Significant findings (p<0.05) are shown in bold. Cones=standardized measure of cannabis dosage (https://cannabissupport.com.au/media/1593/timeline-followback.pdf). Var=site-level variation estimated as an ICC.
Chi-squared test revealed no group differences in sex composition: χ 2 (2, 206)=1.73, p=0.422.
IQ=estimated with the DART (site 1) 99 ; the vocabulary subscale of the WAIS-III (site 2); and the WASI (site 3) 100 .
Cig, smoked cigarettes; DART, Dutch version of the National Adult Reading Test; Dep, dependent cannabis users; F, females; fDep, female dependent cannabis users; fHC, female controls; fNDep, female non-dependent cannabis users; HC, controls; ICC, intraclass correlation; ICV, intracranial volume; IQ, intelligence quotient; M, males; NDep, non-dependent cannabis users; SD, standard deviation; StDr, standard drinks; Var, variation; WAIS-III, Wechsler Adult Intelligence Scale—Third Edition; WASI, Wechsler Abbreviated Scale of Intelligence.
The groups (dependent cannabis user, non-dependent cannabis user, and controls) did not differ in age; there were no sex differences in the age distributions, and no group-by-sex interaction. User groups did not differ in their weekly alcohol use, but there was an effect of sex—whereby males across the sample drank more than females (mean difference=13.8 drinks, p=0.004); even so, there was no group-by-sex interaction (p=0.531).
Groups differed in IQ whereby dependent and non-dependent cannabis users each had significantly lower IQ than controls, but there was no effect of sex on IQ, and there were no group-by-sex interactions. There were group effects on tobacco use and cannabis use: in both cases, cannabis user groups consumed significantly more tobacco and cannabis than the control group, there was no effect of sex on tobacco or cannabis use, and there was no group-by-sex interaction.
No control participants used cannabis on a regular basis (Supplementary Data). There is a group effect on ICV (p=0.046), with non-dependent users showing smaller volumes than controls, and there was a main effect of sex on ICV (p<0.001) whereby females have smaller ICV than males. However, there was no group-by-sex interaction effect on ICV.
Interactive and independent effects of cannabis dependence and sex on amygdala volumetry
Table 2 shows group, sex, and group-by-sex effects on amygdala volumetry, accounting for age, IQ, alcohol, tobacco, ICV, and site. There were no main effects of group on amygdala volumes, and there were no group-by-sex interactions in each case.
Overview of Volumetry for the Amygdala and Amygdala Nuclei, in Males and Females Across Dependent Cannabis Users, Non-Dependent Cannabis Users, and Controls
Post hoc column presents results from post hoc analyses where omnibus test parameters survived FDR correction. Values for amygdala and amygdala nuclei volumes are mean (SD). Bolded p-values remained significant following Benjamini-Hochberg FDR correction. Var, site-level variation estimated as an ICC.
FDR, false discovery rate.
However, there was a main effect of sex on total amygdala (both left and right), basolateral nucleus (left and right), and central nucleus (left and right) volumes (p's=<0.001–0.009). The main effects of sex on all amygdala volumes except for the right central nucleus remained significant following Benjamini-Hochberg FDR correction. In each case, females had smaller volumes than males.
Interactive and independent effects of cannabis dependence and sex on hippocampus volumetry
Table 3 shows group, sex, and group-by-sex effects on hippocampal volumes, accounting for age, IQ, alcohol, tobacco, ICV, and site. There was a main effect of group on left total hippocampus volume (F=3.41, p=0.035), whereby dependent cannabis users had lower volumes than non-dependent cannabis users (mean difference=157 mm3, p=0.01, d=0.22) but neither group differed from controls (p's=0.171 and 0.178).
Hippocampal Total and Subregional Volumes, in Males and Females Across Dependent Cannabis Users, Non-Dependent Cannabis Users, and Controls
Bolded p-values remained significant following Benjamini-Hochberg FDR correction. Values for hippocampal total and subregional volumes are mean (SD).
Post hoc column presents results from post hoc analyses where omnibus test parameters survived FDR correction.
CA1, cornu ammonis subfield 1; CA3, cornu ammonis subfield 3; Var, variation.
There were main effects of group on left and right CA1 (F=5.95, p=0.003 and F=4.52, p=0.01, respectively), whereby dependent cannabis users had lower volumes than non-dependent cannabis users (mean difference=46 mm3, p<0.001, d=0.32) and controls (mean difference=30 mm3, p=0.022, d=0.18). However, there were no group-by-sex interactions in CA1 volumes.
There was a main effect of sex on CA1 volumes (left: F=10.80, p=0.001; right F=7.66, p=0.006), where volumes were significantly smaller in females than males (mean difference=40 mm3, p=0.001 and 34 mm3, p=0.006, respectively).
There were main effects of group on left CA3 volume (F=5.00, p=0.008) and left dentate gyrus volume (F=5.09, p=0.007), with dependent users having smaller volumes than non-dependent users (left CA3: mean difference=19 mm3, p=0.002, d=0.37; left dentate gyrus (DG): mean difference=51 mm3, p=0.002, d=0.31) but neither group differed from controls (left CA3: p's=0.076 and 0.133; left DG: p's=0.077 and 0.124).
The main effects of group on left CA1, left CA3, and left DG and the main effects of sex on left and right CA1 remained significant following Benjamini-Hochberg FDR correction. However, the main effect of group on left total hippocampus and right CA1 volume did not survive adjustment. See Figure 1 for post-hoc tests. Sensitivity analysis were run for left CA1, left CA3, and left dentate gyrus, including cannabis cones past month as an additional covariate, and excluding controls. All main effects of group remained significant, in the same direction. Results are reported in Supplementary Data.
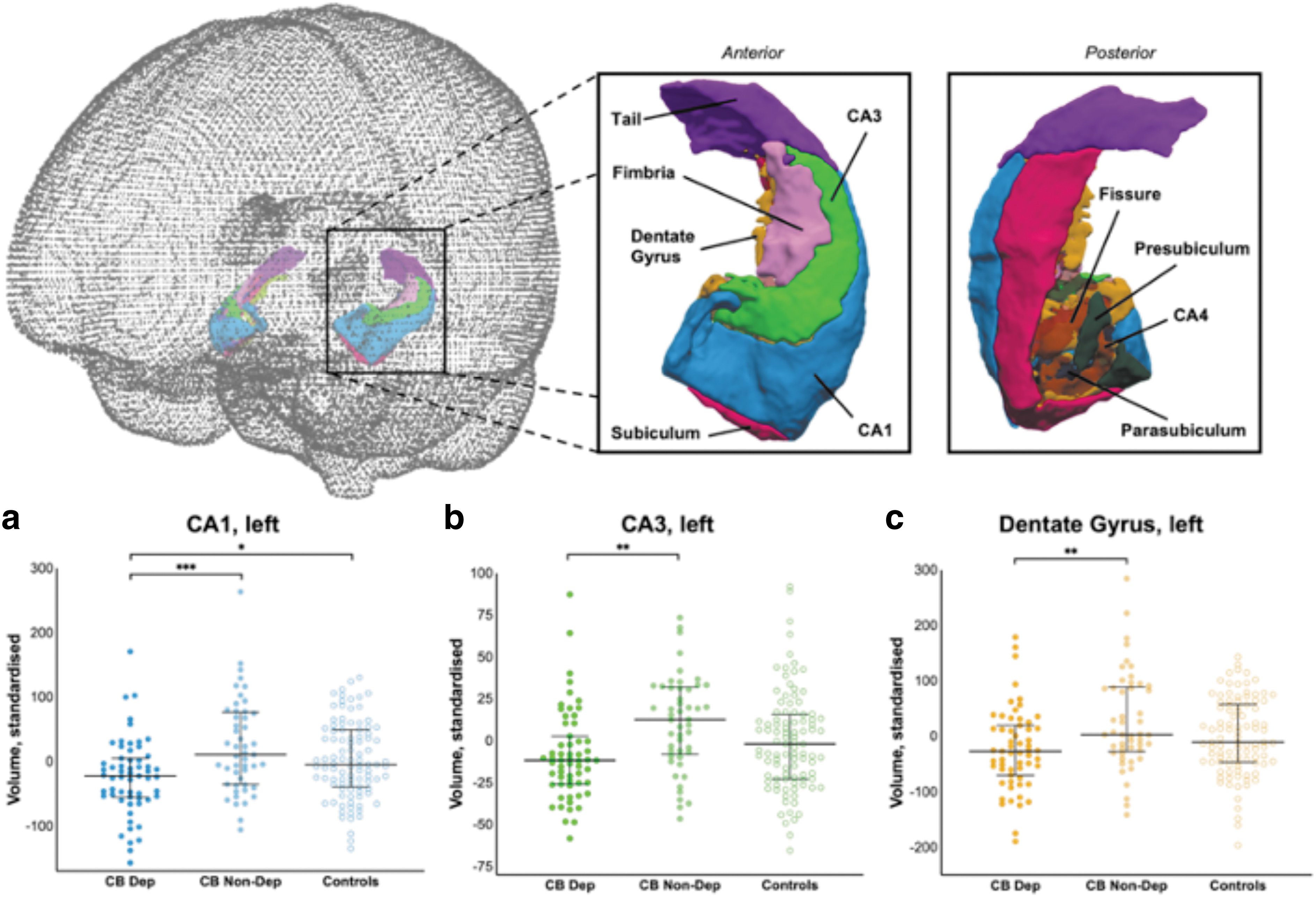
Overview of significant group differences for the volumes of the
There were no group-by-sex interactions on any measure of hippocampal volume.
Results from post hoc correlational analyses are reported in Supplementary Data.
Discussion
In line with our hypotheses, we observed group differences in the volumetry of hippocampus subfields whereby dependent cannabis users showed smaller CA1 volumes than non-dependent cannabis users and controls; and smaller CA3 and dentate gyrus volumes than non-dependent users, accounting for age, ICV, past month standard drinks, cigarettes used in the past month, ICV, and site. These differences had small effect sizes.
However, in opposition to our hypotheses, we found no group effects on amygdala volumetry, and importantly no group-by-sex effects on any hippocampus or amygdala subregion volumes, suggesting that observed group effects on hippocampus subfields are not moderated by sex. Additional exploratory analyses suggest that hippocampal and amygdala volumes were not predicted by age of cannabis onset, or current cannabis use dose after controlling for sex, age, ICV, IQ, alcohol, and tobacco exposure.
Individuals with cannabis dependence had significantly lower CA1 volumes than non-dependent cannabis users and controls, consistent with the hypothesis that cannabis dependence affects hippocampal subregional neuroanatomy. The finding replicates those showing smaller CA1 volumes in dependent cannabis users.42,43,51 Also, the CA1 changes over time in cannabis users have been reported to correlate with cannabis use related problems, that is, scores on the Cannabis Use Disorder Identification Test (CUDIT). 51
The findings also partially align with prominent neuroscientific theories of addiction whereby dependent use is postulated to be associated with neuroadaptations in learning and memory pathways, 67 and potentially extends such theories by confirming that the alterations are observed at a subregional level and in cannabis dependence.
The mechanisms driving such differences remain unclear. The CA1 might be vulnerable to THC exposure. Indeed, CB1 receptors are abundant in GABAergic terminals within the CA1, and THC can change glutamatergic transmission within hippocampal slices. 48 Meanwhile, chronic THC exposure can lead to reduced CA1’ neuronal dendritic spine density, 68 CB1 receptor sensitivity,69,70 synaptic plasticity 71 and downregulation of CB1 receptors.52,72
Therefore, THC exposure might have led to volumetric reduction within the CA1 in dependent cannabis users. Future studies should examine how hippocampal subfields in cannabis dependence relate to: objective metrics of cannabinoid exposure (e.g., THC amount in cannabis products consumed, THC metabolites in urine); and of cannabis use related problems with diagnostic cut-offs (e.g., CUDIT).
People with cannabis dependence showed lower CA3 and dentate gyrus volumes compared with those without dependence but not controls, even though those with cannabis dependence had the lowest mean values for volumetry among the groups. Similarly, prior work reports no detectable difference in CA3 volumes between dependent and non-dependent cannabis users versus controls. 42
The CA3 and the dentate gyrus may be less vulnerable to the impact of cannabis dependence, perhaps due to neurogenetic processes that take place within the dentate gyrus 64 that sends dense projections to the CA3. 73 Also of relevance, measures of anxiety and depression were not available for the present cohort. As both have been previously shown to be comorbid with dependence, 74 we cannot discount the possibility that unobserved mental health factors may be confounding effects.
For differences between dependent cannabis users and controls to be detectable, a more severe level of dependence or endorsement of cannabis dependence for a longer period may be required.
We found no significant differences between non-dependent users and controls in any of the regional volumes measured. This is in line with our previous work43,58 and suggests that exposure alone might not be sufficient to change hippocampal/amygdalar neuroanatomy, and that dependence might be required for observable changes to occur. 7
Inferring implications of these small effects through consulting the extant literature remains challenging due to heterogeneous assessment tools and study designs, and the resultant inconsistentcy in hypothesized brain-behavior correlates. With the eventual goal of discovering mental health treatment targets, future research should prioritise field-standard assessment tools, 75 intervention studies, 76 and longitudinal follow ups 77 to observe how changes in dependence symptoms may be accompanied by neuroanatomical alterations within hippocampal subregions.
The effect of group on the total hippocampal volume did not survive correction for multiple comparisons, consistent with prior work 23 in young adults aged 22–24 such as those examined herein. As major neurodevelopmental processes take place until the age of 26, 78 youth age might protect from hippocampal volume reduction in cannabis users. 39
Perhaps, at younger ages, hippocampal volume reduction initially appears at a subregional level (e.g., within the CA1) and with continued cannabis exposure through later in adulthood, it progressively expands to other subregions to affect total hippocampal volumes.
We also raise the possibility that the observed structural differences predated onset of cannabis use, and may have [directly or indirectly] influenced progression from recreational use to dependence. While the present study's cross-sectional design precludes exploring this possibility, previous work has observed that early environment (socioeconomic status) may influence the trajectory of hippocampal development, and therefore life outcomes. 79
Future longitudinal work should more finely explore the relationship of prenatal and early-childhood factors with both hippocampal developmental trajectories and substance use outcomes.
Interestingly, while it was not a primary focus of the current paper, we observed main effects of sex on total amygdala volume and its bilateral basolateral and left central nuclei, and volume of the left hippocampal CA1. Without a group-by-sex interaction in the current analyses, we can make no assertions about how sex impacts cannabis-related neuroanatomical alterations; however, findings are consistent with neuroimaging studies of normative samples80–82 and highlight the need to account for the effect of sex when measuring subregional neuroanatomy in humans.
Limitations
Several methodological limitations need to be considered when interpreting these results. First, as the consortium relies on already collected data, sites used inconsistent diagnostic assessment of cannabis dependence (Supplementary Table S1 in the Supplementary Data). Future work should replicate the findings using updated diagnostic methods (e.g., DSM-571). Second, volumetry might have been systematically affected by site-specific MRI data acquisition parameters. To mitigate this issue, we implemented standardized MRI quality check protocols (http://enigma.ini.usc.edu/protocols/imaging-protocols), and accounted for systematic site-related variation by modeling it as a random effect in linear mixed models.
Third, sites had inconsistent metrics of variables (e.g., depression, anxiety, psychosis29,32,34) known to affect variation in volumetry.83,84 We (partially) mitigated this issue via screening for diagnosis of major mental health disorders. Future work should measure mental health symptoms using tools with diagnostic cut-offs and in relation to volumetry, to elucidate the clinical significance of the volumetric findings.
Also, we lacked consistent data on cannabis exposure across sites (e.g., duration, cannabinoid compounds such as THC and cannabidiol) and lacked measures of sex hormones. Given the increased availability, potency, and diversification of cannabis products, 85 and the influence of sex hormones on volumetry, 86 future work should integrate metrics of cannabinoids and sex hormones to unpack dosage versus dependence effects in both sexes. Thus, future studies should report cannabis use systematically, including the use of standard THC units that have received global support from researchers and health organizations.44,87–93
Further, unexamined exogenous variables related to biological sex (e.g., contraceptive pills/devices 94 ), and gender roles (e.g., differential caregiving responsibilities95,96) may affect levels of perceived stress and cortisol. This could be of relevance in the present study, as previous research shows that the amygdala and hippocampus function in relationship with the HPA (hypothalamic-pituitary axis); mediating its activity (through facilitation and inhibition, respectively), and receiving input via glucocorticoid receptors.97,98
Finally, our study was cross-sectional: volume differences may predate cannabis dependence onset and contribute to the initiation of cannabis dependence. Longitudinal studies should examine whether the differences in subregional neuroanatomy predate, predict, or follow the onset of cannabis dependence and the time course of the volumetric differences observed herein. Finally, although we found no group-by-sex effect on subregional volumetry, concluding this effect is not present is limited due to the relatively small sample size.
Conclusion
Cannabis dependence was associated with lower volumes within select hippocampus subfields (CA1) in both males and females, after adjusting for age, ICV, past month standard drinks, past month cigarettes used, and site. The cross-sectional design precluded understanding whether these effects predated or followed the onset of cannabis dependence.
We warrant the systematic and concurrent measurement of cannabis dependence and cannabinoid exposure metrics, a subregion-specific approach, and longitudinal design. Such new studies will be critical to gain insight into the neurobiological mechanisms underlying the onset and continuation of cannabis dependence in vulnerable people through youth and adulthood.
Given the increased potency and availability of cannabis products, such new knowledge will be relevant to identify the pathophysiology of increasingly prevalent CUDs, with the goal of informing neurobiological targets for treatment.
Footnotes
Acknowledgments
The authors thank all participants for the time and effort they took to partake in the studies. They also thank D. Thomson, D. Tichelaar, E. Robinson, E. Watt, I. Starcheus, J. Gladwell. J. Tzaferis, K. Kearney, and S. Mizzi, for their significant contributions to the image quality checks. They thank P. Saemann for providing comments on code to generate the images for visual inspection. They thank J. Dominguez for his helpful comments on the code to perform the FreeSurfer segmentation.
Authors' Contributions
V.L. and A.G. co-wrote the first draft (methods/results; and introduction/discussion respectively). A.G. revised the full draft. V.L. further revised the full draft. C.A.R. provided revisions of the full draft and following drafts for co-author feedback. V.L. and C.A.R. integrate co-authors feedback on the draft for submission. M.G.R., Y.C., P.R., S.G., and V.L. conceived and supervised all the statistical and neuroimaging analyses, tables, and figure. E.M. conducted all statistical analyses and produced the three tables of results and figure. A.C. provided input on the manuscript writing. S.G. conducted all neuroimaging data analyses. C.S. provided technical support for neuroimaging data analyses. A.B., M.B., P.B., Y.C., P.C., J.C., H.G., N.S., P.M.T., M.Y., and the ENIGMA Addiction Working Group facilitated data collection, prepared individual datasets, and conducted primary analyses; I.L., P.R., and G.T. provided resource support. All authors reviewed the manuscript and approved it for submission.
Author Disclosure Statement
C.A.R. discloses that he has received grant funding from Unilever, and has acted as consultant for pharmaceutical company Boehringer Ingelheim. Neither of these associations are related to the current work. P.M.T. received partial grant support from Biogen, Inc., for research unrelated to this manuscript. M.Y. has received funding to conduct sponsored Investigator-Initiated trials (including Incannex Healthcare Ltd.). These funding sources had no role in the design, management, data analysis, presentation, or interpretation and write-up of the data. M.Y. also sits on the Advisory Boards of Centre of The Urban Mental Health, University of Amsterdam; Enosis Therapeutics; and Monash Biomedical Imaging Centre. This funding is unrelated to the research reported in this manuscript. All other authors has no conflict of interest to declare.
Funding Information
This work was supported by the MASSIVE HPC facility (![]() ). V.L. was supported by the Investigator Grant from the National Medical and Health Research Council (NHMRC ID: 2016833), the Al & Val Rosenstrauss Fellowship, and Australian Catholic University through a competitive scheme.
). V.L. was supported by the Investigator Grant from the National Medical and Health Research Council (NHMRC ID: 2016833), the Al & Val Rosenstrauss Fellowship, and Australian Catholic University through a competitive scheme.
N.S. received funding from the Clive and Vera Ramaciotti Foundation for Biomedical Research, and the National and Health and Medical Research Council Project grant 459111 and was supported by the Australian Research Council (ARC) Future Fellowship FT110100752. J.C. received funding for the Cannabis Prospective study from ZonMW grant 31180002 from NWO. M.Y. receives funding from other NHMRC schemes, Monash University, and Australian Government funding bodies such as the ARC, Australian Defence Science and Technology (DST), and the Department of Industry, Innovation and Science (DIIS). He has also received philanthropic donations from the David Winston Turner Endowment Fund, Wilson Foundation, as well as payments in relation to couaurrt-, expert witness-, and/or expert review-reports.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.