
Abstract
Objective:
Genome-wide association studies have identified allelic variation in CACNA1C as a risk factor for multiple psychiatric disorders associated with limbic system dysfunction, including bipolar disorder, schizophrenia, and depression. The CACNA1C gene codes for a subunit of L-type voltage-gated calcium channels, which modulate amygdala function. Although CACNA1C genotype appears to be associated with amygdala morphology and function in adults with and without psychopathology, whether genetic variation influences amygdala structure and function earlier in development has not been examined.
Methods:
In this first investigation of the neural correlates of CACNA1C in young individuals, we examined associations between two single nucleotide polymorphisms in CACNA1C (rs1006737 and rs4765914) with amygdala volume and activation during an emotional processing task in 58 adolescents and young adults 13–20 years of age.
Results:
Minor (T) allele carriers of rs4765914 exhibited smaller amygdala volume than major (C) allele homozygotes (β=−0.33, p=0.006). Furthermore, minor (A) allele homozygotes of rs1006737 exhibited increased blood–oxygen-level-dependent (BOLD) signal in the amygdala when viewing negative (vs. neutral) stimuli (β=0.29, p=0.040) and decreased BOLD signal in the amygdala when instructed to downregulate their emotional response to negative stimuli (β=−0.38, p=0.009). Follow-up analyses indicated that childhood trauma did not moderate the associations of CACNA1C variation with amygdala structure and function (ps>0.170).
Conclusions:
Findings indicate that CACNA1C-related differences in amygdala structure and function are present by adolescence. However, population stratification is a concern, given the racial/ethnic heterogeneity of our sample, and our findings do not have direct clinical implications currently. Nevertheless, these results suggest that developmentally informed research can begin to shed light on the time course by which genetic liability may translate into neural differences associated with vulnerability to psychopathology.
Introduction
E
In addition to research linking CACNA1C to various psychiatric disorders, studies have explored how CACNA1C variants relate to the morphology and function of limbic system structures, particularly the amygdala. Most studies have investigated rs1006737, the single nucleotide polymorphism (SNP) with the strongest association with bipolar disorder in a GWAS (Ferreira et al. 2008). The minor (A) allele of rs1006737 has been associated with increased right amygdala volume (Perrier et al. 2011; Wolf et al. 2014) and increased amygdala activation during emotional processing, with AA homozygotes exhibiting the most pronounced associations with amygdala structure and function (Bigos et al. 2010; Wessa et al. 2010; Jogia et al. 2011). CACNA1C variants have been associated with neural differences in adults with (Wolf et al. 2014) and without psychopathology (Wessa et al. 2010; Jogia et al. 2011). Most neuroimaging genetics research with CACNA1C has demonstrated associations in individuals without psychopathology, suggesting that CACNA1C-related differences may be preexisting, and potentially risk-conferring, variations. Although medium to large effect sizes have been detected in some studies of the neural correlates of rs1006737 (e.g., Wessa et al. 2010; Jogia et al. 2011; Perrier et al. 2011), overall, effect sizes observed for single SNPs on intermediate phenotypes such as neural structure and function have been modest in the broader neuroimaging genetics literature (e.g., Stein et al. 2012). Therefore, even though neural phenotypes have been proposed to lie closer to the underlying genetic architecture than more distal clinical outcomes such as psychiatric disorders (e.g., Meyer-Lindenberg and Weinberger 2006), it is important to note that these intermediate phenotypes are, nonetheless, complex and polygenic in nature.
To date, research on CACNA1C and amygdala structure and function has been limited to studies in adults. However, as LTCCs are involved in neural function early in development (Kempton et al. 2009), whether similar patterns of associations between CACNA1C and amygdala structure and function are present in youth represents an important empirical question. Developmentally informed investigations that can begin to elucidate the time course by which genetic liability may translate into neural differences associated with vulnerability to psychopathology are needed. In this study, we examined associations between CACNA1C variation with amygdala structure and function during emotional processing in adolescents. We investigated two SNPs in CACNA1C: rs1006737 and rs4765914 (associated with bipolar disorder, schizophrenia, and depression in a cross-disorder GWAS) (Cross-Disorder Group of the Psychiatric Genomics Consortium 2013). We selected the amygdala as our region of interest given the role of LTCCs in amygdala function (Shinnick-Gallagher et al. 2003) and robust evidence of associations between CACNA1C variation with amygdala structure and function in adults (Bhat et al. 2012). Because previous work suggests that genetic influences on amygdala structure and function vary as a function of childhood trauma (e.g., White et al. 2012), we also examined whether childhood trauma moderated associations between CACNA1C genotype and amygdala volume and activation.
Materials and Methods
Participants and procedure
A community-based sample of 59 adolescents and young adults 13–20 years of age was recruited in Boston and Cambridge, Massachusetts to complete a functional MRI (fMRI) study as part of a larger cohort study of adolescents (see McLaughlin et al. 2014, for details). One individual was missing valid CACNA1C genetic data; therefore, 58 individuals comprised the analytic sample. The analytic sample was 60.3% female (n=35), with a mean age of 16.8 years (SD= 1.5). All females were postmenarchal. The sample was diverse with respect to race and ethnicity: 27.6% white (n=16), 31.0% black (n=18), 20.7% Hispanic (n=12), 6.9% Asian (n=4), and 13.8% biracial or other (n=8). Recruitment efforts were targeted to obtain a sample with high variability in exposure to childhood trauma; 44.8% of participants (n=26) had a history of childhood trauma (as defined subsequently).
Adolescents attended the study sessions with a parent or guardian; informed consent was obtained from the parent/guardian, and assent was provided by adolescents (participants≥18 years of age provided informed consent). Participants completed a neuroimaging session that consisted of a structural scan and fMRI task. During a separate study session, participants provided saliva samples for DNA collection and completed measures of childhood trauma. Study protocol and procedures were approved by the Institutional Review Board of Boston Children's Hospital and Harvard University.
fMRI task
Participants engaged in a widely used event-related task designed to assess neural markers of emotional reactivity and emotion regulation (Ochsner et al. 2004) that has been used in dozens of investigations (Buhle et al. 2014), including with children and adolescents (McRae et al. 2012). Task design and construction of contrasts for analysis were based on the substantial literature on this task. Specifically, participants viewed neutral and negative images drawn from the International Affective Picture System (IAPS) (Lang et al. 2008). Prior to each image, participants were shown an instructional cue to either “look” or “decrease” (see Fig. 1). During look trials, participants were instructed to allow their emotions to unfold naturally and not to engage in active strategies to modify their emotional response. During decrease trials, participants engaged in specific cognitive reappraisal strategies to try to reduce their emotional response. Participants were trained in reappraisal strategies during a training session prior to the scan. After being presented with the neutral or negative image, participants provided a rating of their negative affect on a 0 (low) to 4 (strong) scale. In the current study, we assessed emotional reactivity to negative stimuli by comparing neural activation during passive viewing of negative versus neutral stimuli. We assessed emotion regulation to negative stimuli by comparing neural activation when subjects were instructed to downregulate their emotional response to negative stimuli versus when they were passively viewing negative stimuli.
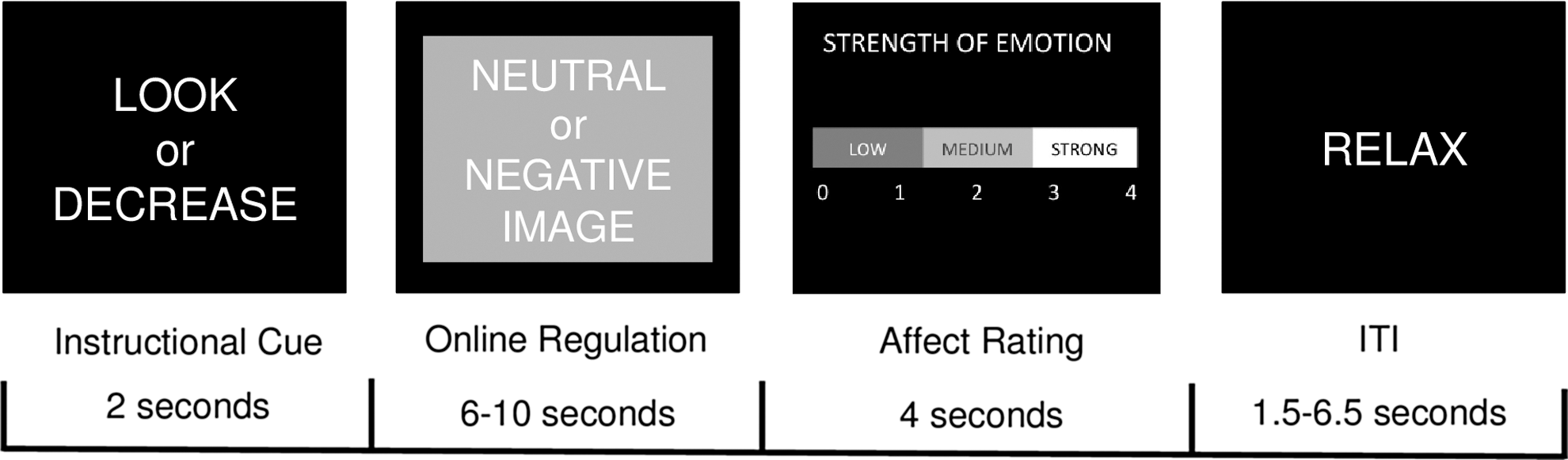
Timeline for events on each trial of the functional MRI (fMRI) task. An instructional cue directed participants to look at the following stimulus or decrease their emotional response to it; participants then followed this instruction when presented with a neutral or negative image. Participants provided a rating of their current negative affect, and then had a period to relax before the start of the next trial. ITI, intertrial interval.
Stimuli were presented in four runs lasting 9 minutes each. The instructional cue appeared for 2 seconds, the emotional stimulus appeared for 6–10 seconds, the affect rating screen appeared for 4 seconds, and the inter-trial interval (ITI) lasted from 1 to 6.5 seconds. The emotional stimulus and ITI were jittered by sampling durations in the following manner: 50% of the shortest duration, 25% of the middle duration, and 25% of the longest duration, following existing recommendations (Ollinger et al. 2001).
Image acquisition
Scanning was performed on a 3T Siemens Trio scanner at the Harvard Center for Brain Science using a 32 channel head coil. Anatomical scans (T1-weighted multi-echo magnetization-prepared rapid acquisition with gradient echo [MPRAGE] volumes) were acquired for co-registration with fMRI (TR=2530 ms, TE=1640–7040 ms, flip angle=7 degrees, field of view [FOV]=220 mm2, 176 slices, in-plane voxel size=1 mm3). To reduce motion-related artifacts, a navigator echo was used prior to scan acquisition, which compared slices to this echo online and permitted up to 20% of slices to be reacquired.
Blood–oxygen-level-dependent (BOLD) signal during functional runs was acquired using a gradient-echo T2*-weighted echo planar imaging (EPI) sequence. Thirty-two 3 mm thick slices were acquired parallel to the anterior commissure (AC)–posterior commissure (PC) line (TR=2000 ms, TE=30 ms, flip angle=90 degrees, bandwidth=2300, echo spacing=0.5, FOV=256×256, matrix size=64×64). Prior to each scan, four images were acquired and discarded to allow longitudinal magnetization to reach equilibrium. An online prospective motion correction algorithm (PACE) was used to reduce the effect of motion artifacts.
Image processing
T1-weighted scans were processed using FreeSurfer version 5.0 (Dale et al. 1999; Fischl et al. 1999; Fischl and Dale 2000; Fischl et al. 2002,\ 2004). Automatic image segmentation was used to identify subcortical gray matter structures. Gray/white matter and gray matter/cerebrospinal fluid (CSF) boundaries were constructed using spatial intensity gradients across tissue classes. Following reconstruction, the cortex was parcellated based on the structure of gyri and sulci (Fischl et al. 2004; Desikan et al. 2006). Results were inspected and manually edited to optimize accurate placement of gray/white and gray/CSF borders based on shifts in the image intensity gradient (Dale et al. 1999; Fischl and Dale 2000). FreeSurfer morphometric procedures have demonstrated good test–retest reliability across scanner manufacturers and field strengths (Han et al. 2006; Reuter et al. 2012), and have been validated against manual measurement (Kuperberg et al. 2003; Salat et al. 2004) and histological analysis (Rosas et al. 2002). These procedures have been used with youth (Østby et al. 2009; Tottenham et al. 2010; Sheridan et al. 2012).
Preprocessing and statistical analysis of fMRI data was performed in Nipype (
Genotyping
Participants provided saliva samples for DNA collection using Oragene® kits (DNA Genotek, Ontario, Canada). We examined two SNPs in CACNA1C: rs1006737 (Bhat et al. 2012) and rs4765914 (Psychiatric GWAS Consortium Bipolar Disorder Working Group 2011; Cross-Disorder Group of the Psychiatric Genomics Consortium 2013). Both SNPs are located in intron 3 of CACNA1C (Bhat et al. 2012), although they did not exhibit substantial linkage disequilibrium (r2 =0.28).
Genotyping was conducted at the Massachusetts General Hospital (MGH) Psychiatric and Neurodevelopmental Genetics Unit Core Lab as part of a larger study from which participants were drawn (McLaughlin et al. 2014). Genotyping of rs4765914 was conducted as part of this analysis for 46 participants in the current study who were drawn from this larger sample. DNA extraction and SNP genotyping were performed using the Sequenom iPLEX Gold® application and MassARRAY® system at MGH. A total of 40 ancestry-informative markers 1 were also genotyped for these individuals. These 40 SNPs were analyzed using principal components analysis to infer underlying population structure. As described subsequently, we included the first two principal components in a sensitivity analysis to address concerns regarding population stratification (i.e., the presence of systematic differences in allele frequencies as a function of subpopulations in the sample; e.g., Pritchard and Rosenberg 1999).
An additional 10 participants were recruited into the neuroimaging sample after this genotyping was completed. Genotyping of rs4765914 for these 10 participants, along with genotyping of rs1006737 (the most commonly studied SNP in CACNA1C) for all 58 participants in the current sample, was completed at the Behavioral and Neurodevelopmental Genetics Laboratory at Tulane University School of Medicine. DNA extraction and SNP genotyping at Tulane were performed using the Applied Biosystems' TaqMan® SNP assays (rs1006737 assay number c_2584015 and rs4765914 assay number c_27892152).
At both sites, quality control to determine sample and genotyping quality was performed. For samples run at MGH, PLINK, a whole genome association analysis tool set, was utilized. Call rate was 95.8% for the MGH assays for rs4765914. Reproducibility of genotyping at MGH was assessed by genotyping two duplicate pairs; concordance was excellent (r2 =1.00). For samples genotyped at Tulane, all samples were run in duplicate with known controls. All duplicates successfully amplified, and no samples had to be repeated. There were no duplicate assays across sites. Given some evidence that neural structural and functional differences are most pronounced for rs1006737 minor (A) allele homozygotes (Kempton et al. 2009; Bigos et al. 2010), we coded rs1006737 genotype as A allele homozygotes versus major (G) allele carriers. Because of the low number of rs4765914 minor (T) allele homozygotes (n=2), we examined T allele carriers versus major (C) allele homozygotes.
Childhood trauma
Childhood trauma was assessed using a self-report questionnaire and an interview. We administered the Childhood Trauma Questionnaire (CTQ) (Bernstein et al. 1997), a well-validated 28 item measure of abuse experienced during childhood and adolescence, with separate subscales for physical, sexual, and emotional abuse (Bernstein et al. 1994, 1997). These subscales had good internal consistency (α=0.75 for physical abuse, α=0.94 for sexual abuse, and α=0.86 for emotional abuse). The Childhood Experiences of Care and Abuse (CECA) interview (Bifulco et al. 1994; 1997) was also administered to assess exposure to physical and sexual abuse during childhood and adolescence. Interrater reliability for maltreatment reports is excellent, and multiple validation studies suggest high agreement between siblings on reports of caregiver behaviors and maltreatment (Bifulco et al. 1994, 1997). Participants who reported physical or sexual abuse during the CECA interview or who had a score on any of the three CTQ abuse subscales (physical, sexual, and emotional abuse) above a previously identified threshold (Walker et al. 1999) were classified as having experienced childhood trauma. No participant was currently experiencing maltreatment, and the proper authorities were contacted in cases that raised safety concerns.
Statistical analysis
To identify task-related activation, regressors were created for each phase of the task: Instructional cue, stimulus, and affect rating periods separately for neutral and negative stimuli for look and decrease trials. Using FSL, a general linear model (GLM) was constructed to estimate the association between variation in BOLD signal and task demands across time for each subject, prior to normalization. Next, we extracted amygdala activation for each participant using a region of interest (ROI) approach. We constructed a structural ROI of the amygdala in each participant's native space using FreeSurfer, and extracted the average estimate of neural activation within the entire amygdala ROI for each participant for emotional reactivity to negative stimuli (look negative>look neutral trials) and emotion regulation to negative stimuli (decrease>look negative trials) contrasts.
We examined CACNA1C variants as predictors of amygdala volume and task-related activation. For the morphology analyses, linear regression was used to predict bilateral amygdala volume (summed across the right and left hemispheres). For analyses of neural function, linear regression was used to predict the average BOLD signal extracted for the emotional reactivity and emotion regulation contrasts of interest. Analyses included age, gender, and white/nonwhite racial/ethnic status as covariates; in the amygdala morphology analyses, we also covaried total intracranial volume. Analyses were conducted separately for the two CACNA1C SNPs; two tailed tests were used. We further explored whether exposure to childhood trauma moderated associations between CACNA1C variation and amygdala volume and function.
Results
Participant characteristics
Descriptive statistics are presented in Table 1. Genotype frequencies for rs1006737 and rs4765914 did not deviate significantly from Hardy–Weinberg equilibrium, ps>0.05. There were no significant differences in gender, white/nonwhite racial/ethnic status, history of childhood trauma, or age as a function of rs1006737 A allele homozygote or rs4765914 T allele carrier status, ps>0.05 (see Table 1). There were trends for the percentages of female and white participants to be higher among rs1006737 G allele carriers compared with A allele homozygotes, although these differences did not reach statistical significance (ps>0.066). Given the racial/ethnic heterogeneity within our sample, we examined whether there were differences in amygdala structure and activation during emotional processing as a function of white/nonwhite racial/ethnic status. No significant differences emerged between white and nonwhite participants in amygdala volume, emotional reactivity, or emotion regulation, ps>0.116. However, nonwhite participants had smaller total intracranial volume (mean=1498809.52, SD=163187.79) than white participants (mean=1629375.00, SD=210030.75, t[56]=-2.51, p=0.015). As noted, white/nonwhite racial/ethnic status, sex, and age were included as covariates in all analyses.
Genotype frequencies did not deviate significantly from Hardy–Weinberg equilibrium, ps>0.05.
Two individuals were missing genotype data for rs4765914.
CACNA1C variation and amygdala structure
Results from the linear regressions predicting amygdala volume are presented in Table 2. rs4765914 genotype was significantly associated with amygdala volume (β=−0.33, p=0.006), such that T allele carriers exhibited smaller amygdala volume (mean=3242.44, SD=558.62) compared with C allele homozygotes (mean=3384.00, SD=420.07; see Fig. 2). 2 rs4765914 genotype accounted for 9% of the variance in amygdala volume over and above the other variables in the model. rs4765914 T allele carrier status was a stronger predictor of right (b=−188.18, SE=64.80, β=−0.33, p=0.005) than of left amygdala volume (b=−134.72, SE=72.22, β=−0.25, p=0.068), although associations were in the same direction for both hemispheres, and the genotype–hemisphere interaction was not statistically significant (p=0.491). History of childhood trauma did not moderate the association between rs4765914 genotype and amygdala volume (F[1, 48]=0.004, b=13.44, SE=223.74, β=0.01, p=0.952). 3 rs1006737 genotype was not significantly associated with amygdala volume, alone (see Table 2) or in interaction with childhood trauma (F[1, 50]=0.06, b=65.98, SE=263.40, β=0.04, p=0.803).
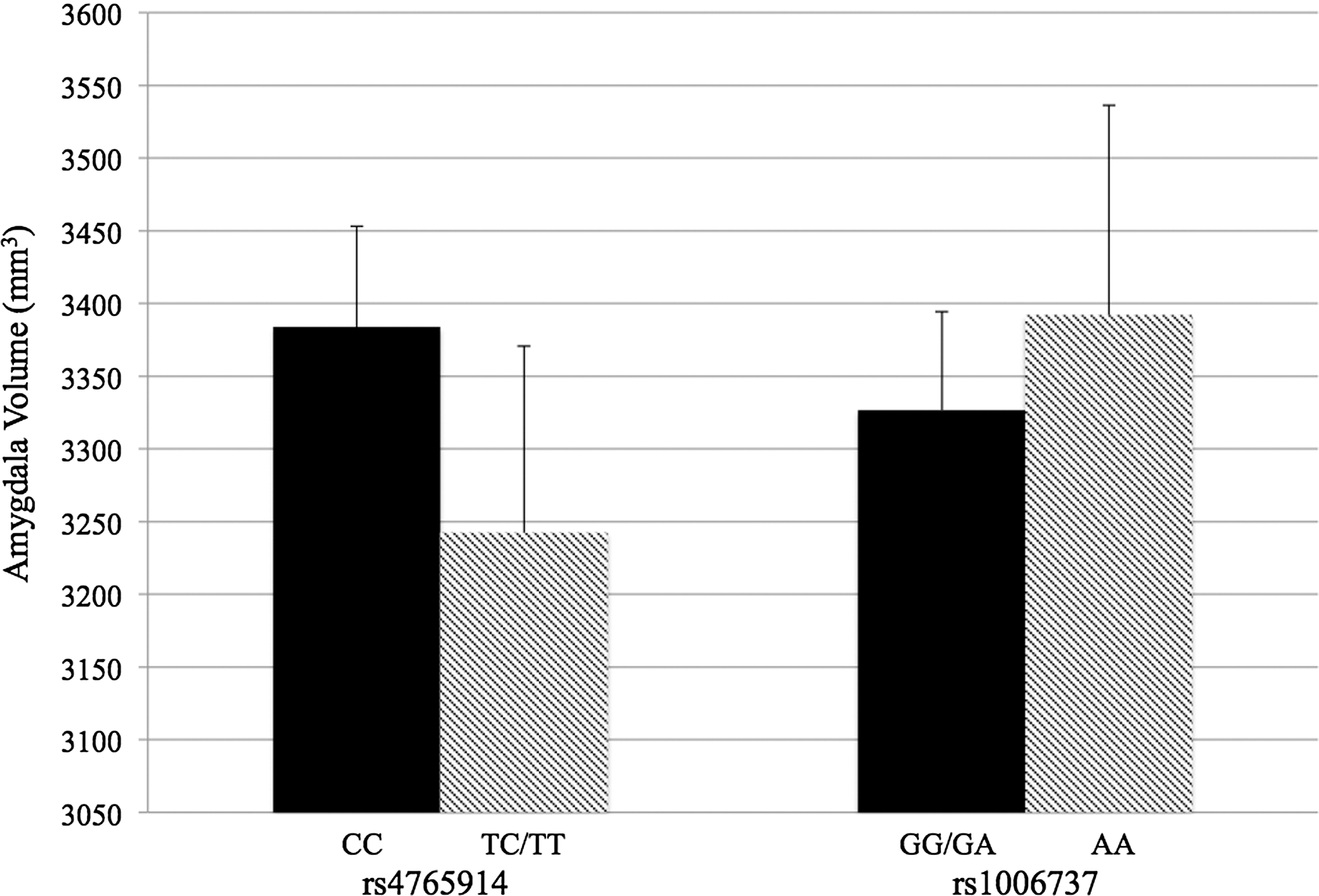
Mean amygdala volume as a function of CACNA1C genotype. Error bars represent standard error of the mean.
rs1006737 genotype coded as A allele homozygote status.
rs4765914 genotype coded as T allele carrier status.
CACNA1C variation and amygdala activation during emotional processing
As shown in Table 3, rs1006737 genotype was significantly associated with BOLD signal in the amygdala during the emotional processing task. During emotional reactivity trials, A allele homozygotes exhibited increased amygdala activation to negative versus neutral stimuli compared with G allele carriers (β=0.30, p=0.032; see Fig. 3a). 4 rs1006737 genotype accounted for 7% of the variance in BOLD signal in the amygdala during the emotional reactivity trials over and above the other variables in the model. The associations between rs1006737 genotype and amygdala activation were similar in magnitude for the right (b=6.27, SE=3.35, β=0.26, p=0.066) and left hemispheres (b=6.46, SE=3.28, β=0.28 p=0.054), and there was no statistically significant genotype–hemisphere interaction (p=0.945). History of childhood trauma did not significantly interact with rs1006737 genotype to predict amygdala activation during emotional reactivity trials (F[1, 51]=1.93, b=−7.95, SE=5.73, β=−0.25, p=0.171). rs4765914 genotype was not significantly related to BOLD signal in the amygdala during emotional reactivity trials, alone (see Table 3) or in interaction with childhood trauma (F[1, 49]=0.67, b=−4.44, SE=5.42, β=−0.19, p=0.417).

Mean amygdala activation to
rs1006737 genotype coded as A allele homozygote status.
rs4765914 genotype coded as T allele carrier status.
rs1006737 genotype also significantly predicted amygdala activation during emotion regulation trials (Table 3). Compared with G allele carriers, A allele homozygotes exhibited lower BOLD signal in the amygdala during trials in which participants aimed to decrease their emotional response to negative stimuli compared with passive viewing of negative stimuli (β=−0.38, p=0.009; see Fig. 3b). 5 Furthermore, rs1006737 genotype accounted for 12% of the variance in BOLD signal in the amygdala during emotion regulation trials after adjusting for the covariates in the model. The associations between rs1006737 genotype and amygdala activation were similar in magnitude for the right (b=−6.55, SE=2.60, β=−0.35, p=0.015) and left hemispheres (b=−6.75, SE=2.91, β=−0.33 p=0.024), and there was no statistically significant genotype–hemisphere interaction (p=0.754). Similar to the emotional reactivity trials, history of childhood trauma did not moderate the association between rs1006737 genotype and BOLD signal in the amygdala during emotion regulation trials (F[1, 51]=2.21, b=6.87, SE=4.62, β=0.26, p=0.143. There was no significant main effect of rs4765914 genotype on amygdala activation during emotion regulation trials (see Table 3) or significant interaction with childhood trauma (F[1, 49]=1.22, b=5.09, SE=4.61, β=0.26, p=0.275).
Addressing population stratification
As noted, we covaried white/nonwhite racial/ethnic status in all analyses given the racial/ethnic diversity of our sample. To further address concerns regarding population stratification (i.e., the presence of systematic differences in allele frequencies as a function of subpopulations in the sample; e.g., Pritchard and Rosenberg 1999), we inferred the underlying population structure from 40 ancestry-informative markers in a subset of participants (n=46). We reran analyses covarying the first two principal components from a principal components analysis of the ancestry-informative markers, along with the other covariates included in the models described. Our overall findings remained highly similar in effect size despite the smaller sample: rs4765914 genotype was still associated with amygdala volume (b=−405.82, SE=145.45, β=−0.39, p=0.008), and rs1006737 genotype was still associated with amygdala activation during the emotional reactivity (b=6.71, SE=3.63, β=0.30, p=0.072) and emotion regulation (b=−7.79, SE=3.04, β=−0.43, p=0.014) trials. These findings suggested that the associations between CACNA1C genotype and amygdala structure and function were not solely the result of confounding effects of population structure.
Discussion
To our knowledge, this is the first demonstration that genetic variation in CACNA1C is associated with amygdala function and morphology in adolescents. Consistent with previous work in adults (Bigos et al. 2010; Wessa et al. 2010; Jogia et al. 2011), we found that adolescents homozygous for the A allele of rs1006737 exhibited increased amygdala activation during processing of negative relative to neutral stimuli compared with G allele carriers. In addition, when instructed to engage in cognitive reappraisal strategies to reduce their emotional response to negative stimuli rather than simply viewing negative stimuli, rs1006737 A allele homozygotes exhibited decreased amygdala activation compared with G allele carriers. Whereas prior research has utilized tasks involving processing of emotional stimuli, this is one of the first studies to demonstrate a relation between CACNA1C genotype and amygdala activation during a task involving regulation of one's emotional responses to valenced stimuli. Together, these results suggest that, in adolescents, rs1006737 A allele homozygote status is associated with greater overall amygdala reactivity. Our findings indicate that these CACNA1C-related functional differences in the amygdala are observed earlier in development and not just in adulthood.
Some researchers have observed a gene-dose effect between the number of A alleles and amygdala activation during emotional processing (Wessa et al. 2010; Jogia et al. 2011). However, similar to Bigos et al. (2010), we only observed a significant CACNA1C genotype-amygdala activation relation for A allele homozygotes. Additional research is needed to better understand how rs1006737 genotype relates to amygdala activation. Moreover, although some prior research in adults has linked the A allele of rs1006737 to decreased functional connectivity within corticolimbic frontotemporal systems (Wang et al. 2011; Radua et al. 2013), these findings were observed during emotional processing, rather than effortful emotion regulation. Further research examining how CACNA1C variation relates to amygdala reactivity to a range of emotional stimuli and regulation instructions over the course of development is needed to better understand these processes. In addition, we found somewhat smaller effect sizes in our sample of adolescents compared with what has been observed in some studies of adults. For example, rs1006737 genotype accounted for 7% of the variance in BOLD signal in the amygdala during emotion reactivity trials and for 12% of the variance in BOLD signal in the amygdala during emotion regulation trials. In contrast, some research in adults has observed medium to large effect sizes when examining rs1006737 genotype and neural phenotypes (e.g., Wessa et al. 2010; Jogia et al. 2011; Perrier et al. 2011). Nevertheless, other neuroimaging genetics research, including genome-wide association meta-analyses (e.g., Stein et al. 2012), has also detected modest effect sizes, and has emphasized the complexity of genetic influences on neuroimaging intermediate phenotypes.
Whereas several studies have examined brain structure and function associated with rs1006737, to our knowledge, this is the first investigation of neural correlates of the rs4765914 SNP. rs4765914 T allele carriers exhibited reduced amygdala volume compared with C allele homozygotes, over and above total intracranial volume. Prior research has found that the A allele of rs1006737 was associated with increased amygdala volume (Perrier et al. 2011; Wolf et al. 2014), but no previous neuroimaging genetics studies with CACNA1C have investigated the rs4765914 SNP. To date, multiple GWAS have found several variants in intron 3 of CACNA1C to be associated with risk for a number of psychiatric disorders characterized by limbic system dysfunction, including bipolar disorder, schizophrenia, and depression (Green et al. 2009; Psychiatric GWAS Consortium Bipolar Disorder Working Group 2011), and reduced amygdala volume is observed in adults and youth with bipolar disorder (Doty et al. 2008; Kalmar et al. 2009). Replication is needed, but our findings suggest that rs4765914 may be associated with structural variation that has been linked to elevated risk for psychopathology, particularly bipolar disorder, and that these structural differences may be present in adolescence.
We did not detect any interactions between CACNA1C genotype and childhood trauma in predicting amygdala structure and function, suggesting that CACNA1C-related differences in amygdala volume and activation are not contingent upon early adversity. Together with the extant literature, our results suggest that CACNA1C-related differences in amygdala morphology and function may represent preexisting factors that are not dependent upon early experience. However, negative life experiences could exacerbate preexisting variation in amygdala structure or function related to genetic risk to lead to the onset of psychopathology. Future research should examine this question in larger samples, and longitudinally across development, to better understand the joint contributions of genetic and environmental risk factors for psychopathology.
Study findings need to be considered in light of several limitations. Our sample size was small, and we may have been statistically underpowered to detect associations, particularly interactions. Our sample was also racially and ethnically diverse, and population stratification (the presence of systematic differences in allele frequencies as a function of subpopulations in the sample; e.g., Pritchard and Rosenberg 1999) is a concern. This concern is partially mitigated because 1) findings emerged when covarying white/nonwhite race/ethnicity, 2) results remained similar in effect size when covarying principal components from a principal components analysis of ancestry-informative markers in a subset of the sample, and 3) race/ethnicity was not differentially related to CACNA1C genotypes or amygdala volume and activation. Prior neuroimaging genetics research with CACNA1C in adults has been restricted to white samples (usually of European descent) in order to minimize population stratification artifacts (e.g., Bigos et al. 2010; Perrier et al. 2011; Wolf et al. 2014). Therefore, more research using larger samples of different ancestral backgrounds is needed to clarify whether these associations hold across race/ethnicity. An additional limitation is that another genetic marker in linkage disequilibrium with the SNPs that we examined could account for the observed associations. As such, our results suggest that variation in CACNA1C is associated with amygdala function and structure, but that potentially these variants are not causative, but are, instead, associated. Furthermore, we found that only rs1006737 genotype was significantly associated with amygdala activation during emotional processing, whereas only rs4765914 genotype was significantly associated with amygdala volume. However, despite both being in intron 3 of CACNA1C, these SNPs did not exhibit substantial linkage disequilibrium. Additionally, the associations with amygdala structure and function were in the same direction for both SNPs, even though the magnitudes of these associations differed.
Conclusions
Despite these limitations, our findings suggest that underlying common genetic risk manifests as differences at the neural level—both in terms of amygdala structure and function—as early as adolescence. These findings have implications for understanding vulnerability to psychopathology. CACNA1C is expressed in the brain (among other areas), and it codes for the α1c subunit of the LTCC (Catterall et al. 2003). Researchers have proposed that alterations in CACNA1C gene expression may impact brain structure and function by influencing neuronal development and survival (e.g., through effects on cAMP response element-binding protein [CREB]-dependent transcription) (Bhat et al. 2012). Furthermore, LTCCs have been found to play a role in the amygdala in fear conditioning, and protein expression of the α1c subunit of the LTCC in the amygdala has been found to be elevated in fear-conditioned animals (Shinnick-Gallagher et al. 2003). It is of interest for future research to investigate how underlying genetic risk and associated differences in amygdala structure and function may translate into the development of different psychiatric disorders. The neural correlates of CACNA1C variants may interact with various environmental experiences to contribute to the development of different psychiatric disorders, and mapping out patterns of multifinality may shed light on mechanisms of risk and resilience associated with different forms of psychopathology. Additional research on the role of the risk variants identified in intron 3 of CACNA1C to date is also of interest. For example, a SNP in substantial linkage disequilibrium with rs1006737 has been associated with differential levels of mRNA expression in human postmortem brain samples (Bigos et al. 2010), suggesting that this variant may be involved in regulating gene expression. In sum, our findings are the first to suggest that CACNA1C-related differences in amygdala structure and function are observed by adolescence, and they provide a foundation for subsequent developmentally informed research on how underlying genetic vulnerability translates into risk for psychopathology across multiple units of analysis.
Clinical Significance
At present, clinical applications for treating psychiatric disorders based on genotype are limited. The associations between CACNA1C genotype and amygdala structure and function in our sample of adolescents were relatively small in effect size, and our findings do not have direct clinical implications currently. Further research is needed to determine whether these results are robust and able to inform clinical interventions. However, it is possible that our findings of CACNA1C-related differences in amygdala structure and function, detected as early as adolescence, may provide targets for further study in order to better understand mechanisms underlying the development of various forms of psychopathology.
Footnotes
Disclosures
No competing financial interests exist.