
Abstract
Introduction:
Cannabis sativa extract has been used as an herbal medicine since ancient times. It is one of the most researched extracts, especially among supportive treatments against cancer. Prostate cancer is one of the most frequently diagnosed cancer types in men worldwide and an estimated 288,300 new cases were diagnosed in 2023. Today, many advanced therapeutic approaches are used for prostate cancer, such as immunotherapy and chemotherapy, but acquired drug resistance, long-term drug usage and differentiation of cancer cells mostly restricted the efficiency of therapies. Therefore, it is thought that the use of natural products to overcome these limitations and improve the effectiveness of existing therapies may offer promising approaches. The present study focused on the investigation of the possible enhancer role of cannabidiol (CBD), which is a potent ingredient compound of Cannabis, on the chemotherapeutic agent etoposide in prostate cancer cells.
Methods:
Herein, we tested the potentiator role of CBD on etoposide in prostate cancer cells by testing the cytotoxic effect, morphological alterations, apoptotic effects, autophagy, unfolded protein response (UPR) signaling, endoplasmic reticulum-associated degradation mechanism (ERAD), angiogenic and androgenic factors, and epithelial–mesenchymal transition (EMT). In addition, we examined the combined treatment of CBD and etoposide on colonial growth, migrative, invasive capability, 3D tumor formation, and cellular senescence.
Results:
Our findings demonstrated that cotreatment of etoposide with CBD importantly suppressed autophagic flux and induced ERAD and UPR signaling in LNCaP cells. Also, CBD strongly enhanced the etoposide-mediated suppression of androgenic signaling, angiogenic factor VEGF-A, protooncogene c-Myc, EMT, and also induced apoptosis through activation caspase-3 and PARP-1. Moreover, coadministration markedly decreased tumorigenic properties, such as proliferative capacity, colonial growth, migration, and 3D tumor formation and also induced senescence. Altogether, our data revealed that CBD has a potent enhancer effect on etoposide-associated anticancer activities.
Conclusion:
The present study suggests that the use of CBD as a supportive therapy in existing chemotherapeutic approaches may be a promising option, but this effectiveness needs to be investigated on a large scale.

Introduction
Prostate cancer is one of the most frequently diagnosed cancer types in men and an estimated 268,490 new cases were diagnosed in 2022. 1 In 2023, it is estimated that 288,300 patients will be diagnosed with prostate cancer and 34,700 deaths will occur from prostate cancer in the United States. 2 Considering the increase in prostate cancer cases diagnosed over the years, it is predicted that this number will increase day by day. Surgery, radiotherapy, hormonal therapy, and chemotherapy are the main therapeutic options for the treatment of prostate cancer.3–5 Furthermore, various chemotherapeutics, including paclitaxel and etoposide are preferred in the treatment protocols. However, due to their potential to develop resistance and cause serious systemic side effects with long-term treatment, there remains a need to investigate new treatment options or adjuvant therapies. 6 Also, newly developed treatment modalities selectively targeting the molecular signaling pathways are expected to be more efficient against prostate cancer. From this perspective, it is thought that research on natural products will contribute to developing more effective treatment options in the future.
Etoposide (VP-16, epipodophyllotoxin) is the first recognized topoisomerase-II inhibitor and one of the anticancer drugs that are derived from natural compounds. It is a semisynthetic derivative of podophyllotoxin, which is a nonalkaloid lignan isolated from the dried roots and rhizomes of Podophyllum peltatum or Podophyllum emodi species. The clinical use of etoposide has been approved since 1983 in the United States and is still extensively used in the treatment of many cancer types, including lung and breast cancer, brain tumors, leukemia, and testicular carcinoma.7–11 It has also efficient anticancer effects in patients with metastatic castration-resistant prostate cancer. 6 However, etoposide has disadvantages that limit its therapeutic efficacy, such as low water solubility, metabolic inactivation, poor bioavailability, and development of drug resistance. 12 Furthermore, it can cause severe side effects, including immunosuppression, nephrotoxicity, cardiotoxicity, and also acute myelocytic leukemia.12,13 Therefore, ongoing efforts are to reduce drug-related side effects and increase therapeutic efficacy by combining existing anticancer drugs with other drugs or natural compounds. In the present study, we aimed to investigate the results of the combination of etoposide and cannabidiol (CBD) on prostate cancer cells.
Cannabis sativa (hemp or marijuana) plant extract has been used as an herbal medicine since ancient times. 14 There are over 100 different cannabinoids that were isolated from C. sativa; Δ9–tetrahydrocannabinol (Δ9-THC) and CBD are the most widely known and studied derivatives.15,16 The therapeutic activities of cannabinoids in cancer treatment have long been of great interest. 17 The derivatives of cannabinoids, including Δ8-THC, Δ9-THC, and CBD have been shown to inhibit DNA synthesis and growth of lung adenocarcinoma in cultured cells and mouse tumor models. 18 Furthermore, their anticancer effects have been demonstrated by in vitro and in vivo models of other cancer types, such as breast, pancreas, prostate cancers, lymphoma, and glioma.19–21 Also, the combinatory effect of CBD with several anticancer agents, including doxorubicin, oxaliplatin, temozolomide, and methotrexate, are extensively investigated in various cancer models such as osteosarcoma.22–24 The predicted anticancer mechanisms of CBD are cell cycle arrest and induction of apoptosis, inhibition of migration, adhesion, and invasion. 25 Moreover, cell cycle, angiogenic mechanisms, and also protein quality control mechanisms are intensively investigated in terms of the biochemical activity of anticancer drugs and natural compounds.21,26 Recent studies have drawn attention to the role of protein control mechanisms in tumor progression for various cancer types, including prostate cancer. A recent study noted that CBD suppresses autophagy and modulates the endoplasmic reticulum (ER) stress signaling and ER-associated protein degradation (ERAD) mechanism in prostate cancer cells, which are the main components of the protein quality control process of eukaryotic cells. 21
Recent studies have focused on ER protein quality control mechanisms such as ERAD and ER stress-related signaling mechanisms, such as unfolded protein response (UPR) signaling, which may be a potential target for developing drugs that reduce hypoxia, angiogenesis, inflammation, and drug resistance. Moreover, the effect of various anticancer agents and their combined administration with natural compounds on tumorigenesis via ER stress has been investigated.26–31
Autophagy is another physiologically important mechanism in eukaryotic cells that plays a critical role in the elimination of macromolecules, such as misfolded or unfolded proteins and also damaged organelles and intracellular pathogens.32–33 It has also been involved with pathophysiological conditions in humans, including cancer.34,35 Besides, it has a dual role in tumorigenesis, tumor progression, and chemotherapy resistance of cancer cells. 36 In addition, autophagy is recognized as a type II programmed cell death mechanism. It may trigger cell death in nutrient deprivation or under stressful cellular conditions. 37 Although autophagy-targeted anticancer therapies have been intensively investigated for a long time, no conclusive data on their advanced role associated with tumor suppressive or oncogenic function have been provided.
Recent studies have shown that natural products as a potential source of biologically active metabolites have a critical role in drug development for cancer therapy. 38 Moreover, evidence suggests that natural products have been shown to exert anticancer effects by modulating autophagy in various cancer types, including breast, liver, ovarian, pancreatic, and prostate cancers.39,40
In the present study, we aimed to investigate the possible potentiator roles of CBD on anticancer effects of etoposide in prostate cancer cells. Although etoposide is an effective anticancer drug, long-term use of etoposide can cause serious health problems due to its severe systemic side effects. For this aim, we evaluated the booster role of CBD on etoposide in prostate cancer cells by testing their roles on androgen signaling, autophagy, ERAD, and apoptotic cell-death-related critical executor proteins. We also tested the effect of combinatory treatment on invasion, migration, and colonial and tumor growth of prostate cancer cells. Our findings suggest that CBD has significant effects on the enhancement of the anticancer effects of etoposide.
Material and Methods
Materials
Cell culture ingredient materials, including growth media, fetal bovine serum (FBS), L-glutamine, 0.25% and 0.05% Trypsin, and other requirements were provided from Biological Industries. Etoposide (#341205) was purchased from Sigma-Aldrich. CBD was obtained from Suleyman Demirel University, Natural Products Application and Research Center (SUDUM). The source of the CBD was the extract of Cannabis sativa L., Cannabaceae. CBD content is >99.9 and the THC content is <0.01. limits of residual alcohol and heavy metals comply with United States Pharmacopeia (USP) and European (EU) pharmacopeia. CBD was solubilized in ethanol (EtOH) as 1000× concentrated. So that applied CBD to cells, the ethanol concentration did not exceed 0.1%.
Staurosporine (#9953) and bafilomycin A1 (#54645) were purchased from Cell Signaling Technologies. X-Gal (5-Bromo-4-chloro-3-indolyl α-D-galactopyranoside) (#3117073001) was provided from Roche.
Rabbit polyclonal antibodies anti-glycoprotein 78 (gp78; #16675-1-AP; 1:3000), anti-c-Myc (#10828-1-AP; 1:2500), anti-protein kinase R (PKR)-like endoplasmic reticulum kinase (PERK; #24390-1-AP; 1:2000), anti-IRE1α (#27528-1-AP; 1:2000), anti-XBP-1s (#24868-1-AP; 1:1000), anti-caspase-3 (#19677-1-AP; 1:1500), anti-ubiquitin (#10201-2-AP; 1:1000), anti-PSA (#10679-1-AP; 1:2500), anti-AR (#22089-1-AP; 1:2000), anti-E-cadherin (#20874-1-AP; 1:2500), anti-N-cadherin (#22018-1-AP; 1:2500), and anti-PARP-1 (#13371-1-AP; 1:3500) were obtained from Proteintech. Rabbit polyclonal anti-eIF2α (#9722; 1:2000), anti-phospho-eIF2α (Ser51; #9721; 1:1000), anti-LC3A/B (#12741; 1:2500), anti-p62/SQSTM1 (#5114; 1:2000), and rabbit monoclonal antibody against HMG-CoA reductase degradation protein 1 (Hrd1; #14773; 1:3000) were obtained from Cell Signaling Technology and polyclonal rabbit anti-VEGF-A (#E-AB-53277; 1:2000) was purchased from Elabscience. Mouse monoclonal anti-beta-actin antibody (#A5316; 1:10,000) was purchased from Sigma-Aldrich. HRP-conjugated goat anti-mouse (#31430; 1:5000) or goat anti-rabbit (#31460; 1:5000) IgG (H + L) was purchased from Thermo Scientific.
Cell culture
Human androgen-sensitive prostate adenocarcinoma cell line, LNCaP (CRL-1740TM), was obtained from the American Type Culture Collection (ATCC). LNCaP cells were cultured in RPMI-1640 media enriched with 10% FBS and 2 mM L-glutamine (Capricorn Scientific). Cells were propagated in conventional cell culture conditions (humidified atmosphere of 5% CO2 and 95% air at a constant temperature of 37°C).
WST-1 assay
Cell proliferation was analyzed by WST-1 cell proliferation premix (#MK400) according to the manufacturer’s instruction (TaKaRa). A total of 10.000 cells were seeded per well on a 96-well plate and 24 h later, compounds were applied to the cells at indicated doses for 48 h. After that, WST-1 cell proliferation premix was applied to the cells according to the manufacturer’s protocol. The absorbance was measured at 450 nm, with 600 nm set as the reference wavelength by microplate spectrophotometer (BioTek, Epoch 2). Averages of the absorbance values were calculated. Data were presented as a % cell proliferation in a graph. The proliferation analysis was performed in 3 biological and 3 technical replicates.
Microscopic examination
Morphological alterations were examined by using a phase-contrast inverted microscope (Sunny SopTop ICX41) and a digital camera system (OD400UHW). Cells were seeded in a 12-well plate at 1.5 × 105 cells/well, and 24 h later, compounds were administrated for 24 h. After that, cells were photographed at a magnification of 20x lens using brightfield microscopy.
Protein isolation and immunoblotting
Proteins were isolated from cells as described before. 26 Cell pellets were lysed with radioimmunoprecipitation assay (RIPA) buffer containing a mammalian protease inhibitor cocktail and then centrifugated at 14.000 rpm for 20 min at 4°C. The supernatant was collected and total protein concentration was determined by bicinchoninic acid (BCA) protein assay (TaKaRa #T9300A). Typically, 30 μg of soluble protein was used in immunoblotting studies. Protein samples were mixed with 4× Laemmli buffer and denatured at 70°C for 15 min. Samples were separated on hand-cast 10–15% sodium dodecyl sulfate–polyacrylamide gels and transferred to an Immun-Blot® polyvinylidene difluoride membrane (Bio-Rad). The membrane was blocked in 5% nonfat dry milk in phosphate-buffered saline (PBS) containing 0.1% Tween (PBS–Tween) for 1 h at room temperature. The primary and origin-specific secondary antibodies were applied for 2 h at room temperature, respectively. Interested proteins were monitored by using an enhanced chemiluminescence (ECL) solution (Thermo Scientific) in ChemiDoc XRS+ system (Bio-Rad). The protein bands were quantitated by Image StudioTM Lite (LI-COR®) software and indicated on the protein blots. Also, the quantification of independent experiments was presented as a bar graph with ±SD in the Supplementary Data S1. All analyses were made in triplicate.
Quantitative real-time PCR
Total RNA was isolated from cell pellets by using the Monarch® Total RNA Miniprep Kit (NEB #T2010) (New England Biolabs) according to the manufacturer’s protocol. The quality of RNA was determined by a micro-spectrophotometer (Allsheng). Around 1 μg RNA was reverse transcribed using the iScript™ cDNA Synthesis Kit with oligo dT and random primers (Bio-Rad). The 5 min at 25 ̊C, 20 min at 46 ̊C and for 1 min at 95 ̊C steps were followed in cDNA synthesis. To amplify the targets, primers were specifically designed for the gene of interest, including BAX (ID:581) and BCL-2 (ID:596). The information on primer sequences is available upon request. Real-time PCR amplification was performed using SsoAdvancedTM Universal SYBR Green Supermix (Bio-Rad) according to the manufacturer’s protocol and the fluorescence signal was detected on a CFX connect instrument (Bio-Rad). Each sample was examined in triplicate for each quantitative real-time PCR (qRT-PCR). Ribosomal Protein Lateral Stalk Subunit P0 (RPLP0) was used as a housekeeping gene and RPLP0 mRNA levels were used as an endogenous control for normalization. The mRNA expressions were expressed as normalized ratios in relative arbitrary units. Averaged triplicate measurements were scaled by standard deviation. Relative quantification of gene expression was calculated with the comparative 2−ΔΔCt method. 41 MIQE guidelines were followed during the qRT-PCR. 42
Live and dead cell staining
Cells were seeded in a 12-well plate (1.5 × 105 cells/mL). After that, cells were treated with compounds for 24 h and full-media was aspirated and cells were washed with 1×PBS twice. Cells were stained using the LIVE/DEADTM Cell Imaging Kit (InvitrogenTM) according to the manufacturer’s protocol. Stained cells were visualized using a fluorescence microscope (Olympus EX71 microscope, DP74 camera system, Japan) at fluorescein isothiocyanate (FITC) and Texas Red™ filters.
DAPI staining
Cells were seeded in a 12-well plate (1.5 × 105 cells/mL) included in the cover slide. Thereafter, cells were treated with compounds for 24 h and then cells were washed with ice-cold 1×PBS twice and fixed with 4% formaldehyde solution. Then, cells were blocked with bovine serum albumin (BSA) and stained with DAPI (4′,6-diamidino-2-phenylindole, dihydrochloride) (InvitrogenTM). Nuclear staining was visualized with a fluorescence microscope (Olympus EX71 microscope, DP74 camera system, Japan) at a DAPI filter. Quantification was carried out by counting the condensations in five independent fields. The experiment was performed as three biological replicates. Data were presented in the graph as the fold change compared with the control group.
Propidium iodide staining
Cells were seeded in a 12-well plate (1.5 × 105 cells/mL) and when cells were attached to the cell culture plate, compounds were administered for 24 h. After that, cells were harvested and washed with ice-cold 1×PBS twice and centrifugated at 300 g for 5 min. Cells were gently resuspended with a staining buffer containing propidium iodide (PI) and incubated at room temperature for 10 min. After 30 μL cell suspension spreading the slide and investigating in a fluorescence microscope (Olympus EX71 microscope, DP74 camera system, Japan) at a PI fluorescence channel. The experiment was performed as three biological replicates.
Senescence-associated β-galactosidase assay
Cells were seeded in 12-well cell culture plates. After cells were treated with compounds for 24 h and then cells were washed twice with 1×PBS and fixed with 4% formaldehyde solution. Cells were then incubated at 37°C for 2 days in X-gal staining solution (1 mg/mL X-gal, 40 mM citric acid/sodium phosphate (pH 6.0), 5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, 150 mM NaCl, 2 mM MgCl2) and then washed twice with 1×PBS and fixed with methanol and then monitored at a magnification of 20× objective using brightfield microscopy. To quantify senescent cells, images were taken from five independent fields and stained cells were counted. The experiment was performed as three biological replicates.
2D colony formation assay
Cells were seeded in six-well plates with 1000 cells/mL grown under conventional culture conditions for 24 h and then compounds were applied to the cells for 72 h. At the end of the experiment, cells were washed twice with ice-cold 1×PBS. Afterward, grown colonies were fixed with methanol and stained with 0.05% Crystal Violet in 1×PBS. % colonial growth was quantified with ImageJ software. The experiment was performed as three biological and three technical replicates.
Wound-healing assay
Cells were seeded in 12-well cell culture plates with 2 × 105 cells/well and propagated under conventional cell culture conditions for 24 h. After that, the wound area was created using a yellow 200 μL tip, and cells were washed twice with cell culture grade preheated 1×PBS to remove the cell residues. After this, compounds were applied and cells were grown in a culture medium for 72 h. The percentage of wound closure rates was analyzed with ImageJ software (http://imagej.nih.gov/ij/). The experiment was performed as three biological and three technical replicates.
3D cell culture assay
Soft agar assay was modified according to Borowicz et al. 43 Equal volumes of 2× RPMI-1640 media enriched with 20% FBS and sterile 1.2% low-melting agar were gently mixed and added to 12-well plates. Cell suspensions prepared in 150 μL were mixed with 250 μL of 2× RPMI-1640 media enriched with 20% FBS and 250 μL of 0.6% agar and then placed on the solidified bottom agar layer. After the addition of 100 μL of 2× media containing 10% FBS to each well, the plate was incubated for 1 day and then Thr was administrated to the cells. The images of cells were taken using Sunny SopTop invert microscope and OD400UHW camera system. To quantify the growth of 3D formations, five images were taken from independent areas, and the developing colonies were counted.
Statistical analysis
Data were presented as means ± standard deviation. The statistical significance of differences between groups was determined by a two-tailed equal variance Student’s t-test with a minimum of 95% confidence interval or one-way ANOVA followed by post-hoc Tukey’s test was used for multiple pairwise comparisons by GraphPad Prism 7. The significant level was set at 5% and probability values of p < 0.05 were considered statistically significant for all tests.
Results
Cotreatment of etoposide with cannabidiol more efficiently limits the proliferative capacity of prostate cancer cells
First, we investigated the possible antiproliferative booster effect of CBD on etoposide in LNCaP cells. With this aim, we treated the cells with 10 μM etoposide; 5, 10, and 20 nM CBD or combined with CBD or etoposide for 48 h. Nontoxic CBD concentrations were selected based on previous studies on prostate cancer cell lines. 21 Microscopic examinations revealed that CBD treatment only did not induce the cell death pattern in LNCaP cells, whereas etoposide administration markedly induced the shrinking and rounding cell formation (Fig. 1a). Combined treatment was also very similar to the etoposide-induced cell formation trend (Fig. 1a). Moreover, WST-1 assay revealed that CBD treatment slightly reduced the proliferation rate of LNCaP cells in a dose-dependent manner (Fig. 1b). As expected, etoposide administration significantly decreased the proliferation of LNCaP cells. Combined treatment of etoposide with CBD more strongly reduced the proliferation of LNCaP cells compared with indicated doses of CBD or etoposide treatment alone (Fig. 1b).

Testing the effect of etoposide, cannabidiol and their combination on cell viability of LNCaP cells. Cells were treated with 10 µM etoposide and 5, 10, and 20 nM CBD or with their combination for 48 h.
Combined treatment of cannabidiol and etoposide markedly suppressed the autophagic flux in LNCaP cells
To test the possible enhancer effect of CBD and etoposide on autophagy in LNCaP cells, the levels of autophagy-related proteins Sequestosome 1 (p62/SQSTM1) and conversion of microtubule-associated protein 1A/1B-light chain 3 (LC3)-I to LC3-II were evaluated by immunoblotting. Selective turnover of p62/SQSTM1 protein is often used to determine the autophagic flux in mammalian cells. The conversion of LC3-I to LC3-II through lipidation is utilized to analyze the autophagic status. 44 Herein, we also used the well-characterized autophagic inhibitor Bafilomycin A1 (BafA1) as a positive control of autophagy inhibition. 44 CBD treatment dose-dependently increased the p62/SQSTM1 levels compared with the control group. The conversion from LC3-I to LC3-II increased due to CBD administration in a dose-dependent manner (Fig. 2a). Etoposide treatment also increased the level of p62/SQSTM1, whereas conversion from LC3-I to LC3-II was not importantly affected by etoposide (Fig. 2a). More strikingly, cotreatment of etoposide with CBD strongly increased the protein level of p62/SQSTM1 and conversion from LC3-I to LC3-II in a dose-dependent manner (Fig. 2a). Similar results were obtained in the BafA1-treated group; the levels of p62/SQSTM1 increased and the conversion from LC3-I to LC3-II exhibited a similar pattern (Fig. 2a).
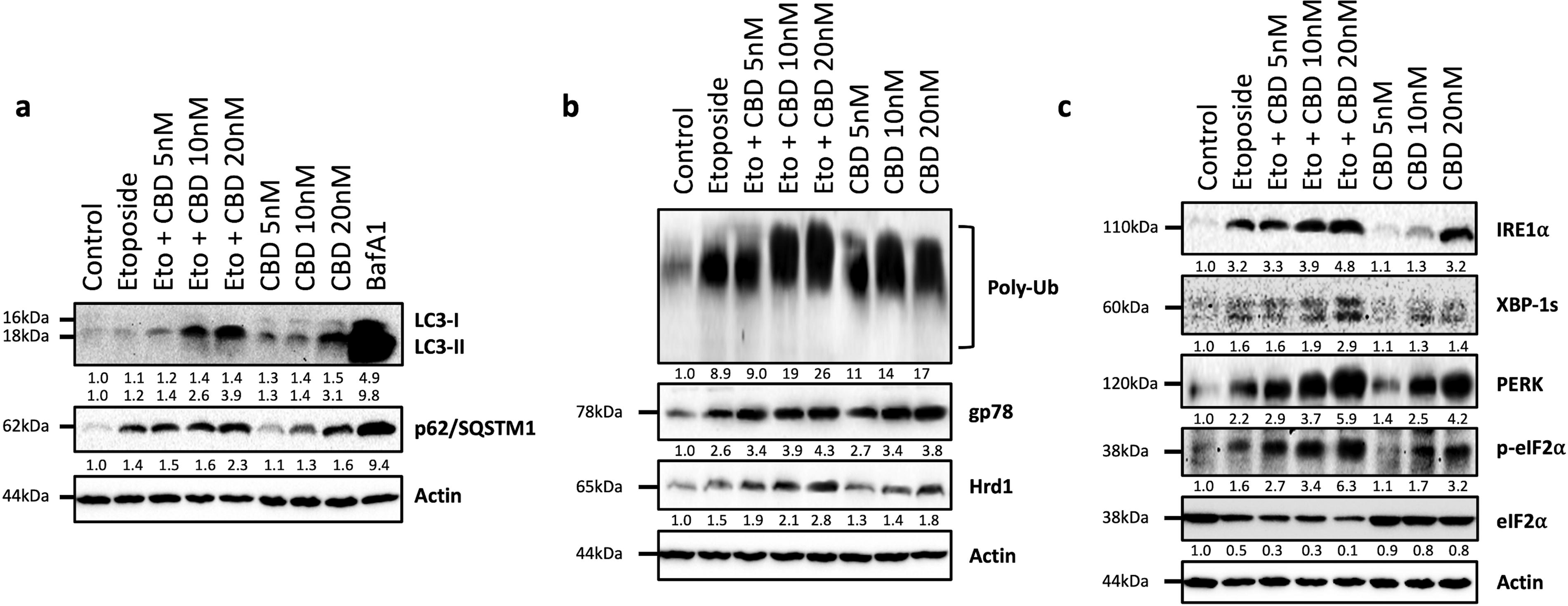
Evaluation of the effect of etoposide, cannabidiol, and their combination on autophagic flux and UPS and UPR signaling-related proteins in LNCaP cells.
Combined treatment significantly induces the protein levels of ERAD-related E3 ligase enzymes and also polyubiquitinated protein status in prostate cancer cells
We examined the effect of coadministration of CBD and etoposide on ERAD E3 ligase enzymes, including Hrd1 and gp78. Our results revealed that combined treatment dose-dependently increased the expression level of Hrd1 and gp78 proteins (Fig. 2b). Also, CBD or etoposide treatment alone increased the levels of Hrd1 and gp78 compared with the control group. Additionally, we investigated the steady-state levels of polyubiquitinated protein status in LNCaP cells. We also obtained similar results to ERAD proteins, combined treatment significantly increased the polyubiquitinated protein status in LNCaP cells compared with CBD or etoposide treatment alone (Fig. 2b).
Cannabidiol and etoposide combined treatment induces ER stress by activating IRE1α and PERK
Abnormal accumulation of unfolded or misfolded proteins at the ER induces cellular stress called ER stress. ER proteostasis surveillance is mediated by UPR, a signal transduction pathway. UPR sensitivelyadjusts the protein folding capacity of the eukaryotic cells by activating ER membrane-resident transmembrane sensor proteins, including PERK and inositol-requiring enzyme 1α (IRE1α) branches. 45 In addition, this mechanism can also work as a survival or cell death-inducing mechanism for prostate cancer cells. 46
In this study, we tested the effect of cotreatment of CBD and etoposide on PERK and IRE1α branches of UPR signaling. CBD or etoposide treatment alone increased the activated IRE1α-mediated produced spliced X-box-binding protein 1 (XBP-1s) and total IRE1α protein levels compared with the control group (Fig. 2c). Coadministration more strongly induced the XBP-1s and IRE1α protein levels (Fig. 2c).
We also found that combined treatment markedly induced the phosphorylation of eukaryotic translation initiation factor 2 subunit alpha (eIF2α) that is controlled by activated PERK signaling. 47 In addition, total PERK protein levels were increased in a dose-dependent manner, whereas total eIF2α was decreased in etoposide or CBD alone groups (Fig. 2c). Similar results were also obtained with CBD and etoposide cotreatment, but the combined treatment more strongly affected all tested IRE1α and PERK signaling proteins (Fig. 2c).
Coadministration of cannabidiol and etoposide causes apoptotic cell death in prostate cancer cells by inducing cleavage of caspase-3 and PARP-1 and also causing DNA fragmentation and loss of cellular integrity
To test the effects of CBD and its combination with etoposide on apoptotic cell death, the levels of total and cleavage forms of caspase-3 and PARP-1 proteins were evaluated by immunoblotting. The cleavage of caspase-3 is the main mediator for apoptosis and is responsible for the cleavage and inactivation of various proteins, including, nuclear enzyme poly (ADP-ribose) polymerase-1 (PARP-1). Also, it is one of the main cleavage targets of caspase-3 and the 89 kDa cleavage form is considered a marker of apoptosis. 48 CBD administration did not affect the levels of cleavage forms of caspase-3 and PARP-1 proteins compared with the control group. Etoposide treatment significantly induced cleavage forms of caspase-3 and PARP-1 proteins (Fig. 3a). Cotreatment of CBD and etoposide remarkably decreased the total caspase-3 and PARP-1 levels in a dose-dependent manner and also strongly increased cleavage forms of caspase-3 and PARP-1 compared with both CBD and etoposide-alone groups (Fig. 3a). Also, in these studies, a well-known apoptotic cell death inducer staurosporine was used as a positive control. Similar results were obtained in 1 µM staurosporine-treated group; the levels of total PARP-1 and caspase-3 were decreased, while the levels of cleavage forms significantly increased compared with the control group (Fig. 3a).
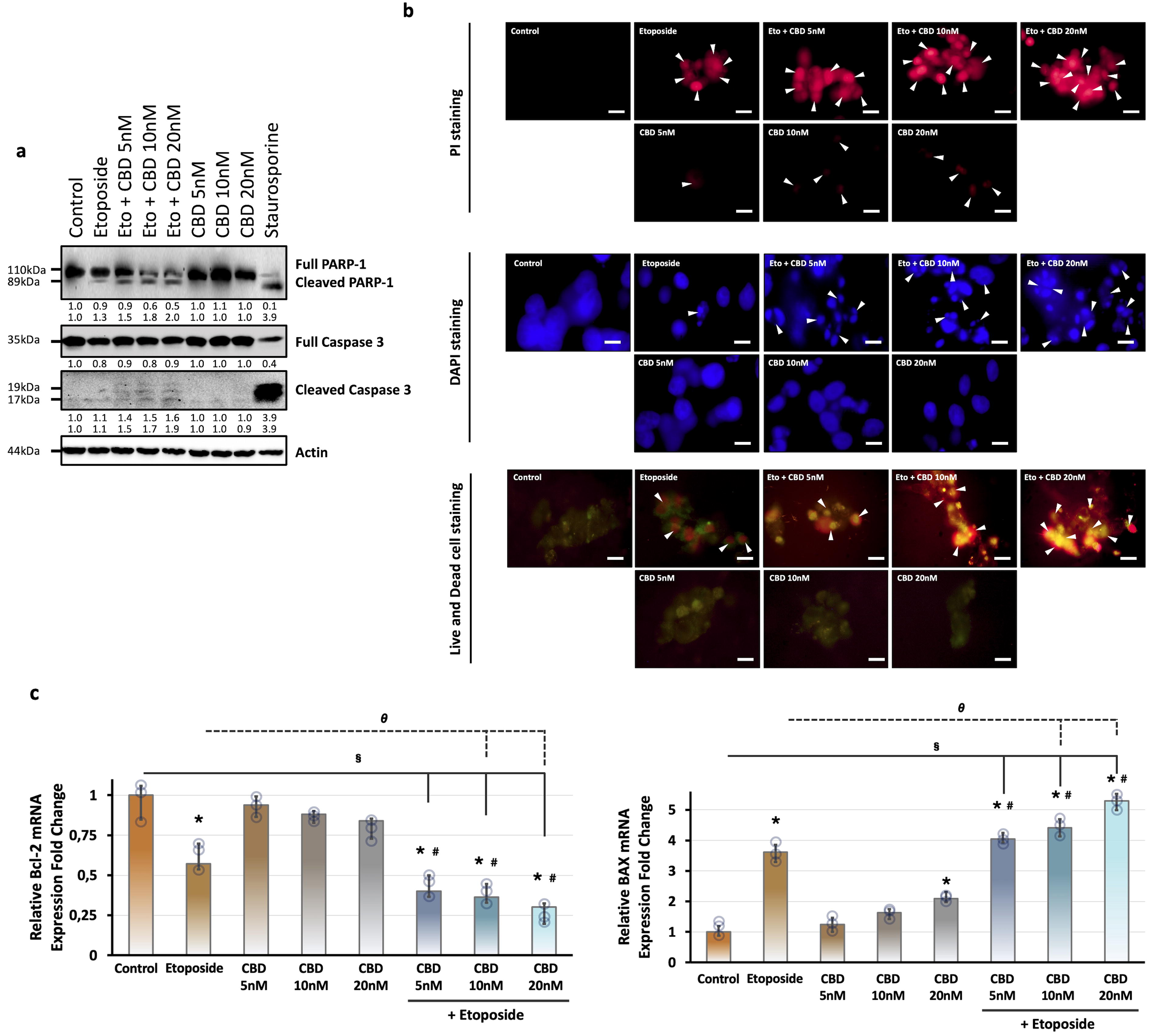
Investigation of the effect of etoposide, cannabidiol, and their combination on programmed cell death in LNCaP cells. Cells were treated with vehicle or 10 μM etoposide and 5, 10, and 20 nM CBD or with their combination for 24 h.
Moreover, we investigated the nuclear formation of LNCaP cells by using DNA staining dyes, including DAPI and PI. The difference between these methods is that they work with fixed cells in DAPI staining, whereas PI staining was performed as a vital staining. Also, the viability status of LNCaP cells was evaluated by a fluorescent-based method, the live
Additionally, we evaluated the proapoptotic gene BAX and antiapoptotic gene BCL-2 mRNA expression levels in LNCaP cells by qRT-PCR. Etoposide treatment significantly induced the mRNA expression levels of BAX and reduced the BCL-2 mRNA levels compared with the control group (Fig. 3c). Only 20 nM CBD treatment significantly increased the BAX mRNA levels. Also, CBD treatment did not significantly affect the mRNA level of BCL-2 compared with the control group (Fig. 3c). Cotreatment of CBD and etoposide significantly decreased the BCL-2 levels and also importantly increased the mRNA level of BAX in a dose-dependent manner compared with the etoposide-treated group alone (Fig. 3c).
Cannabidiol remarkably improves the anticancer effect of etoposide on tumorigenic protein levels in prostate cancer cells
Epithelial-mesenchymal transition (EMT) is one of the hallmarks of cancer. 49 To evaluate the effects of CBD on the EMT process, we tested the basal levels of EMT-related proteins, such as N-cadherin and E-cadherin by immunoblotting. CBD treatment slightly increased E-cadherin and decreased N-cadherin protein levels in a dose-dependent manner (Fig. 4a). Etoposide administration also increased the E-cadherin levels and decreased the level of N-cadherin protein (Fig. 4a). The combined treatment of CBD with etoposide significantly increased the basal levels of E-cadherin and more strongly decreased the N-cadherin levels (Fig. 4a).
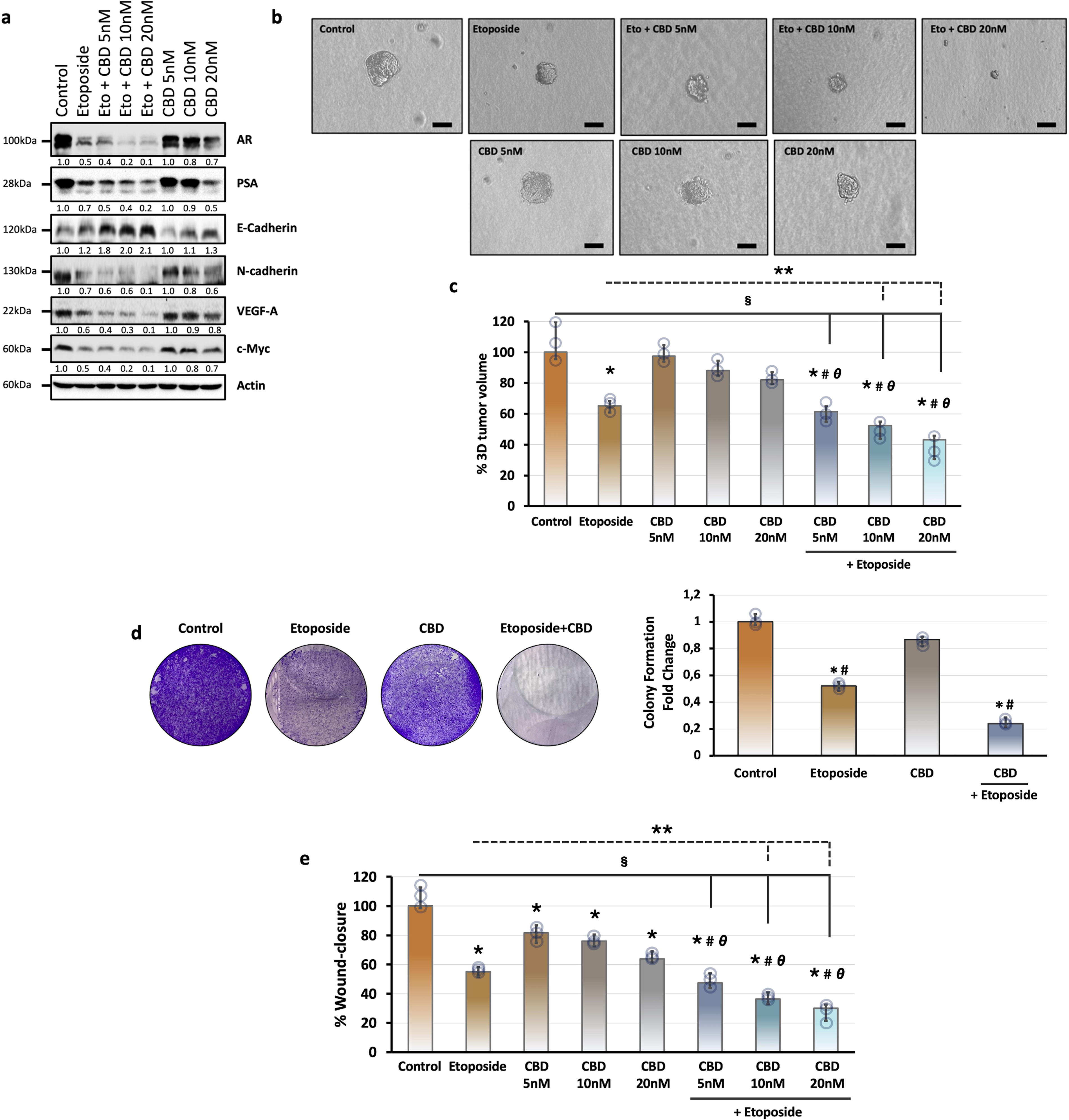
Evaluation of the effect of etoposide, cannabidiol, and their combination on prostate carcinogenesis-related proteins and tumorigenic features of LNCaP cells.
Angiogenesis is another important process in cancer progression and it is regulated by both activator and inhibitor molecules. Vascular endothelial growth factor (VEGF) is one of the identified angiogenesis activator groups of proteins. 50 Our results indicated that CBD treatment alone led to a slight decrease in the level of VEGF-A protein compared with the control group, while the combined treatment with etoposide strongly decreased the level of VEGF-A compared with the etoposide-treated group (Fig. 4a). Etoposide administration alone also decreased the expression level of VEGF-A protein compared with the control group (Fig. 4a).
We also tested the levels of c-Myc, which is one of the proto-oncogenic transcription factors involved in the proliferation, chemoresistance, and also reprogramming of multiple types of cancer cells, including prostate cancer. 51 Etoposide alone decreased the protein levels of c-Myc. CBD treatment alone also slightly decreased the c-Myc levels, whereas its combination with etoposide importantly decreased the level of c-Myc protein compared with the CBD-treated group in a dose-dependent manner (Fig. 4a).
Coadministration of cannabidiol with etoposide markedly reduces the androgenic signaling protein levels and also inhibits the 3D tumor formation
Androgen receptor (AR) and AR-mediated transcriptional programs have a crucial role in the pathogenesis of prostate cancer. Androgenic signaling is also the main target for therapeutic strategies against prostate cancer and alterations in the androgen signaling modify resistance against therapeutic approaches. 52 To test the effect of CBD on androgenic signaling, we investigated the protein levels of AR and prostate-specific antigen (PSA). CBD administration dose-dependently reduced the AR and PSA levels compared with the control group (Fig. 4a). Etoposide alone also decreased the expression level of AR and PSA (Fig. 4a). Cotreatment of CBD and etoposide markedly decreased the AR and PSA levels dose-dependently (Fig. 4a).
In addition, we examined the possible improvement effect of CBD on etoposide by 3D cell culture model. Etoposide treatment importantly reduced the tumor volume and also CBD administration slightly decreased the 3D tumor formation of LNCaP cells (Fig. 4c). Our results indicated that combined administration of CBD with etoposide exhibited a more effective inhibitor effect on 3D tumor formation of prostate cancer cells in a dose-dependent manner compared with etoposide or CBD treatment alone (Fig. 4c).
Cannabidiol remarkably improves the antitumorigenic properties of etoposide on migration, invasion, and colonial growth of prostate cancer cells
To evaluate the enhancer effect of CBD on etoposide in prostate cancer cells, we treated the LNCaP cells with etoposide and its combination with various doses of CBD and then tested the migration, invasion, and colonial growth features of LNCaP cells by wound-healing assay, Matrigel-coated Boyden chamber assay, and 2D plate colony formation assay, respectively. Our findings indicated that etoposide and 5, 10, and 20 nM CBD administration alone significantly limited the tumorigenic features of LNCaP cells in a dose-dependent manner compared with the control group (Fig. 4d, e). Coadministration of etoposide and CBD as indicated doses more strongly reduced the migration, invasion, and colonial growth of LNCaP cells (Fig. 4d, e). Altogether, these results indicated that cotreatment of CBD with etoposide importantly improved the anticancer properties of etoposide on tumorigenic features of LNCaP cells.
Cannabidiol strongly increases the etoposide-induced senescence in prostate cancer cells
Cellular senescence is an important marker of aging. To examine whether CBD supports the etoposide-induced cellular senescence, we treated the LNCaP cells with etoposide and its combination with CBD for 24 h, and then cellular senescence was investigated by senescence-associated β-galactosidase assay. Our findings indicated that etoposide-treated cells exhibited a large, extended, and flattened cell shape typical of senescence and the cells were significantly stained with SA-β-gal compared with the control group (Fig. 5a). CBD administration alone slightly increased the SA-β-gal staining in a dose-dependent manner compared with the control group. Coadministration of CBD and etoposide triggered the significant increase of SA-β-gal staining in a dose-dependent manner compared with etoposide or CBD-treated groups alone (Fig. 5b).
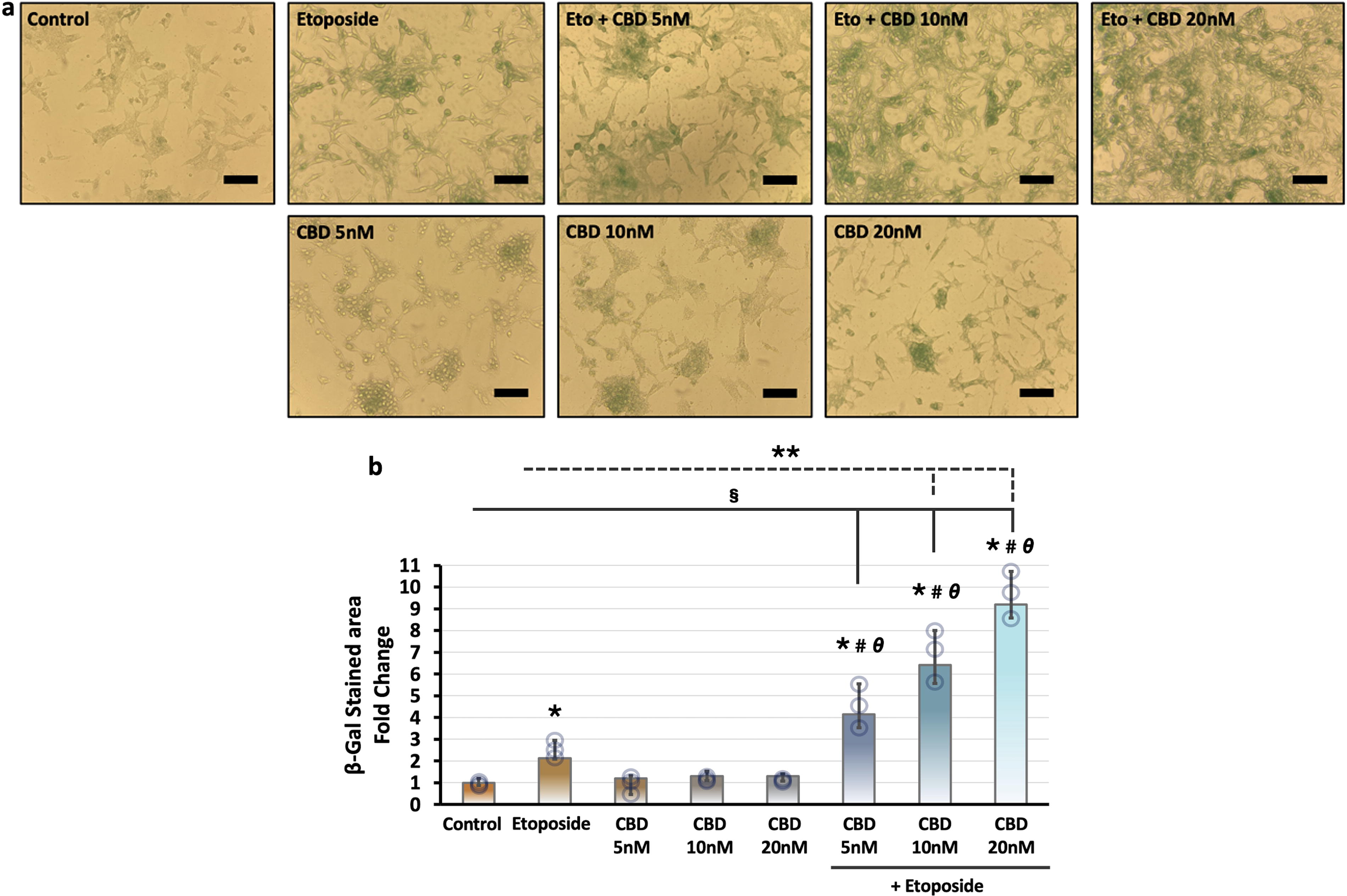
Evaluation of the effect of etoposide, cannabidiol, and their combination on cellular senescence in LNCaP cells.
Discussion
Cannabinoids are specific compounds found in Cannabis sativa and since ancient times they have been used in modern medicine and Chinese medicine.53–56 Several studies have focused on its anti-inflammatory, antioxidant, antipsychotic, and analgesic properties.57–61 The study on colorectal carcinoma cells showed a chemopreventive effect of CBD by reducing cell proliferation. 61 Another study reported that CBD inhibited the viability of human prostate cancer cells, stimulated apoptosis, and elevated reactive oxygen species. 62
Over the past years, many chemotherapeutics have been developed and commonly used in the treatment of numerous cancer types. 63 Today, there are 34 drugs approved for prostate cancer treatment. 64 However, acquired resistance to chemotherapeutics leads to failure in the therapeutic efficacy of the drug in 90% of patients with metastatic cancer. 65 Novel studies have been focused on the potential anticancer effects of natural compounds. Combinatory drug therapies against cancer are a new wave of treatment and are thought to offer more effective treatment approaches. A recent study reported that CBD strongly reduced the endogenous AR expression and limited the nuclear translocation of AR in human androgen-dependent prostate cancer cells. 21 Similarly, etoposide was reported to disrupt androgenic signaling in prostate cancer cells. 66 On the other hand, Cattrini et al. reported that castration-resistance DU145 and PC3 cells were more sensitive than castration-sensitive LNCaP cells. 67 Thus, we turned to test the possible potentiator effect of CBD on etoposide in androgen-dependent prostate cancer cells (LNCaP) for improving the anticancer effects of etoposide. Herein, we aimed to investigate the anticancer effects of CBD and its combination with etoposide in androgen-dependent prostate cancer cells by using various methodologies. Also, one of the important points of the present research is the booster effects of low-concentration CBD applications (5–20 nM) on prostate cancer cells. Li et al. showed that 10 μM CBD significantly reduced survival in PC3 cells. 68 Fisher et al. demonstrated that 0–50 μg/mL CBD administration reduced cell viability of SK-N-SH and NUB-6 neuroblastoma cells in a dose- and time-dependent manner. 69 In another study, performed on A549 human lung cancer cells, examined the effect of CBD doses in the range of 0–120 μM on cell viability, and IC50 values were found to be 37 μM for CBD. 70 The forceful biochemical effects of low-concentration CBD application on prostate cancer cells compared with other cancer cells suggest that it may be an essential tool for the treatment of prostate cancer.
Autophagy maintains homeostasis in eukaryotic cells by providing energy and a sustainable source of biomolecules under stressful conditions, such as hypoxia, oxidative stress, ER stress, or tumor microenvironment.71,72 The relationship between autophagy and cancer is controversial due to the dual role of autophagy in the survival or suppression of tumors based on the status of cancer cells.73,74 Thus, many of the autophagy-targeted cancer therapies remain unsuccessful. The p62/sequestosome 1 (SQSTM1) is a ubiquitin receptor protein that binds cargo directed to autophagic degradation to LC3 when forming autophagosome, and suppression of autophagic flux causes its accumulation. 75 LC3 is a precursor protein and its cleavage by ATG4 leads to phosphatidylethanolamine (PE) conjugation and conversion of LC3-I to LC3-II. 76 Based on the crucial role of autophagy in cancer cells, to understand the effects of CBD and its combination with etoposide on autophagic activity, we tested the levels of p62/SQSTM1 and conversion of LC3-I to LC3-II. According to our results, CBD administration increased levels of p62/SQSTM1 and conversion from LC3-I to LC3-II (Fig. 2a). The combination CBD with etoposide more strongly caused an increase in the levels of p62/SQSTM1 and LC3-I to LC3-II conversion (Fig. 2a). Regarding these results, cotreatment with etoposide even more strongly enhanced CBD-induced autophagic blockage compared with the CBD or etoposide alone in LNCaP cells. Today, modulation of autophagy is among the anticancer mechanisms of natural products and anticancer agents.77,78 Our findings suggest that CBD mediates a more potent anticancer activity by potentiating the decrease in autophagic activity caused by etoposide through modulation of autophagy.
ER has also a pivotal role in the quality control of cellular proteins by the process known as ERAD, and this mechanism maintains the transportation of properly folded and assembled proteins to their final destinations.79–82 UPR, which is an integrated signal transduction pathway, initially acts as a compensatory mechanism to reconstruct ER homeostasis, but under prolonged ER stress, it can trigger cell death by activation of PERK and IRE1α branches excessive UPR stimulation. 83 At this point, the PERK/ATF4/CHOP pathway plays a major role in the direction of the UPR-mediated cellular death process. 47 Herein, to understand the association of the potentiator role of CBD on etoposide with ERAD and UPR signaling in prostate cancer, we tested protein levels of IRE1α and PERK branches by immunoblotting studies. Our findings demonstrated that CBD treatment dose-dependently increased the IRE1α and its effector protein XBP-1s levels (Fig. 2c). Also, CBD importantly increased the levels of PERK and phosphorylation of eIF2α (Fig. 2c). Cotreatment of CBD and etoposide more strongly increased IRE1α and PERK branch-related protein levels (Fig. 2c). These results suggested that CBD, either alone or combined with etoposide, induced ER stress by activating UPR signaling in prostate cancer cells. Present findings suggest that CBD exerts a potentiating effect on etoposide.
To elucidate the effect of CBD, either alone or in combination with etoposide, on one of the tumorigenicity-associated mechanisms, EMT, we evaluated N-cadherin and E-cadherin protein levels. EMT has been associated with increased invasive capacity and metastasis of cancer cells and the development of drug resistance.49,84 Therefore, novel EMT-targeted therapies are thought to be promising in the treatment of various cancer types. 85 As a result of our findings, CBD treatment slightly increased E-cadherin levels, whereas N-cadherin levels were decreased in a dose-dependent manner (Fig. 4a). Moreover, the combination of CBD with etoposide notably inhibited the EMT process in a prostate cancer cell by increasing E-cadherin and decreasing N-cadherin levels (Fig. 4a). Additionally, the effect of combinatory treatment on the levels of oncogenic factors, including c-Myc, PSA, and VEGF-A, was evaluated by immunoblotting. PSA screening is used for prostate cancer diagnosis in clinical practice and a significant increase in PSA levels is seen in prostate cancer patients. 86 Also, previous studies have reported the suppressive effect of CBD on AR signaling in prostate cancer cells. 21 Therefore, suppression of PSA levels with treatment is essential in regressing the cancerization process in prostate cancer cells. Our findings demonstrated that PSA level was reduced by CBD administration in a dose-dependent manner (Fig. 4a) and cotreatment of CBD and etoposide led to a significant decrease in the PSA level (Fig. 4a). Consistent with these results, AR levels were also reduced with the cotreatment compared with CBD or etoposide alone (Fig. 4a). Altogether, our findings supported that CBD potentiated the suppressive effect of etoposide on androgenic signaling.
Angiogenic factor VEGF-A plays a central role in tuning the angiogenic capacity in divergent cancer types. Besides, high metastatic capacity has been observed in VEGF-secreting tumors.87,88 Anti-VEGF treatment strategies that target VEGF pathways in tumors become effective in avoiding tumor progression and are widely used in clinical treatments. 89 Our data demonstrated that CBD treatment led to a slight decrement in VEGF-A protein levels. The combined treatment of CBD with etoposide strongly decreased VEGF-A levels compared with etoposide alone (Fig. 4a). These results mean that CBD has a potentiator activity on the anticancer activity of etoposide.
Elevated oncogenic transcription factor c-Myc expression leads to changes in cell adhesion, metabolic pathways, angiogenesis, and apoptosis. It is also associated with tumor aggression and poor clinical outcomes.90–93 Our immunoblotting results showed that CBD administration slightly decreased the c-Myc level in a dose-dependent manner, whereas cotreatment of CBD and etoposide strongly decreased the c-Myc level (Fig. 4a). These results suggest that the suppressive effect of etoposide on c-Myc can be improved with CBD. Consistent with the effect of coadministration of CBD and etoposide on tumorigenic proteins in prostate cancer cells, functional analysis data, such as colonial growth, migration, and anchorage-independent growth also strongly supported the potentiator role of CBD on etoposide (Fig. 4b–e). Also, CBD notably improved the cellular senescence-inducing activity of etoposide (Fig. 5a).
To test the potential effects of etoposide and CBD on programmed cell death, referred to as apoptosis, we examined the activation of cell death-related proteins, caspase-3 and PARP-1. Caspase-3 is a key mediator for apoptosis and is activated via extrinsic and intrinsic pathways in apoptotic cells.92,94,95 PARP-1 has an important role in DNA replication, transcription, and cell death and it is one of the proteins that caspases have a role in the cleavage and inactivation. 96 Our results showed that CBD treatment at the concentrations we used did not alter the levels of the cleavage forms of caspase-3 (17, 19 kDa) and PARP-1 (89 kDa) proteins. On the other hand, the coadministration of CBD and etoposide has boosted the etoposide-mediated reduced total caspase-3 and PARP-1 levels while increasing cleavage forms of caspase-3 and PARP-1 (Fig. 3a). Consistent with these findings, coadministration significantly increased the mRNA level of apoptotic gene BAX and decreased antiapoptotic BCL-2 compared with CBD or etoposide alone (Fig. 3c). These results confirmed the potentiator role of CBD on etoposide in prostate cancer cells.
Within the scope of our studies, we tried to evaluate the potentiator effects of CBD on the anticancer activity of etoposide through limited signaling pathways. Some of the underlying reasons for these effects of CBD may be its effects on common molecular targets with etoposide. Etoposide inhibits the topoisomerase II enzyme by stabilizing the enzyme–DNA complex. 97 CBD oxidation product HU-331 has a potent inhibitory activity against human topoisomerase II, whereas CBD does not have a strong effect on the topoisomerase II enzyme.98,99 In our coadministration studies, one of the mechanisms underlying the enhancing effect of CBD on the anticancer activity of etoposide may be its strengthening of topoisomerase II enzyme interaction with etoposide. Moreover, CBD may cause an increase in intracellular levels of etoposide by affecting the levels of transporter proteins. The therapeutic efficacy of etoposide is limited due to low solubility and poor bioavailability levels. 100 Previous studies have reported that CBD could alter P-glycoprotein (P-gp) and breast cancer-resistant protein (BCRP) expression and inhibit P-gp efflux function in BeWo, Jar, and MCF/P-gp cells. 101 Nabissi et al. reported that CBD increases transient receptor potential cation channel subfamily v member 2 (TRPV2) expression and activity in glioblastoma cells and CBD increases drug uptake by increasing TRPV2-dependent Ca2+ influx, thus improving anticancer drug efficiency. 102 Fraguas-Sánchez et al. reported that CBD-loaded microparticles potentiated the anticancer effect of paclitaxel (PTX) and doxorubicin (DOX) in MCF-7 and MDA-MB-231 breast cancer cells. 103 Considering the previous reports, it is suggested that CBD exerts a potentiator effect on anticancer compounds by possibly contributing to the intracellular uptake of chemotherapeutic agents. Moreover, Li et al. revealed that CBD may potentiate the anticancer effect of doxorubicin in a synergistical manner in osteosarcoma. 22 In addition, Hao et al. reported that CBD protects the heart tissues against DOX-induced cardiomyopathy. 104 In addition, CBD potentiates the oxaliplatin parental and oxaliplatin-resistant human colorectal cancer cell lines and their mouse xenografts and also cisplatin toxicity in endometrial cancer cell lines. 23 Supporting current studies, our findings also revealed that CBD exhibited anticancer effects on prostate cancer cells by exerting effects on physiologically essential mechanisms, such as UPR, ERAD, and autophagy. Moreover, it also exhibits enhancing effects on the etoposide due to its suppressive properties on EMT, angiogenesis, and androgenic signaling in prostate cancer cells.
Present in vitro results suggest that CBD has potentiating effects on etoposide, an effective chemotherapeutic agent in prostate cancer cells. However, for the usage of CBD in cancer treatment as a potentiator to be reflected in clinical applications, in vivo studies need to be carried out, and the dose
Conclusion
Overall, the present study data suggest that CBD administration, in addition to etoposide, significantly improves the anticancer activity of etoposide in prostate cancer cells. The acceleration of advanced characterization studies to elucidate the molecular mechanism of action of CBD on cancer cells suggests that it may offer an effective adjuvant approach to increase the efficacy of existing chemotherapeutics.
Footnotes
Acknowledgments
The authors thank Suleyman Demirel University-Innovative Technologies Application and Research Center (YETEM). They thank Dr. Ozlem Ozmen (Department of Pathology, Faculty of Veterinary, Mehmet Akif Ersoy University) for allowing them to access the use of fluorescence microscope. They also thank Erzurumlu Laboratory member, Hatice Kubra Dogan, for assisting with some experiments.
Authors’ Contributions
Y.E. initiated and directed the project. Y.E. designed and conducted the experiments, analyzed and interpreted the results, and wrote the article. D.C. assisted experimental studies. All correspondence and requests for materials should be addressed to Y.E. All authors have given the final approval of the submitted article.
Data Availability
The datasets are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare no competing financial interests.
Funding Information
This study was partially supported by Suleyman Demirel University internal funds (
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.