
Abstract
Objectives:
Pediatric bipolar disorder is characterized by core deficits in mood and executive function and commonly co-occurs with attention-deficit/hyperactivity disorder (ADHD). We aimed to examine response inhibition in this population, as an element of executive function, which, if aberrant, may interfere with learning and information processing.
Methods:
Children (9–18 years) with bipolar I or II disorder (BD, n = 26) and age, gender, and intelligence quotient (IQ) comparable healthy children (HC, n = 22) without any psychopathology were given a standardized Go/NoGo computerized task measuring response inhibition. A whole-brain functional magnetic resonance imaging (MRI) group analysis was performed using statistical parametric mapping software (SPM2) for comparing NoGo to Go epochs.
Results:
There were no statistically significant group differences between groups in age, gender, or ethnicity. The BD group had high rates of co-morbid disorders, including 81% with ADHD, 62% with oppositional defiant disorder (ODD), and 46% with anxiety disorders. This BD group had fewer correct responses on Go (84% vs. 96%, T[46] = 3.35, p = 0.002) and overall (85% vs. 94%, T[46] = 4.12, p = 0.0002) trials as compared to the HC group. However, there were no statistically significant group differences in response inhibition on NoGo trials (p = 0.11). In the NoGo−Go contrast, the BD group showed increased neural activation in the right dorsolateral prefrontal cortex (DLPFC) compared to HC (T[46] = 4.21, p < 0.001).
Conclusions:
During accurate NoGo but impaired Go trial performance, children with BD showed increased right DLPFC activation versus controls, suggesting increased recruitment of executive control regions for accurate response inhibition. Studies relating these results to mood regulation in pediatric BD are warranted.
Introduction
Among several elements of executive functioning originating from frontostriatal regions of the brain, component functions of response inhibition include the ability to withhold a preplanned response, interrupt an already initiated process, avoid interference, and delay a response (Tamm et al. 2002). Consequences of dysfunction in response inhibition include behavioral dyscontrol and impairments in sustained attention, which may manifest in a variety of psychiatric disorders. Although response inhibition improves with maturity level (Tamm et al. 2002), deficits in response inhibition have been consistently observed in adults (Blumberg et al. 2003a; Larson et al. 2005), as well as in youths with (Leibenluft et al. 2007) and at high risk for (Singh et al. 2008) BD when compared to age-comparable controls. In addition to being observed across the lifespan, deficits in this domain of executive function have also been observed across mood states in affected individuals (Blumberg et al. 2003a) and their unaffected relatives (Frangou et al. 2005; Giakoumaki et al. 2007), suggesting that impaired response inhibition may represent an important underlying trait of BD, particularly in the presence of co-occurring ADHD.
Like its behavioral manifestation, the neural basis of response inhibition may be task-dependent (Mostofsky et al. 2003; Wager et al. 2005; Simmonds et al. 2008) and developmentally dynamic (Bunge et al. 2002; Tamm et al. 2002; Rubia et al. 2006). However, many studies agree that successful response inhibition generally involves interactive engagement of the ventrolateral (VLPFC) and dorsolateral (DLPFC) prefrontal cortices to provide inhibitory control (Liddle et al. 2001), the striatum (Vink et al. 2005) to control the execution of planned motor responses, and the anterior cingulate cortex (ACC) to make and monitor response-related decisions (Braver et al. 2001; Liddle et al. 2001). Individuals with BD are at particular risk for unsuccessful response inhibition due to frontostriatal dysfunction implicated in the underlying pathophysiology of this condition (Blumberg et al. 2003a; Blumberg et al. 2003b; Chang et al. 2004; Elliott et al. 2004; Strakowski et al. 2004; Adler et al. 2004) and due to its common co-occurrence with ADHD (Adler et al. 2005).
Among functional magnetic resonance imaging (fMRI) studies, few have investigated the neural correlates of co-occurring BD and ADHD and have shown inconsistent findings (Adler et al. 2005; Leibenluft et al. 2007). In one study, BD adolescents with (n = 11) and without (n = 15) co-morbid ADHD were compared. The groups performed equally well on the single-digit continuous performance task administered during fMRI. However, relative to the group without ADHD, subjects with co-morbid BD and ADHD showed decreased activation in the VLPFC and ACC and increased activation in the posterior parietal and medial temporal gyrus during the task (Adler et al. 2005). The task in this study did not directly examine response inhibition, and the study was limited by a lack of a healthy control group. Another fMRI study using a stop-signal task paradigm compared motor inhibition in bipolar adolescents with and without ADHD and healthy adolescents without a personal or family history of any psychiatric disorders. The healthy adolescents showed greater bilateral striatal and right VLPFC activation than bipolar subjects during unsuccessful stop trials (Leibenluft et al. 2007). During this task, patients with BD and ADHD did not differ in any region of interest from BD subjects without ADHD. In addition, comparison subjects had greater activation than BD subjects with ADHD in the striatum, bilateral anterior cingulate, and bilateral ventral prefrontal cortex, but only showed greater activation compared to BD without ADHD in the bilateral accumbens and left caudate. Even with a comparison group of bipolar adolescents without ADHD, this study illustrated the challenges associated with trying to assess the independent contribution of ADHD to be discriminated from that contributed by BD with respect to between-group behavioral or brain activation differences. This study was limited by region-of-interest analyses, which may have reduced detection of multiple regions involved in response inhibition (Aron et al. 2007). Careful task selection, employment of a whole-brain analysis, and examination with larger numbers of subjects with comparable heterogeneity may clarify these inconsistent results and provide a more convergent model of frontostriatal dysfunction in adolescents with co-occurring BD and ADHD.
With these considerations in mind, we compared behavioral variables and corresponding whole brain activation between BD and community, age-comparable healthy adolescents (healthy controls, HC) while performing a Go/NoGo task. To optimize statistical power insufficient in event-related designs and to ensure adequate performance on executive function in a pediatric population with a clinically heterogeneous disorder, a traditional Go/NoGo task was employed with experimental blocks comprised of both Go and NoGo trials with half the trials in the experimental block containing NoGo conditions.
Thus, based on our prior work and the extant literature on response inhibition in BD, we hypothesized that, compared to HC, BD adolescents would have high rates of co-occurring ADHD and would exhibit lower accuracy and increased error rates while performing the Go/NoGo task. We also predicted that the BD group would manifest reduced activation in frontostriatal regions compared to controls, which, in turn, would correspond to performance during the response inhibition task. Finally, we hypothesized that more pronounced symptoms of ADHD, but not mania or depression, would be correlated with decreased neural activation in frontostriatal regions during response inhibition in our BD group.
Materials and Methods
Subjects
Children and adolescents between the ages 9 and 18 years who met Diagnostic and Statistical Manual of Mental Disorders, 4th edition text revision (DSM-IV-TR) (American Psychiatric Association 2000) diagnostic criteria for BD I or II (n = 26) were recruited by referral to academic adult and pediatric BD clinics and the surrounding community. Age- and gender-comparable healthy control children and adolescents (HC, n = 22) without a history of psychiatric diagnoses or the occurrence of such diagnoses in their first-degree relatives were recruited from the same community. Demographic and clinical characteristics were collected for all of subjects at the beginning of the study, including age, sex, intelligence quotient (IQ), and handedness. Handedness was assessed by the Crovitz Handedness Questionnaire (Crovitz and Zener 1962). BD subjects were excluded by the presence of a pervasive developmental disorder, seizure disorder, substance use disorder, IQ less than 80, and the presence of metallic implants or braces. A total of 81% of BD subjects were offspring of parents with BD I or II. Because a previous study has suggested that stimulants have a direct effect on the performance of a response inhibition task (Epstein et al. 2007b), all participants in the study were washed out from stimulants 24 hours prior to neuroimaging. To avoid risk of mood destabilization, BD subjects were allowed to continue any other current psychotropic medications, including mood stabilizers, atypical antipsychotics, or antidepressants. Subjects' medication history was obtained and used for exploratory and covariate analyses of neural findings.
All participants were evaluated using the Washington University in St. Louis Kiddie-Schedule for Affective Disorders and Schizophrenia (WASH-U K-SADS) (Geller et al. 1996), administered separately to parents and children, by raters blind to diagnostic group, with established symptom and diagnostic reliability (κ > 0.9). All diagnoses were determined by a consensus conference attended by a child and adolescent psychiatrist (K.D.C.) and the WASH-U K-SADS interviewer, after both parent and child interviews were completed. Mood symptom severity in BD subjects was assessed using the Young Mania Rating Scale (YMRS) (Young et al. 1978), and the Children's Depression Rating Scale–Revised Version (CDRS-R) (Poznanski et al. 1979) by raters with established symptom reliabilities (ICC > 0.9). An independent parent assessment of symptoms of inattention and hyperactivity (ADHD) was obtained using the 10-item abbreviated Conners' Rating Scale (Rowe and Rowe 1997; Tillman and Geller 2005). Parent psychiatric assessments in both groups were confirmed with the Structured Clinical Interview for DSM-IV (SCID-P) (First et al. 1996) by raters who were blind to diagnostic group and who had established diagnostic interrater reliability (κ > 0.9).
Behavioral assessment and analysis
Subjects were administered the Wechsler Abbreviated Scale of Intelligence (WASI) (Psychological Corporation 1999) and a computerized and standardized neuropsychological task which measures response inhibition. The experimental task consisted of two alternating conditions: Go and NoGo/Go (Fig. 1). Throughout both conditions, subjects viewed a series of letters once every 2 seconds (500 msec stimulus 1,500 msec interstimulus interval [ISI]) and responded with a key press, using the forefinger of the right hand, to every letter except the letter “X,” to which they were instructed to withhold response. In the Go (control) condition, subjects were presented a random sequence of letters other than the letter “X.” In the NoGo/Go (experimental) condition, subjects were presented with the letter “X” 50% of the time. The entire task lasted a total of 372 seconds and consisted of 12 alternating 26-second epochs of Go and NoGo/Go conditions flanked at the beginning and end by 30-second rest epochs during which the subject passively viewed a blank screen. Each epoch consisted of a 2-second instruction alerting the subject to the present task condition followed by 12 trials per epoch. Responses and reaction times were recorded.
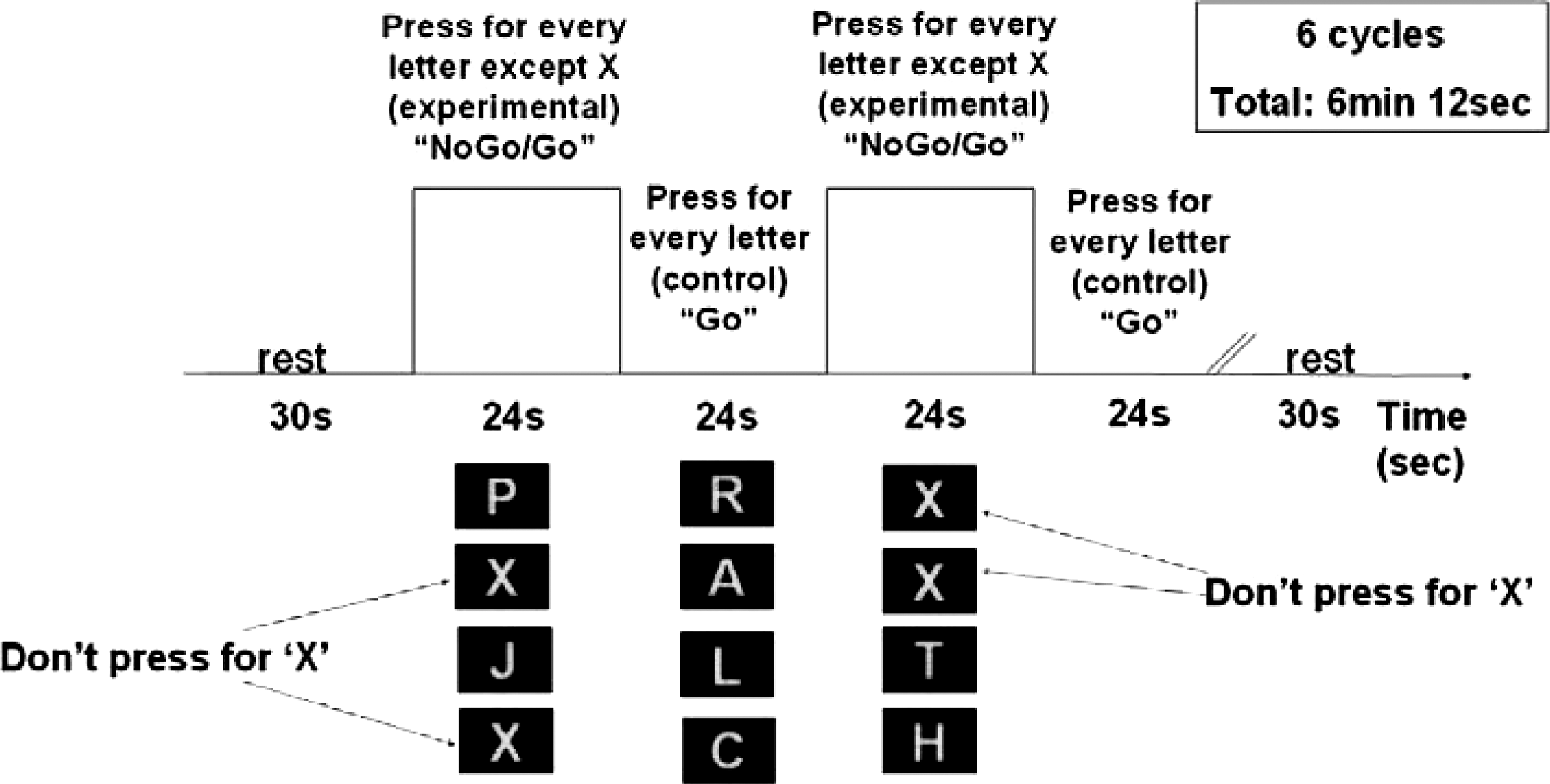
The Go/NoGo task sequence consisting of 12 alternating experimental and control epochs. In each epoch, subjects viewing a series of letters presented for 500 msec each, with a 1,500-msec interstimulus interval (ISI). In the experimental epoch, subjects were instructed to respond with a key press for every letter except “X.” In the control epoch, subjects were presented a random sequence of letters other than the letter “X” and instructed to respond for every letter.
For the in-scanner task, the proportion of trials to which subjects correctly responded in a Go trial (Go% correct) and correctly inhibited in a NoGo trial (NoGo% correct) was calculated. Participants were excluded from the fMRI analysis if their performance on the scanner task fell at or below chance on both Go% correct and NoGo% correct. reaction times (RTs) for correctly pressing the key in Go trials (GoRT) and mistakenly pressing the key in NoGo trials (NoGoRT) were also calculated. Accuracies to NoGo trials (false alarms) are the conventional means of indexing cognitive control in this paradigm. However, to take into account target hits as well as false alarms, d-prime was calculated by subtracting z-transformed proportions of false alarms from proportions of positively responded Go trials (hits). d-Prime provides a measure of sensitivity in the discrimination and ultimate detection of target stimuli relative to nontarget stimuli (Wickens 2002), a key component of cognitive control (Nigg and Casey 2005) (i.e., how well the subject can discriminate and appropriately respond to targets and nontargets). Each behavioral measure was subjected to t-test or Wilcoxon Rank Sum Test, for parametric and nonparametric data, respectively, to assess for significant differences in performance between the two groups.
Functional MRI procedures
Image Acquisition, Motion Correction, and Processing
Images were acquired with a 3T GE Signa scanner (General Electric, Milwaukee, WI) using a standard fMRI whole-head coil. Functional images were collected with a T2*-weighted spiral pulse sequence with parameters of repetition time (TR) = 2,000 msec, echo time (TE) = 30 msec, flip angle 80°, voxel size 3.12 × 3.12 mm, and slice thickness 3.5 mm with 1-mm spacing. An automated high-order shimming method was used before acquiring fMRI data to reduce field inhomogeneities. Structural images were collected to aid in localization of the functional data, using high-resolution, T1-weighted, spoiled gradient-recalled acquisition in the steady-state (GRASS) three-dimensional MRI sequences with the following parameters: TR = 35 msec, TE = 6 msec, flip angle = 45°, field of view = 24 cm, 124 slices in the coronal plane, and a 256 × 192 matrix. Functional images were processed with statistical parametric mapping software (SPM2), including realignment, transformation to Montreal Neurological Institute (MNI)-space, followed by smoothing with full width at half-maximum (FWHM) = 8 mm.
Custom motion-processing methods were applied to maximize the number of usable subjects in this dataset, given that a proportion of them had high motion during the scan (7/22 HC and 12/26 BD subjects moved more than 1 mm). Simply discarding high-motion subjects could result in insufficient remaining subjects to achieve a strong group result. Conversely, including subjects with large motion can influence group results by motion-induced errors from the large motion subjects. Thus, artifact repair was used to detect and suppress outliers caused by large sharp movements that, in turn, caused image reconstruction and spin history errors (Mazaika et al. 2007) (
Of the originally recruited 27 HC and 34 BD subjects, 13 were excluded due to poor scan quality, poor behavioral data, motion greater than 5 mm, or poor global quality scores, leaving 22 HC and 26 BD subjects available for group analysis. A combination of these methods recovered many large-motion fMRI data sets and indicated which data sets were not recoverable. In this way, we were able to maximize the number of usable subjects in our clinical group study to 79% (48/61) of all fMRI data sets acquired.
Data analysis
Chi-squared analysis, two sample t-tests, and effect sizes (Cohen d) were applied to demographic, clinical, and behavioral data. Statistical analysis was performed for individual and group data using the general linear model and the theory of Gaussian random fields as implemented in SPM2. Activation foci were superimposed on high-resolution, T1-weighted images, and their locations were interpreted using the Talairach atlas and known neuroanatomical landmarks. Individual contrast images were computed for conditions in which performance of Go blocks was subtracted from the NoGo blocks. These contrast images were analyzed with SPM2 using a general linear model to determine voxelwise t-statistics. One-sample t-tests were used to determine within-group activation and two-sample t-tests were used to determine between-group differences. Between-group analyses used a random effects model (Holmes and Friston 1998). Clusters of activation to display were defined as those surpassing a height threshold of p < 0.001 and an extent threshold of 40 voxels for all within- and between-group analyses (Friston et al. 1995), where the height threshold provides a statistical threshold for the number of activated regions within a given volume such as an individual voxel (i.e., is analogous to a t-test), and an extent threshold represents the minimum size of contiguous voxels which comprise a particular cluster (
Results
Demographic and clinical characteristics
There were no statistically significant group differences between BD (n = 26) and HC (n = 22) subjects in age (15.4 ± 2.7 vs. 14.3 ± 2.5 years), sex (female, 38% vs. 41%), handedness (right, 87% vs. 100%), ethnicity (Caucasian, 88% vs. 81%), or full-scale IQ scores (109 ± 12 vs. 111 ± 8). (The between-group activation differences presented below did not change significantly after covarying for these demographic variables.) Eighteen (69%) of the BD children met DSM-IV-TR criteria for BD I and 8 (31%) met criteria for BD II. Consistent with previous reports (Findling et al. 2001; Biederman et al. 2004; Singh et al. 2006), 21 (81%) BD children had ADHD, 16 (62%) BD subjects had oppositional defiant disorder, and 12 (46%) met criteria for anxiety disorders (Table 1). HC children were free of DSM-IV Axis I mood, psychotic, disruptive behavioral, or anxiety disorders.
Abbreviations: BD = Bipolar disorder; HC = healthy controls; IQ = Intelligence quotient; SD = standard deviation; DSM-IV, Diagnostic and Statistical Manual of Mental Disorders, 4th edition; YMRS = Young Mania Rating Scale; CDRS = Children's Depression Rating Scale; N/A = not applicable.
Behavioral results
The BD group had lower proportions of correct responses on Go (84% vs. 96%, T(46) = 3.35, p = 0.002) and overall Total (85% vs. 94%, T(46) = 4.12, p = 0.0002) trials as compared to the HC group (Table 2). This suggests significant deficits in sustained effort or vigilance during a low cognitive load component of the task. However, there were no statistically significant group differences in response inhibition as determined by percentage correct on NoGo trials (p = 0.11), reaction times needed for correctly responding in Go trials (Z = 0.86, p = 0.39), or reaction times for false alarms (Z = 0.82, p = 0.41). The BD group showed lower scores on d-prime than the HC group (3.98 [2.5] vs. 5.69 [3.2], T[46] = 2.07, p = 0.045).
T = 3.35, df = 46, p = 0.002, d = 1.27, Levene's test: F(1,46) = 5.25, p = 0.03.
T = 4.12, df = 46, p = 0.0002, d = 1.31, Levene's test: F(1,46) = 6.33, p = 0.02.
T = 2.07, df = 46, p = 0.045, d = 0.60.
Abbreviations: SD = Standard deviation; BD = bipolar disorder; HC =healthy control.
Functional MRI results
Table 3 and Fig. 2 summarize the whole-brain functional neuroimaging results contrasting the NoGo minus Go (NoGo−Go) conditions. There were no statistically significant differences between groups for the Go-NoGo, Go-Rest, NoGo-Rest contrasts. For the NoGo−Go contrast of interest, the BD group showed greater activation in the right DLPFC (BA 9) relative to the HC group. The HC group did not show any regions of statistically greater activation relative to the BD group for the NoGo−Go contrast.

Areas of greater activation during the Go/NoGo task in the NoGo−Go (NoGo−Go) contrast in subjects with bipolar disorder compared with controls. R = Right; ACC = anterior cingulated cortex; L = left; Inf = inferior; DLPFC = dorsolateral prefrontal cortex.
Abbreviations: BA, Brodmann area; BD, bipolar disorder; IFG, Inferior frontal gyrus; R, right; ACC, anterior cingulate cortex; HC, healthy controls; DLPFC, dorsolateral prefrontal cortex.
Within-group analyses showed that the HC group had greater activation in the inferior frontal gyrus (Brodmann area [BA] 9, posteriorly) and the ACC during the NoGo condition relative to the Go condition. Within the BD group, significant activation was seen in the inferior frontal lobe, right ACC, and in the left occipital lobe for the NoGo−Go contrast. For the BD group, YMRS and CDRS symptom scores did not correlate significantly with activation in any brain region for the NoGo−Go contrast. (Symptom scores were clustered around 0 for the HC group and therefore were not assessed.) Conners' raw scores were positively correlated with greater activation in the brainstem for the NoGo−Go contrast within the BD group after adjusting for age (T[26] = 4.03, p < 0.001, k = 102).
On an exploratory basis, we examined whether activations in BD subjects for the NoGo−Go contrast were related to prolonged (>6 months) exposure to psychotropic medication. Lithium-exposed BD subjects (n = 8) had significantly greater activation in the left anterior cerebellum (T[24] = 4.96, p < 0.001, cluster size, k = 90) than those not exposed to lithium, whereas those subjects not exposed to lithium showed relatively greater activation in the left ACC (BA 32, T[24] = 4.62, p < 0.001, k = 90). BD subjects exposed to atypical antipsychotics (n = 6) had significantly greater activation in the right ACC (BA 32, T[24] = 5.07, p < 0.001, k = 51) and right precuneus (BA 31, T[24] = 4.71, p < 0.001, k = 242) than BD subjects unexposed to antipsychotics. Atypical antipsychotic exposure was also associated with relatively reduced activation in the right medial frontal gyrus (T[24] = 4.31, p < 0.001, k = 44). There were no statistically significant differences in brain activation patterns in BD subjects exposed versus unexposed to valproic acid (n = 13), psychostimulants (n = 14), or antidepressants (n = 16) in the NoGo−Go contrast. None of the regions noted above overlapped with the spatial location of increased DLPFC activation in BD subjects relative to the HC group.
Discussion
Our results show that, relative to HC subjects, children and adolescents with BD demonstrate increased DLPFC activation in the presence of comparable performance during the Go/NoGo task. At the behavioral level, BD subjects had more inaccurate Go trials (lower Go% correct), which may be due to impairments in sustained attention (Aron et al. 2004), and lower d-prime scores, which may indicate difficulty discriminating and appropriately responding to targets and nontargets (Wickens 2002). Analysis of reaction times or errors in omission or commission did not indicate significant performance differences between the groups. This suggests that subjects from both groups were able to perform the inhibitory control task adequately at the presented level of difficulty.
Taken together, these results are consistent with the view that excessive recruitment of the right prefrontal cortex by the BD group during NoGo blocks may represent a mechanism for improving behavioral performance during response inhibition. Furthermore, our results support previous behavioral studies, suggesting that problems in sustained attention may represent a trait deficit of BD (Clark and Goodwin 2004; Doyle et al. 2005; Fleck et al. 2005). Despite the discrepancy in Go trial behavioral performance, there were no significant between-group activation differences for the Go versus Rest contrast. This finding may indicate that, unlike the increased DLPFC activation observed for NoGo blocks, BD subjects were not able to neurally compensate for intrinsic deficits in sustained attention. Selective increase in DLPFC activation during NoGo blocks in the BD group may have resulted from the subjects' deliberate attempt to comply with the experimental focus on inhibiting inappropriate responses, as opposed to regulating more automatized behavior such as that required for Go blocks.
Our results are partially consistent with previous fMRI investigations that have reported abnormalities in prefrontal circuitry in pediatric BD (Blumberg et al. 2003b; Chang et al. 2004; Leibenluft et al. 2007). While performing a color-naming Stroop task, 10 adolescents with BD, relative to controls, demonstrated increased activation in subcortical regions, including the putamen and thalamus, relative to prefrontal areas, suggesting a developmental disturbance in prefrontal functioning (Blumberg et al. 2003b). In a study of twelve 9- to 18-year-old males with BD, BD subjects showed greater left DLPFC activation compared to controls during a visuospatial working memory task, and greater bilateral DLPFC activation while viewing negative relative to neutral-valenced pictures (Chang et al. 2004). Some (n = 10) among this cohort also performed the Go/NoGo task, and their results are presented in our study. Taken together, these studies suggest prefrontal dysfunction in pediatric BD across a variety of tasks, but are in contrast to the recent finding of increased frontostriatal activation in healthy controls relative to BD subjects during a stop signal task (Leibenluft et al. 2007).
In contrast to prior studies, our findings suggest an atypical pattern of dorsal prefrontal activation in BD while achieving behavioral performance comparable to healthy controls. Alternatively, increased DLPFC activation in the BD group may be generally associated with greater neural effort needed to perform the cognitive task (MacDonald et al. 2000; Compton et al. 2003) compared to the HC group. Differences in the tasks, behavioral conditions, statistical contrasts employed, sample sizes, demographics, and clinical status across studies may all contribute to the variable results. For example, the lack of findings in the striatum in our current study may be attributable to the use of a simple design that may not have featured complex cognitive processes mediated by the striatum. Nevertheless, consistent with previous findings alluding to abnormalities in prefrontal circuitry in BD, the prefrontal activation profile in the presence study may be associated with emotional and attentional dysregulation in BD youth.
Some studies in adults with BD have suggested mood state-dependent changes in subcortical neural activation during simple motor tasks (Caligiuri et al. 2003; Caligiuri et al. 2006; Lohr and Caligiuri 2006). On the basis of mood symptom ratings, our BD group, on average, had mild severity of manic symptoms (mean YMRS score = 15.6) and symptoms commensurate with a depressed state (mean CDRS score = 44.1). However, neither mania nor depression symptom severity was correlated with any task-related behavioral measures or brain activation within the BD group, suggesting that prefrontal overactivation observed in the BD group during this task may be independent of a current mood state. This lack of correlation between mood state and fMRI activation is supported by other studies performed in BD subjects during periods of euthymia (Strakowski et al. 2004), and longitudinal studies in which frontostriatal activations remain consistent across several mood states (Marchand et al. 2007).
Alhough we did not observe any indication that mood state affected our primary results, previous studies from our group and others suggest that increased prefrontal activation in subjects with mood dysregulation may be a sign of increased cortical control over a hyperactive limbic system (Mayberg 1997; Chang et al. 2004; Phillips and Vieta 2007; Pavuluri et al. 2008). There may be several possible reasons why we did not observe limbic hyperactivation in this study. First, our task was not designed to target amygdalar activation specifically. Nevertheless, it is possible that prefrontal structures may have successfully suppressed any task independent limbic hyperactivation in the BD group, even in the presence of mild to moderate hypomanic and depressive symptomatology. Alternatively, functional differences in limbic structures may only be present during active (Foland et al. 2008) or prolonged symptoms of mania. In support of the latter hypothesis, bipolar adults who have a longer course of manic symptoms show patterns of limbic hyperactivity (Wessa et al. 2007) and diminished prefrontal activity (Malhi et al. 2007) while performing both cognitively and emotionally relevant fMRI tasks.
The co-occurrence of ADHD may be important when attempting to interpret behavioral and neural correlates of response inhibition in BD (Henin et al. 2007). In our study, most (81%) BD subjects met criteria for ADHD and had high overall raw and standardized Conners' scores. Given the preponderance of ADHD symptoms in our sample, it is surprising that our results did not show reductions in frontostriatal activation during tasks of cognitive control as demonstrated in populations with ADHD only (Durston et al. 2003; Rubia et al. 2005; Vaidya et al. 2005). Other studies have also shown increased activation in the ventrolateral prefrontal cortex during interference control and response competition in ADHD (Schulz et al. 2005).
To address this issue directly, we would need to have had more equal proportions of BD subgroups with and without ADHD and have an additional comparison group comprised of children with just ADHD of comparable severity to our BD group. Although there were no significant correlations between ADHD symptom severity as measured by Conners' scores and task-related behavioral measures, brainstem activation in the level of the pons was positively correlated with higher (more severe) Conners' scores after adjusting for age, suggesting activation of regions other than frontostriatal networks previously examined in similar co-morbid populations (Adler et al. 2005; Leibenluft et al. 2007). Increased pontine activation could facilitate a compensatory level of alertness in the BD group to permit more accurate or control level behavioral performance on Go and NoGo epochs. Feasibility of brainstem fMRI activation has been demonstrated (Komisaruk et al. 2002; Campbell et al. 2007), but further investigation of this finding in pediatric populations is warranted. Moreover, more precise measures of ADHD symptom severity than the parent-reported Conners' rating scale may be needed to confirm the relationship between brain activation and symptom severity.
The NoGo−Go contrast employed in this study is associated with several cognitive processes and component functions of response inhibition, including sustained attention, target detection, and rule maintenance (Aron and Poldrack 2005). We chose a block design to examine a combination of these processes over a sustained period of time to capitalize on a higher proportion of NoGo trials represented as compared to those generated by an event-related analysis. Although event-related designs and analyses might be able to extract individual cognitive processes subserved by activated brain regions (e.g., differentiating neural processes occurring during failed versus successful NoGo trials), they generate a lower proportion of NoGo trials than a block design and are extremely difficult for children with serious mood disorders to perform.
Moreover, in the context of high individual variances due to complex clinical heterogeneity from psychiatric disorders co-occurring with BD such as ADHD, a block design approach was preferred over an event-related design. This approach has been successfully used in typically developing children (Tamm et al. 2002) and those with posttraumatic stress symptoms (Carrion et al. 2008), fragile X (Hoeft et al. 2007), and ADHD (Epstein et al. 2007b). Using this method, the neural result of dorsolateral prefrontal overactivation in BD compared with HC is of particular importance because high cortical cognitive areas may be vulnerable to neurodegeneration and dysfunction with progression of bipolar illness (Chang 2007).
Limitations
There are a few limitations to this study, including prior and current history of medication exposure in the BD group and a lack of an ADHD-only comparison group. The generalizability of our results and our ability to detect some functional group differences may have been limited by interindividual differences in mood states and cognitive strategies employed to optimize task performance. Exposure of our subjects to psychotropic medications represents another potential confound. However, due to ethical and practical considerations, it is difficult to perform MRI studies on unmedicated children with BD. Exploratory analyses suggested that there may be general effects of medication exposure on brain activation in BD subjects, although the group sizes for these analyses were quite small and the length of exposure and number of medications variable. This suggests the possibility of type I and II errors, as has been described in previous studies examining medication effects on fMRI results (Leibenluft et al. 2007).
Thus, the results of the medication analyses should be interpreted with caution and viewed as preliminary and hypothesis generating. For multiple reasons, including the co-occurrence of ADHD symptoms (Epstein et al. 2007a), the BD group had high potential for motion, which necessitated the use of a mock scanner training protocol prior to the scan and applying a motion correction algorithm during postprocessing procedures. Behavioral or functional subgroup analyses on BD subjects based on motion severity were not possible due to the small sample size, but task-correlated motion was not significantly different across the BD and HC groups. Future studies examining larger samples would permit additional subgroup analyses relating bipolar symptoms to cognitive performance on executive functioning tasks.
Conclusions
In summary, the results of this study indicate that children and adolescents with BD may need to recruit supplementary prefrontal resources to successfully perform a response-inhibition task at healthy control levels. These findings contribute to our current understanding of the neurofunctional phenotype in pediatric BD and suggest future directions for elucidating a neurobiological explanation for the co-occurrence of pediatric BD with ADHD. Additional assessment of neuropsychological performance coincident with functional neuroimaging investigations of children, adolescents, and adults with co-occurring BD and ADHD are necessary to further clarify the role of brain networks underlying response inhibition in these populations.
Footnotes
Disclosures
Drs. Singh, Mazaika, Garrett, Adleman, and Reiss, Mr. Kelley and Ms. Howe have no financial ties or conflicts of interest to disclose. Dr. Chang has received research grants from AstraZeneca Pharmaceuticals, Eli Lilly and Company, Otsuka America Pharmaceutical, Inc., and GlaxoSmithKline; is on the speakers bureau of Abbott Laboratories, AstraZeneca Pharmaceuticals, Bristol-Myers Squibb, and Eli Lilly and Company; is a consultant for Abbott Laboratories, AstraZeneca Pharmaceuticals, GlaxoSmithKline, Eli Lilly and Company, and Shire Pharmaceuticals; and is on the advisory boards of Abbott Laboratories and Eli Lilly and Company.
Acknowledgments
The authors gratefully acknowledge the support of the Helena Anna Henzl-Gabor Young Women in Science Fund, the National Institute of Mental Health (K23 MH064460), the National Alliance for Research on Schizophrenia and Depression (NARSAD), the Hahn Family, and the Klingenstein Third Generation Foundation.
Portions of this manuscript were presented at the 53rd Annual Meeting of the American College of Neuropsychopharmacology, Boca Raton, FL, December 9–12, 2007.