
Abstract
Introduction:
Early life is a sensitive period for brain development. Perinatal exposure to cannabis is increasingly linked to disruption of neurodevelopment; however, research on the effects of cannabidiol (CBD) on the developing brain is scarce. In this study, we aim to study the developmental effects of neonatal CBD exposure on behavior and dendritic architecture in young adult rats.
Materials and Methods:
Male and female neonatal Sprague Dawley rats were treated with CBD (50 mg/kg) intraperitoneally on postnatal day (PND) 1, 3, and 5 and evaluated for behavioral and neuronal morphological changes during early adulthood. Rats were subjected to a series of behavioral tasks to evaluate long-term effects of neonatal CBD exposure, including the Barnes maze, open field, and elevated plus maze paradigms to assess spatial memory and anxiety-like behavior. Following behavioral evaluation, animals were sacrificed, and neuronal morphology of the cortex and hippocampus was assessed using Golgi–Cox (GC) staining.
Results:
Rats treated with CBD displayed a sexually dimorphic response in spatial memory, with CBD-treated females developing a deficit but not males. CBD did not elicit alterations in anxiety-like behavior in either sex. Neonatal CBD caused an overall decrease in dendritic length and spine density (apical and basal) in cortical and hippocampal neurons in both sexes. Sholl analysis also revealed a decrease in dendritic intersections in the cortex and hippocampus, indicating reduced dendritic arborization.
Conclusions:
This study provides evidence that neonatal CBD exposure perturbs normal brain development and leads to lasting alterations in spatial memory and neuronal dendrite morphology in early adulthood, with sex-dependent sensitivity.
Introduction
The use of cannabinoids has increased significantly in the past two decades in the United States concurrent with the decriminalization and legalization of cannabis for medicinal and recreational uses in multiple states. While this has been largely perceived by the public as a net positive and a sign of societal progress,1,2 there are public health concerns regarding the long-term use of these compounds and how they could affect development for vulnerable populations.
Brain development occurs both in utero and over the course of childhood with highly regulated and evolutionarily conserved systems.3–6 There are countless examples of exogenous factors, which can influence these systems, from environmental stresses to toxic exposures,7–18 which have varying consequences and distinct windows of vulnerability.
The endogenous cannabinoid (endocannabinoid) system plays important roles in brain development that span in utero time through childhood. The endocannabinoid system regulates neurogenesis, gliogenesis, neuronal migration, synaptogenesis, and synaptic pruning. 19 Importantly, the timing of these processes are different in males and females, which may lead to sex differences in temporal susceptibilities.20–22
Cannabidiol (CBD) and delta-9-tetrahydrocannabinol (THC) are the two major phytocannabinoids present in cannabis. Unlike THC, CBD does not produce euphoric or perceptual changes and is generally perceived as safe. The use of CBD among pregnant women has increased 106% from 2003 to 2016, with 7% of pregnant women reporting its use.23,24 Nonetheless, CBD does have neurological effects. A Food and Drug Administration (FDA)-approved oral formulation of CBD (Epidiolex) is currently used to treat a rare form of childhood epilepsy, demonstrating CBD’s interaction with the brain and modulation of synaptic activity, although the precise mechanism of the drug’s effects has not yet been elucidated. 25
To study the long-term effects of CBD on brain development and cognition, we established an animal model of developmental neurotoxicity to assess behavior and neuronal morphology. Neonatal male and female Sprague Dawley rats were exposed to CBD or vehicle and evaluated using a spatial memory and anxiety measurement tasks. We analyzed neurons from the cortex and hippocampus to quantify ultrastructural changes in dendritic properties that could account for behavior effects. These regions have a high density of cannabinoid receptors,26,27 are a primary site of action for cannabinoids, and are critical for spatial memory retrieval. 28
Materials and Methods
Animals
Procedures were conducted within an American Association for Accreditation of Laboratory Animal Care-accredited animal facility at UCSF and according to Institutional Animal Care and Use Committee-approved protocols. Mixed-sex (5 male/5 female) Sprague Dawley rat litters with one dam per cage were purchased from Charles River Laboratory, South San Francisco, CA, USA. Animals arrived at the age of postnatal day (PND) 1 and were kept under standard animal housing conditions with a reversed 12-h light–12-h dark cycle (lights ON at 08:00 pm). Animals had access to standard laboratory food and water ad libitum. At PND 21, animals were weaned, separated by sex, and housed in groups of 3–4/cage.
Drug preparation and administration
CBD was purchased from Cayman Chemical (Ann Arbor, MI) with ≥98% purity. CBD was dissolved in sesame oil 29 with vigorous vortex. CBD was intraperitoneally administered to neonate pups at PND 1, 3, and 5 at 50 mg/kg (volume 0.1 mL/10 g of body weight). The vehicle group was administered an equal volume of sesame oil. Treatment was assigned randomly by litter to eliminate the chance of contamination of control animals with CBD exposure. Experimental groups thus included animals from the same litter as well as different litters minimizing any litter effects.
Body and brain weight measurement
Rats were weighed from arrival (PND 1) until sacrifice (PND 57) at weekly intervals. Brains were weighed after sacrifice. Brain index was calculated as brain weight divided by body weight (g). 30
Behavioral assessments
Animals were habituated to the experimenter and experimental conditions before testing. Tests were started at PND 42 and completed during the dark cycle. Red light illumination was used for the experimentalist behind the screen of the testing arena. Standard white lights were used to illuminate the behavioral paradigms (Barnes maze [BM], open field test [OFT], and elevated plus maze [EPM]) to facilitate video recording and provide a small negative stimulus. Behavioral testing was divided into cohorts to accommodate testing all animals within the same day, within their active phase. Each cohort had groups consisting of 2–3 litters. The BM was completed first, with 5 days of consecutive training, followed by 1 week of no training or testing, then the probe trial. The OFT and EPM were performed sequentially, with at least 1 day of nontesting between the tests. Animal movements were tracked and recorded using a GigE camera (Basler Inc; Exton PA), and EthoVision XT 11.5 software (Noldus; Leesburg VA).
BM test
The BM test was conducted as previously described31,32 (Supplementary Fig. S1A). Rats underwent one training trial per day (4 min time limit). One week after completion of training, memory retrieval was tested in a probe test in which the rat was allowed to explore the maze without the escape box for 90 sec.
OF test
The OFT arena was divided into an outer and inner zone (Supplementary Fig. S1B). The test was performed as previously reported. 32 The rat was allowed to explore the OFT arena for 5 min.
EPM test
The EPM apparatus consists of two open and two closed arms that are perpendicular to each other (Supplementary Fig. S1C). The test was conducted as previously described. 32 The rat was placed in the center of the apparatus and allowed to move freely for 5 min.
Sample collection
After behavioral evaluation, animals were anesthetized deeply with isoflurane and perfused transcardially with 0.1 M phosphate-buffered saline (PBS). The whole brain was extracted and rinsed with PBS.
Golgi–Cox staining
Golgi–Cox (GC) staining was used to label neurons in the brain and identify dendrite structure with slight modifications. 33 Briefly, the brains were placed in glass vials containing GC solution for 2 weeks in the dark at 4°C, exchanged with the fresh GC solution weekly. Brains were sectioned at 150 µm, washed in PBS, mounted on slides, and dried overnight at room temperature in the dark, followed by slide development with 75% ammonia solution and 5% sodium thiosulfate. The slides were dehydrated with increasing concentrations of alcohol, cleared with xylene, and mounted with DPX.
Neuronal morphology study
Image acquisition
A Nikon Eclipse 80i microscope (Nikon; Melville, NY) with a Hamamatsu camera was used for image acquisition. We studied the dendritic morphology of cortical pyramidal neurons (layer III/IV) (+1.5 mm to −4 mm anterior to posterior of Bregma). Neurons from the hippocampus in regions DG/CA1/CA3 were analyzed from both dorsal (−3 mm to −5 mm posterior of Bregma) and ventral (−5 mm to −6 mm posterior of Bregma) hippocampus. A 20× objective was used to acquire image stacks (step-size 0.5 µm). For spine density analysis, dendritic segments in the region of interest (ROI) were scanned using a 100× oil-immersion objective. Six representative neurons from each ROI per animal were selected.
a. Neuronal tracing
The selected neurons were traced in three dimensions (3D) using Neurolucida software (MBF Bioscience Inc., Williston, VT) to identify soma, axon, and apical and basal dendrites.
b. Dendrite arborization
Neuronal tracing was visualized and analyzed using the NeuroExplorer component of Neurolucida. Dendritic length was measured. Sholl analysis 34 was conducted by measuring the intersections with each concentric ring (10 µm apart) from the soma. Branches beyond 300 µm were excluded to avoid analyzing incomplete branches. To quantify spine density, ten dendrites from each dendrite were randomly selected. Dendritic spines were counted using a cell counter plug-in of ImageJ-win64.
Statistical analysis
Group size was determined with a power analysis using G*Power 3.1 software (Dusseldorf, Germany), using results from previous studies utilizing early-life isoflurane or diazepam exposure and BM as the primary behavioral outcome.22,32,35 The calculated effect size fell within the range of 0.7 to 0.9, with a difference in means ranging from 17 to 25 and standard deviation from 24 to 27. With a significance level (α) of 0.05 and a power of 0.8, we determined that a sample size of 15–25 animals per group was needed.
Data are expressed as mean ± standard errors of the mean (SEM). Body weight was analyzed with a repeated measure three-way analysis of variance (ANOVA) and Tukey’s post hoc test. BM training was analyzed with a mixed-effects model with Tukey’s multiple comparison test (MCT). BM probe trial was analyzed with a curve fit (nonlinear regression) to test the pattern of exploration from the goal as either a linear or one-phase decay function. A one-phase decay suggests that the group learned the position of the escape hole, spending less time the further the positions are from the goal. A linear function suggests no preference to a particular position in the maze for the group and a lack of memory for the escape hole. In addition, a Dunnett’s MCT was performed per group. Two-way ANOVA with Sidak’s MCT was used to assess brain index, anxious behavior, dendrite length, spine density, and dendritic intersections of GC-labeled neurons. Area under the curve (AUC) was compared with one-way ANOVA. Data were analyzed using GraphPad Prism 9.2.0 for Windows, GraphPad Software, San Diego, CA, USA. Significant differences are shown as *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001.
Mortalities or animals excluded during behavior led to differences in cohort sizes. Five male and three females died after CBD injection. During BM, animals that never reached the escape box during training were excluded from the probe trial analysis, as they never demonstrated acquisition of the task; Male/Vehicle (n = 2); Male/CBD (n = 3); Female/Vehicle (n = 1); and Female/CBD (n = 1).
Results
The experimental schema is presented in Fig. 1.
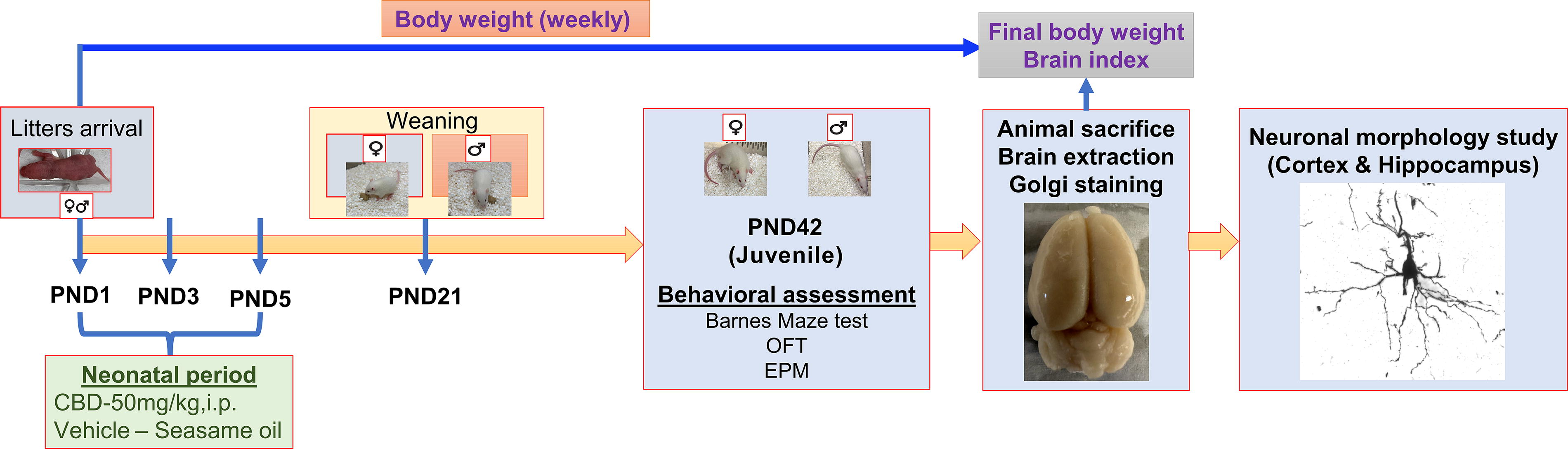
Experimental design. Study timeline illustration of the developmental neurotoxicity model.
Neonatal CBD exposure impairs spatial memory in female rats
All groups learned the location of the escape box over the training period, as evidenced by progressive decreases in latency (F(4,324) = 53.30, p < 0.0001), number of errors (F(4,324) = 13.03, p < 0.0001), and total distance traveled (F(4,324) = 5.890, p = 0.0001). Fecal boli count during the training sessions decreased for all groups over the training period (F(4,324) = 27.53, p < 0.0001), indicative of decreasing anxiety with repeated exposure to the context. There were no significant interactions between treatment, sex, and training days for latency (p = 0.7869), errors (p = 0.6724), distance traveled (0.5583), or fecal boli count (p = 0.6527) (Fig. 2A–D).
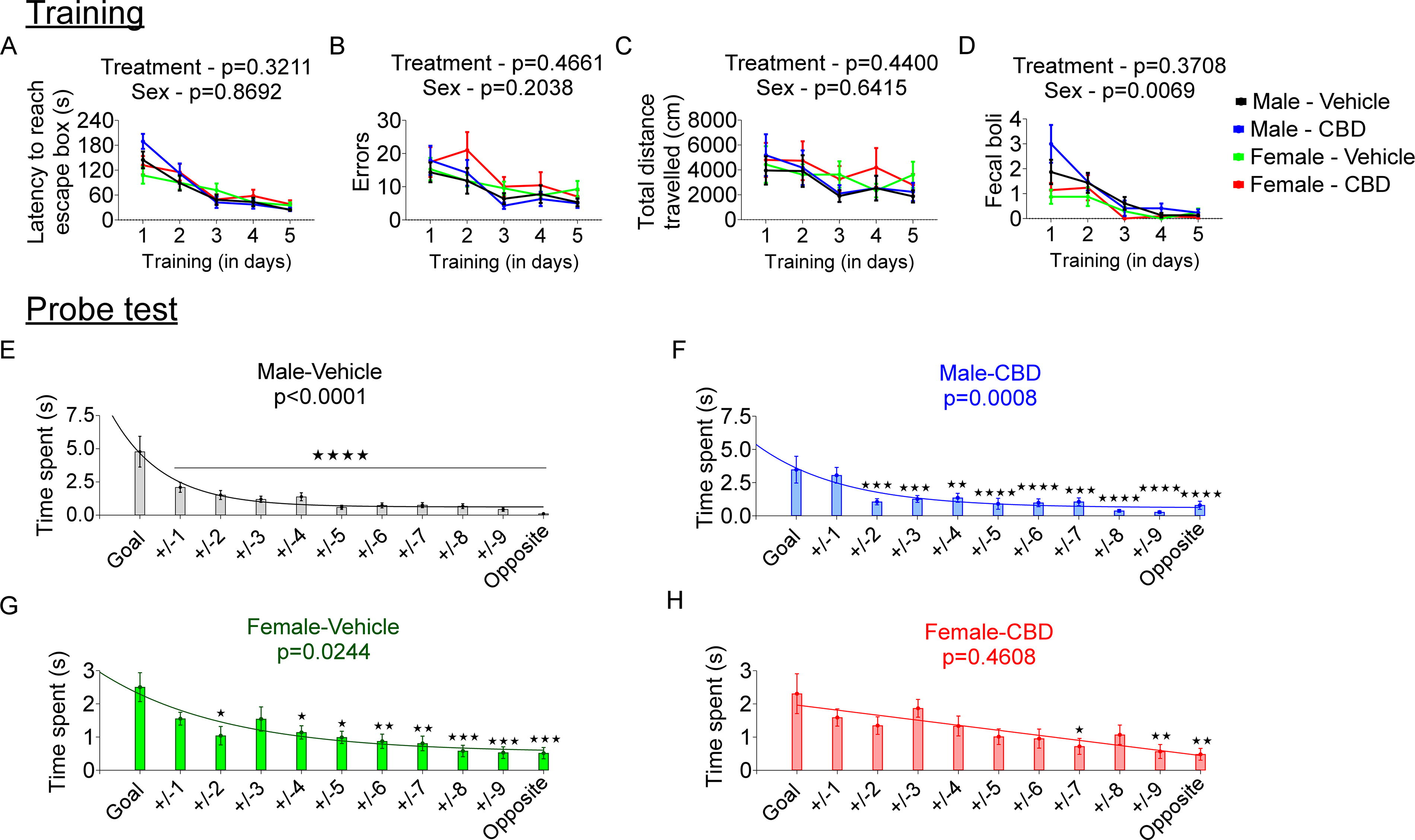
The effect of early neonatal CBD exposure on spatial learning and memory in young adult male and female rats. Spatial learning evaluation during the training phase:
In the probe trial, the Male/Vehicle (F(1,250) = 29.62, p < 0.0001), Male/CBD (F(1,184) = 11.53, p < 0.0008), and Female/Vehicle (F(1,261) = 5.124, p < 0.0244) groups, all differentiated the goal position from the other averaged positions as evidenced by the 1 phase decay pattern by curve fit analysis (Fig. 2E–G). Multiple comparison testing also shows significant time differences from most of the positions in the maze compared with the goal position for these three groups. In contrast, the Female/CBD exposed group did not differentiate the goal position from other positions as evidenced by the flat line of the curve fit (F(1,228) = 0.5459, p = 0.4608) (Fig. 2G).
Neonatal CBD administration does not alter anxiety-like behavior in young adult rats
Given the association of cannabinoid use with anxiety-like behaviors,29,36 we subjected animals to the OFT and EPM. In OFT, male rats spent more time in the inner zone than females (F(1,70) = 5.043, p = 0.0279). However, neonatal CBD administration did not change the proportion of time spent in the inner zone of the OFT arena in males (p = 0.3072) or females (p = 0.9922) (Supplementary Fig. S2A). Similarly, in the EPM, CBD-treated animals performed similar to the vehicle groups (p = 0.4660). There was no difference across sex (p = 0.4755), nor interaction (sex × treatment: p = 0.5632) (Supplementary Fig. S2B).
Neonatal CBD decreases dendritic complexity in young adult rats
Spine density analysis of apical and basal dendrites in the cortex and three regions of the hippocampus (DG, CA1, and CA3) revealed a consistent reduction associated with CBD treatment (Fig. 3; Supplementary Table S1). There were no effects of sex, but spine density in the cortex showed a significant effect of interaction between sex and treatment (Supplementary Table S1).
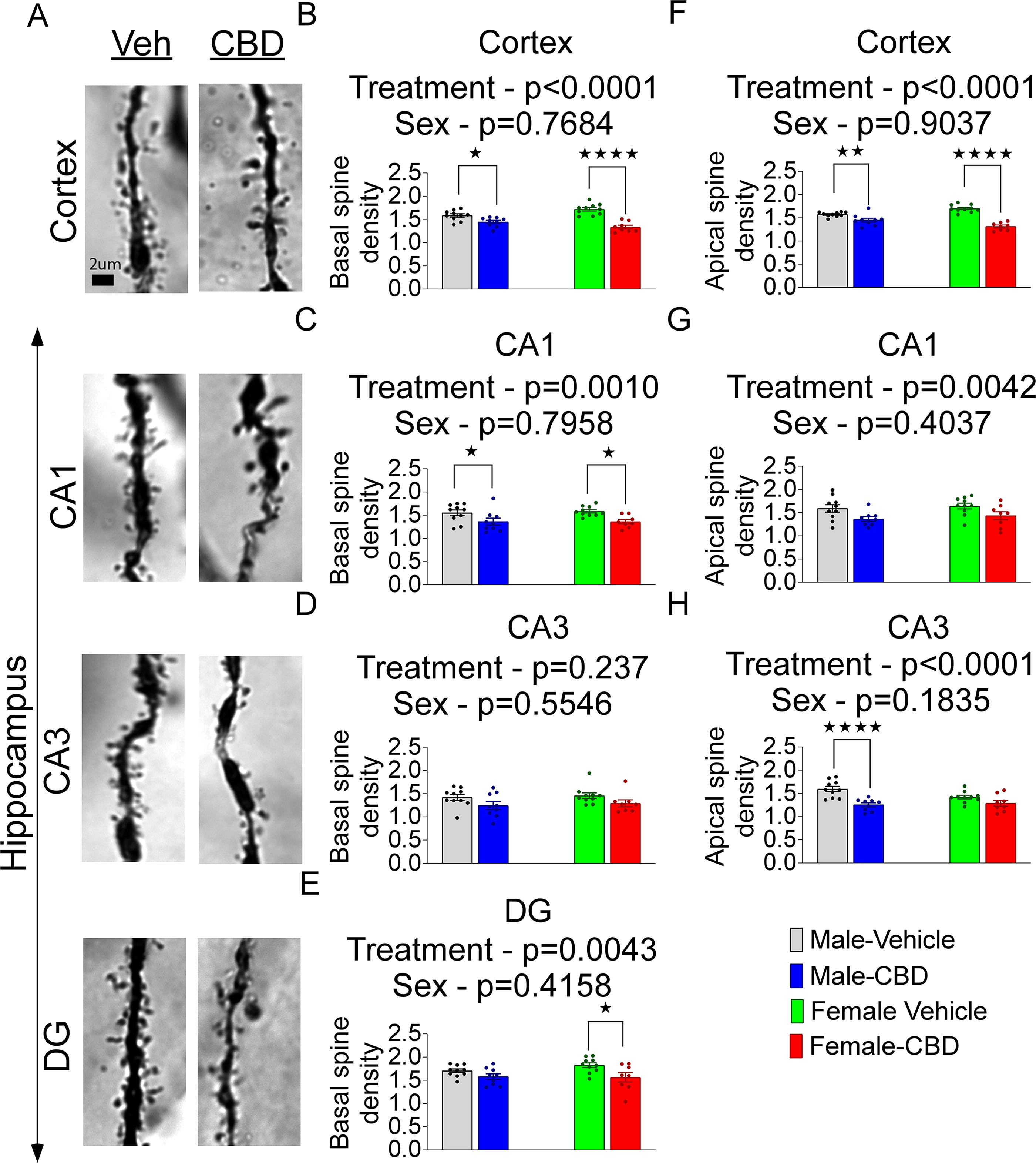
Spine density of basal and apical dendrites in cortex and hippocampus after neonatal CBD administration in males and females.
Next, the apical and basal dendrite length in cortical and hippocampal neurons were measured. Neonatal CBD exposure decreased basal dendritic length in the cortex (F(1,33) = 9.009, p = 0.0051) but hippocampal regions were not different between control and treatment (Fig. 4; Supplementary Table S2). The apical and basal dendritic length of the cortex was negatively affected by sex, (SexApical: F(1,33) = 5.570, p = 0.0243; SexBasal: F(1,33) = 4.700, p = 0.0375). However, sex-specific CBD-associated changes in dendritic length were nonsignificant in most ROI (Fig. 4B–H, Supplementary Table S2).
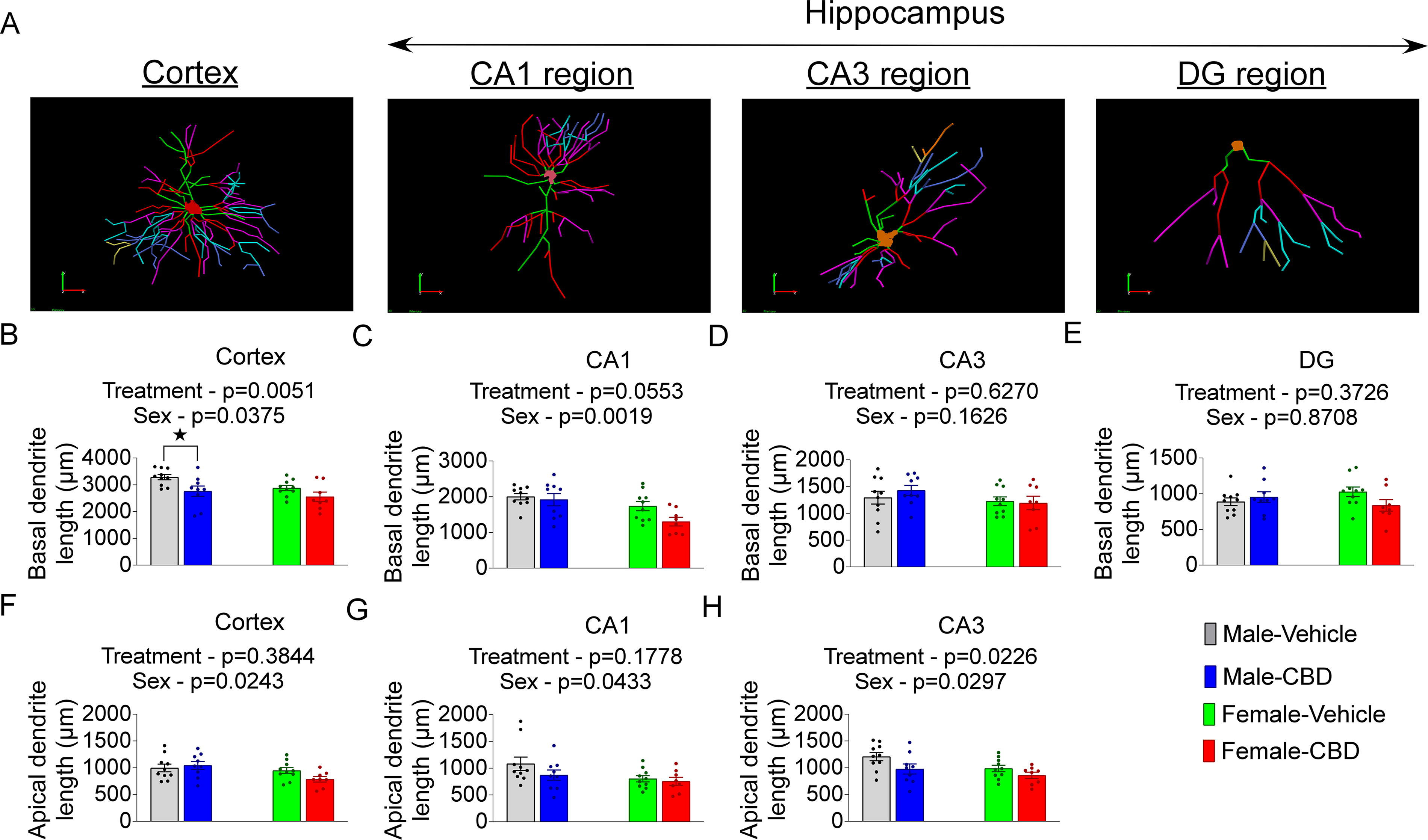
The effect of neonatal CBD administration on dendrite length in young adult males and females.
We performed a Sholl analysis, which revealed alterations in dendritic arborization by measuring the number of intersections between dendritic processes and invisible concentric spheres surrounding the soma (Fig. 5A). Neonatal exposure to CBD decreased the number of dendritic intersections with increasing radial distance in adult cortical, CA1, and DG neurons (Supplementary Table S3). The pattern of arborization differed across treatment groups at multiple radial distances, with decreased dendritic intersections in the cortex of CBD-treated male rats and the CA1 of CBD-treated females compared with vehicle groups (Fig. 5B–C). Neonatal CBD did not decrease dendritic intersections in CA3 neurons (Fig. 5D).

Sholl analysis of intersections in male and female young adult rats after neonatal CBD administration.
To further understand the overall impact of CBD on dendritic complexity, we calculated AUC of the Sholl analysis. CBD exposure was associated with a decreased AUC in the cortex (F(1,33) = 8.160, p = 0.0074). There was an effect of sex for cortex (F(1,33) = 8.253, p = 0.0071), CA1 (F(1,33) = 13.28, p = 0.0009), and CA3 (F(1,33) = 4.645, p = 0.0385), indicating a general reduction of dendritic intersections in females for those regions (Fig. 5F–H). Other regions were not significantly altered by CBD or sex (Supplementary Table S4, Fig. 5G-1).
Neonatal CBD mediates physiological changes throughout development in rats
There were significant differences in body weight among groups by sex (F(1,387) = 1719, p < 0.0001) as well as treatment (F(1,405) = 93.36, p < 0.0001) (Supplementary Fig. S3A), with neonatal CBD associated with increased weight gain into early adulthood. Brain index was calculated at the end of the behavioral experiment and showed no differences by treatment (Supplementary Fig. S3B).
Discussion
The present study demonstrated the impact of early-age exposure to CBD on cognition and morphology of cortical and hippocampal neurons in young adult male and female rats. We found that neonatal CBD exposure impaired spatial memory in females but not males and did not affect anxiety-like behavior in either sex. Neonatal CBD decreased dendritic complexity in both sexes in the cortex and hippocampus. These findings highlight the lasting effects of neonatal CBD exposure in rats, including sex-specific behavioral and neuroanatomical alterations, and underscore the vulnerability of the developing brain to cannabinoid exposure. Notably, CBD is widely perceived as safe compared with other drugs due to its natural origins and nonpsychoactive profile, despite a lack of research on its developmental effects. While further studies are needed to define the underlying mechanisms and identify dosing thresholds, our findings support that extreme caution is warranted when considering the use of cannabis or cannabinoids, including CBD, during pregnancy or while breastfeeding. 37
Our finding that neonatal CBD exposure impairs spatial memory in early adulthood illustrates the long-term effects of cannabinoid exposure on cognition. Importantly, only CBD-treated females exhibited impaired spatial memory, highlighting the influence of sex on outcome determination after cannabinoid exposure. This behavioral sex difference after developmental CBD exposure is in line with previous studies that have reported sex-specific effects of perinatal CBD, although the exposures spanned in utero development and used different behavioral assessments.38,39 While heightened anxiety can impact performance on memory tests, this was unlikely the cause of our finding, as we observed similar performance of all groups on paradigms designed to assess anxiety-like behavior, regardless of CBD treatment.
Spatial memory processing is dependent on the hippocampus and cortex,40,41 which undergo cannabinoid receptor-mediated changes in development during early postnatal life.42 Hippocampal neurocircuitry exhibits rapid maturation and development of dendrites and dendritic spines during postnatal life.43,44 Dendritic length, spine density, and distribution of dendritic arbors are major determinants of neuronal connectivity, and alterations change the functional properties of the neuronal network, which can impact behavior.45,46 The CBD-induced disruption of dendritic complexity demonstrated in our study represents a lasting loss in synaptic connectivity and disrupted neural circuitry. Taken together, the morphological changes reflect a modification in neuronal remodeling that may underlie altered behavior.
Interestingly, despite morphological changes in both males and females, behavioral impairment was only observed in females. Notably, there were some sex differences between the specific brain regions that displayed alterations in dendritic morphology. CBD decreased spine density only in males in the CA1 and only in females in the CA3 regions of the hippocampus. The CA1 and CA3 have distinct roles in spatial memory processing 47 and disruption of a specific hippocampal subnetwork through the CA3 versus the CA1 is likely to yield distinct effects. Together with the female-specific impairment in spatial memory, these results suggest that the precise brain regions affected by CBD may determine behavioral outcome. Canonical brain development is sexually dimorphic in certain brain regions, with critical developmental events occurring at different rates and timepoints in males and females.48,49 The precise timing of an insult may incur sexually divergent outcomes, dependent on the sex-specific state of development that is disrupted. Our finding that neonatal CBD leads to spatial memory impairment in females, as well as disruption of dendritic morphology, emphasizes the importance of considering age and sex in determining CBD’s effects on the brain.
We also observed an increase in body weight in both sexes associated with neonatal CBD treatment. This change in body weight was especially significant at later ages. Cannabinoids are well known to affect metabolism and influence appetite,50 which may underlie the observed weight gain. It is noteworthy that these changes are observed long after the exposure to CBD in the first week of life suggesting an important role for cannabinoids in setting a developmental trajectory for metabolism that can last into young adulthood.
The dosing regimen employed in this study is specific to our model, yet it does facilitate a comparison with human CBD exposure. For the oral formulation of CBD, Epidiolex, approved by the FDA for treating two childhood epilepsy syndromes, the recommended dosing is up to 20 mg/kg/day. FDA-published preclinical data from three individuals (one healthy and two with seizure disorders) exposed to a daily oral CBD dose of 20 mg/kg/day exhibited plasma levels with a Cmax ranging from 219–330 ng/mL.51 Our prior research has demonstrated the significant impact of route and vehicle type on the drug’s pharmacokinetic properties in rats. With a single intraperitoneal injection of CBD dissolved in oil at 50 mg/kg, we anticipate achieving a similar Cmax range of 293 ng/mL in our rodent model. 37 The oil’s depot function extends the effective exposure of a single injection compared with a singular oral exposure, which is likely dosed twice daily. In this study, we administered three separate injections during the first week of life, mimicking a multiday exposure scenario similar to what a child receiving CBD for epilepsy might undergo.
There are several limitations to our study. First, additional experiments are required for a detailed investigation of the mechanisms underlying the morphological changes and memory impairment after neonatal CBD administration. We hypothesize that structural dendritic alterations may underlie the behavioral outcome, but more research is needed to demonstrate causation. Second, the precise role of sex factors in the generation of the sex-specific outcomes remains to be elucidated. It is possible, for example, that males are similarly susceptible to neonatal CBD, but during a different developmental window or with a different dose, as metabolism of cannabinoids differs between the sexes. Third, we chose to use intraperitoneal injection of CBD to minimize dose variability between subjects. This route is not directly translatable to humans as neonatal human exposure most commonly occurs through oral ingestion, breastfeeding, or secondhand smoke. We chose not to use forced oral gavage for the potential confounding effects of nutrition and also chose not to use a breastfeeding model to ensure equal exposure among treated animals. Future studies focusing on routes of exposure that are more clinically relevant for this age group are needed in determining the full translational impact.
Conclusion
Our data that neonatal CBD exposure affects spatial memory performance and alters cortical and hippocampal neuronal morphology in young adult rats challenge the perception that it is safe to expose the developing brain to CBD during pregnancy or in early childhood. Furthermore, the current data indicate that females are more sensitive to neonatal CBD exposure. While future research is required to identify the underlying mechanisms of the observed outcomes and determine whether these findings are fully translational to humans, our current study provides strong evidence that CBD perturbs brain development during this vulnerable period of early life and leads to lasting consequences.
Footnotes
Acknowledgments
The authors would like to acknowledge additional assistance from Katrina Duong, Deeya Amatya, Isha Soni, and Jason Leong for their help at various times to complete these studies.
Authors’ Contributions
M.W.: Investigation, Methodology, Validation, and Writing—original draft G.A.C.: Conceptualization, Methodology, Funding Acquisition, Supervision, and Writing—review and editing J.M.S.R.: Methodology and Writing—review and editing J.H.: Conceptualization and Writing—review and editing. J.W.S.: Conceptualization, Supervision, Funding acquisition, and Writing—review and editing.
Author Disclosure Statement
The authors have no conflicts of interest to declare.
Funding Information
This work was supported by a grant from the
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.