
Abstract
Objective:
The purposes of this study were to examine the impact of 3 weeks of amphetamine administration on intrinsic connectome-wide connectivity patterns in adults with attention-deficit/hyperactivity disorder (ADHD) and explore the association between stimulant-induced symptom improvement and functional connectivity alteration.
Methods:
Participants included 19 adults (age 20–55 years) diagnosed with ADHD using the Diagnostic and Statistical Manual of Mental Disorders, 4th ed., Text Revision (DSM-IV-TR) criteria per the Adult Clinician Diagnostic Scale taking part in amphetamine trials. For each patient, two 6-minute resting-state functional magnetic resonance imaging (R-fMRI) scans were acquired at baseline and after treatment. A fully data-driven multivariate analytic approach (i.e., multivariate distance matrix regression [MDMR]) was applied to R-fMRI data to characterize the distributed pharmacological effects in the entire functional connectome. Clinical efficacy was assessed using ADHD rating scale with adult prompts and the Adult Self-Report Scale v1.1 Symptom Checklist. We linked stimulant-induced functional connectivity changes to symptom amelioration using Spearman's correlation.
Results:
Three weeks of administration of a stimulant significantly reduced ADHD symptoms. MDMR-based analyses on R-fMRI data highlighted the left dorsolateral prefrontal cortex (DLPFC, a key cognitive control region) and the medial prefrontal cortex (MPFC, the anterior core of default network) whose distributed patterns of functional connectivity across the entire brain were altered by psychostimulants. Follow-up intrinsic functional connectivity revealed that stimulants specifically decreased the positive functional connectivity between DLPFC–insula, DLPFC–anterior cingulate cortex, and MPFC–insula. Importantly, these functional connectivity changes are associated with symptom improvement.
Conclusion:
These results suggested that ADHD is associated with increased functional integration or decreased functional segregation between core regions of cognitive control, default, and salience networks. The apparent normalization of intrinsic functional interaction in these circuits (i.e., increased functional segregation) may underlie the clinical benefits produced by 3 weeks of amphetamine treatment.
Introduction
A
Behaviorally, stimulants robustly reduce core clinical symptoms (Faraone and Glatt 2010). The therapeutic effects of stimulants are likely mediated by increases in catecholamines (i.e., dopamine and norepinephrine) via inhibition of their uptake or increase of their release within brain regions closely related to ADHD (e.g., prefrontal cortex [PFC] and striatum) (Arnsten 2006; Del Campo et al. 2011). As neuroimaging techniques advance, understanding the neural consequences of stimulant therapy at the macroscopic level should be attainable.
Task-based functional magnetic resonance imaging (fMRI) studies of children with ADHD have suggested that, at least for a subset of regions exhibiting ADHD-related differences, methylphenidate (MPH) administration changes patterns of brain activity and connectivity during task performance. In particular, a growing literature reports normalization of function in task-related networks (e.g., fronto–striato–parieto–cerebellar circuits) and the default network (a network of medial wall-based brain regions typically deactivated during goal-directed tasks) (Schweren et al. 2013; Spencer et al. 2013; Rubia et al. 2014). Although fewer in number, fMRI studies of ADHD in adults have also highlighted normalization of function (Epstein et al. 2007; Bush et al. 2008). Still, stimulant effects may differ between children and adults. For example, in a single study, MPH increased frontostriatal and cerebellar activation in children with ADHD, whereas it increased striatal activation but decreased frontoparietal activation in adults with ADHD (Epstein et al. 2007). Thus, it is critical to independently study the neural systems underlying stimulant effects to better guide the treatment of adult patients.
A central challenge for integrating stimulant effects in task-based fMRI findings is that the results obtained depend on specific experimental settings, making it difficult to compare them directly. Although a few regions have been implicated in therapeutic effects across tasks [e.g., PFC, anterior cingulate cortex (ACC), and striatum in Spencer et al. (2013)], large heterogeneity exists in terms of brain regions involved and the direction and magnitude of effects (Schweren et al. 2013). For example, stimulant-induced brain functional changes during a working memory task and an emotion task are likely to differ. Brain responses to stimulants also depend on task difficulty (Shafritz et al. 2004) and context (rewarded vs. nonrewarded) (Rubia et al. 2009). Furthermore, even in the same experiment, interindividual differences in performance may obscure treatment effects.
Resting-state fMRI (R-fMRI) is a powerful complement to task-based fMRI, which removes dependencies in findings associated with cognitive tasks. A substantial literature has demonstrated its ability to detect ADHD-related differences in functional connectivity patterns (Castellanos and Proal 2012). For example, an influential theory of ADHD pathogenesis, the default network dysregulation hypothesis, emphasizes the role of impaired coordination between task-positive and default networks in attentional lapses in ADHD (Sonuga-Barke and Castellanos 2007). To date, R-fMRI investigations of stimulant effects on ADHD are limited to two studies in youth: one examined striatum functional connectivity using seed-based correlation analyses (Hong et al. 2015) and another only examined local connectivity using regional homogeneity (ReHo) (An et al. 2013). The goal of the present work was to explore the effects of 3-week amphetamine treatment (dextroamphetamine/amphetamine extended release or lisdexamfetamine [LDX]) on whole-brain functional interactions in adults with ADHD by applying multivariate distance matrix regression (MDMR) to R-fMRI data (Shehzad et al. 2014).
Neurobiological evidence supports the notion that the pathophysiology of ADHD involves abnormal functional interactions among large-scale brain systems (Castellanos and Proal 2012; Cortese et al. 2012). Theoretically, systemically administered medications are likely to have complex distributed effects (Del Campo et al. 2011). The MDMR approach is sensitive to spatially distributed information as it is based on the functional interactions among voxels. Thus, compared to massive univariate analysis methods (e.g., seed-based correlation analysis or ReHo), this data-driven multivariate approach may be more sensitive to pharmacological effects, providing a more comprehensive characterization of brain–treatment relationships. To understand the brain mechanisms by which stimulants change behavior, we also linked stimulant-induced functional connectivity changes to behavioral amelioration.
Materials and Methods
Participants
Participants included 19 adults with ADHD (age 20–55 years) from two rater-blind clinical trials. Fourteen patients were in a study comparing adherence and efficacy between immediate-release mixed amphetamine salts (MAS IR) and extended-release MAS (MAS XR). Flow of patients is shown in Figure 1. Five patients were in a study evaluating effectiveness and duration of effect of LDX dimesylate (Vyvanse®) in adults with ADHD. In both studies, ADHD was diagnosed using the Diagnostic and Statistical Manual of Mental Disorders, 4th edition, Text Revision (DSM-IV-TR) (American Psychiatric Association 2000) criteria per the Adult Clinician Diagnostic Scale (ACDS v1.2; Adler and Cohen 2004). Other inclusion and exclusion criteria were detailed by Adler et al. (2011, 2013). ADHD patients were scanned twice: at baseline and after 3 weeks of stimulant treatment. Healthy controls (HCs) were scanned once at baseline.
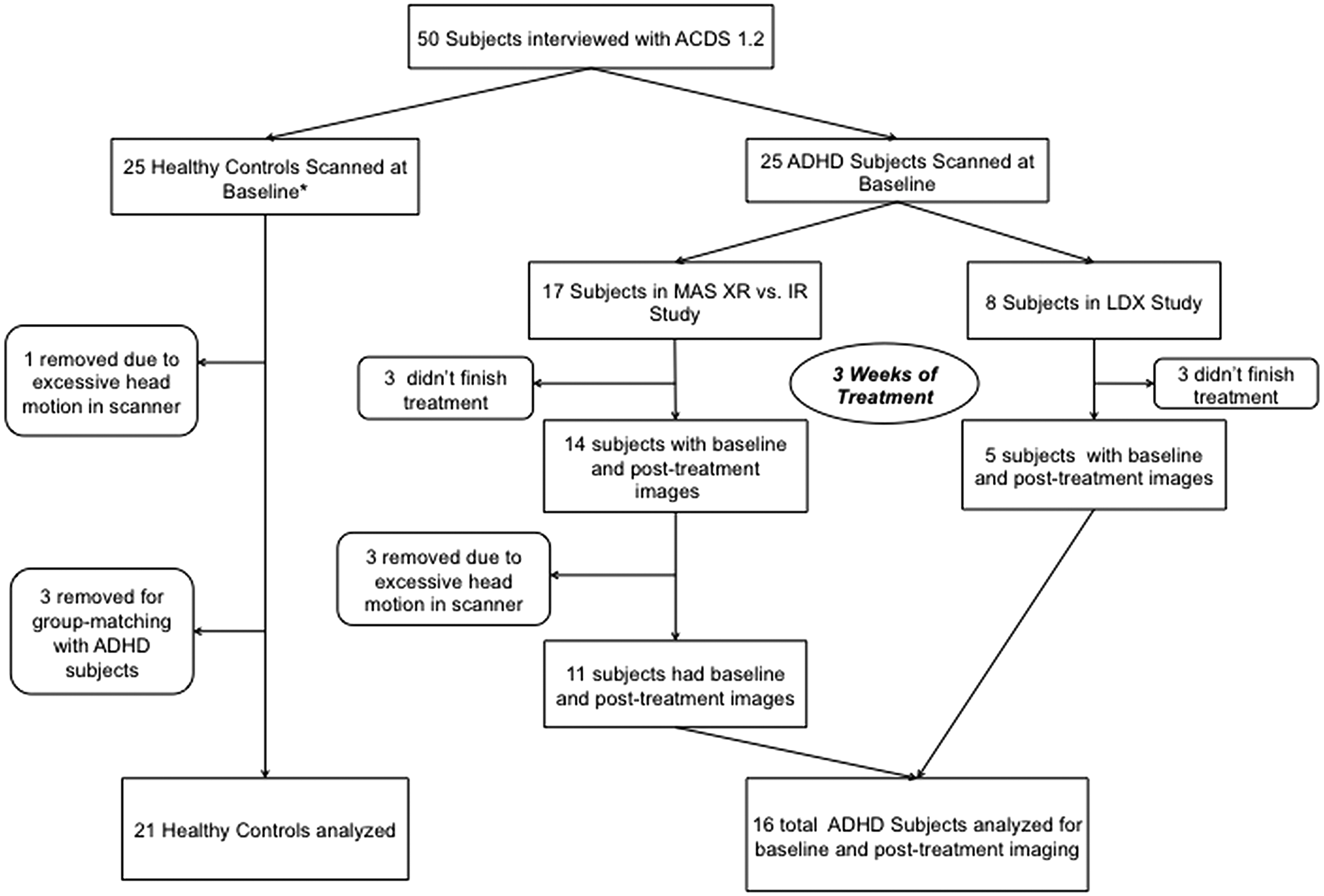
CONSORT diagram showing study subject flow. *Downloaded from 1000 Functional Connectomes Project. ACDS, Adult Clinician Diagnostic Scale; ADHD, attention-deficit/hyperactivity disorder; IR, immediate release; LDX, lisdexamfetamine; MAS, mixed amphetamine salts; XR, extended release.
Three MAS XR patients were excluded due to excessive scanner head motion: >50% of volumes with framewise displacement (FD) (Jenkinson et al. 2002) >0.2 mm on either scan or mean FD above three times the standard deviation relative to their cohort mean (ADHD: computed across all 38 scans). To group match the remaining 16 patients (11 MAS XR and 5 LDX) in age (mean: 34.3 ± 9.3 years) and sex ratio (males/females: 11/5), 21 (mean age: 34.1 ± 9.3 years; males/females: 14/7) were selected from the publicly released data set collected in an ADHD study performed at the NYU Child Study Center (downloaded from the 1000 Functional Connectomes Project database via
The study was approved by the NYU School of Medicine Institutional Review Board. Study procedures were consistent with the Declaration of Helsinki and Good Clinical Practice guidelines. All participants provided written informed consent before completing any study-related procedures.
Study design
The MAS study consisted of two 3-week treatment periods during which patients received MAS IR and MAS XR formulations with an intervening 7-day washout period (ClinicalTrials.gov identifier NCT00468143). The order of MAS IR or MAS XR was randomized and counterbalanced. Imaging data were collected during the last day of the washout period (baseline) and the last day of MAS XR treatment (treated). The LDX study (NCT01101022) consisted of 12 weeks of treatment: a 4-week phase with weekly clinic visits and an 8-week maintenance phase with biweekly clinic visits. Imaging data were collected at baseline and after 3 weeks of treatment to match the MAS XR study.
At baseline, eligible participants were initiated on MAS XR at a dose of 15 mg daily and LDX at a dose of 30 mg/day. At any given treatment visit, participants who did not achieve 30% reduction in baseline ADHD rating scale (ADHD-RS) total score received an increase of 15 mg/day for MAS XR or 20 mg/day for LDX; maximum doses MAS XR: 45 mg/day and LDX: 70 mg/day. The second scan was done on a stable dosing of MAS or LDX and ∼6 hours (6.1 ± 1.6 hours) after taking that day's medication.
Treatment efficacy measures
Clinical efficacy was assessed using ADHD-RS with adult ADHD prompts and the Adult Self-Report Scale (ASRS) v1.1 Symptom Checklist. The treatment effect score was computed as the difference between posttreatment and off-medication baseline. For the MAS XR study, the baseline measure was obtained by averaging the initial prestudy baseline and the baseline obtained after the 7-day washout period to account for potential carryover effects.
The ADHD-RS with adult ADHD prompts, a semistructured scale designed to assess current symptomatology (DuPaul et al. 1998; Adler et al. 2009), includes the 18 Diagnostic and Statistical Manual of Mental Disorders, 4th edition (DSM-IV) (American Psychiatric Association 1994) ADHD items scored on a 4-point scale (0 = not at all, 1 = mild, 2 = moderate, and 3 = severe). The ADHD-RS is composed of two subscales (inattentive and hyperactive/impulsive). Embedded in the scale are 6–10 prompted questions, using adult-specific language, for each of the 18 symptoms to ensure adequate exploration of the breadth and impact of these 18 symptoms in an adult. The ASRS v1.1 Symptom Checklist was developed to provide a self-assessment (Kessler et al. 2005; Adler et al. 2006) rating the 18 symptoms using adult-specific language in a frequency-based Likert scale (0 = never, 1 = rarely, 2 = sometimes, 3 = often, and 4 = very often). Participants completed these scales based on the preceding month at baseline and the interval since the previous visit at all other visits.
MRI data acquisition
We used a Siemens Allegra 3.0 Tesla scanner (Siemens) at the NYU Center for Brain Imaging. Two 6-minute resting scans were acquired for patients at baseline and after treatment, and one 6-minute resting scan was acquired for HCs. During the scan, participants were instructed to rest with their eyes open. Each scan comprised 192 contiguous whole-brain functional volumes acquired using a multiecho echo-planar imaging (EPI) sequence (effective TE = 30 milliseconds; TR = 2000 milliseconds; flip angle = 90°; 33 slices; voxel size = 3 × 3 × 4 mm; and field of view [FOV] = 240 × 192 mm). A high-resolution T1-weighted anatomical image was also acquired using a magnetization-prepared gradient-echo sequence (MPRAGE, TR = 2530 milliseconds; TE = 3.25 milliseconds; TI = 1100 milliseconds; flip angle = 7°; 128 slices; FOV = 256 mm; and acquisition voxel size = 1.3 × 1.3 × 1.3 mm).
Imaging preprocessing
Imaging data were preprocessed using an alpha version of the Configurable Pipeline for the Analysis of Connectomes (CPAC 0.3.9,
MDMR analysis
MDMR was performed using R package connectir (
Voxelwise MDMR analyses included two steps (see Shehzad et al. 2014 for a detailed procedure). To reduce the computational demands, we restricted our analyses to gray matter voxels (in 3 mm3 MNI space) present in all patients (100% gray matter mask). For a given voxel, step 1 includes creating a whole-brain connectivity map for each participant and comparing the similarity between the connectivity maps of all possible pairings of participants using spatial correlation. This results in an n × n distance matrix (n = 16 of participants). In step 2, MDMR was performed using these distance matrices to evaluate the complex multivariate pattern of connectivity associated with psychostimulant treatment across subjects while controlling for in-scanner head motion. This yielded a pseudo-F statistic, the significance of which (p-value) was determined using permutation testing (15,000 permutations). This procedure was repeated for each gray matter voxel, yielding a voxelwise significance map. Multiple comparisons were corrected using Gaussian random field (GRF) theory (voxel level: Z > 1.65, which corresponds to p < 0.05 in an F-test; cluster level: p < 0.05).
MDMR-guided intrinsic functional connectivity analyses
The MDMR approach can reveal the presence of significant relationships between medication and whole-brain connectivity patterns for specific brain areas, but the directionality of the specific functional connections is unknown. To further characterize MDMR results, we performed follow-up seed-based functional connectivity analyses using clusters showing a significant stimulant effect in MDMR analyses as seed regions of interest (ROIs). At the participant level, the average time series of each seed was correlated with all voxels within the 100% gray matter mask using Pearson's correlation. These correlation coefficients were then Fisher Z transformed to promote normal distribution. At the group level, voxelwise general linear model was performed on those functional connectivity maps with medication as a within-subjects factor and mean FD as a nuisance variable. Voxelwise significance map was corrected for multiple comparisons using GRF. Analyses were Bonferroni corrected for the number of seed ROIs (at the cluster level [Z > 2.33; p < 0.025]).
Seed-based functional connectivity analyses identified clusters that exhibited a significant treatment effect. The mean connectivity of each cluster was computed for ADHD patients at baseline (ADHD-baseline) and posttreatment (ADHD-treated). A paired sample t-test was performed to compare mean functional connectivity of ADHD-baseline with that of ADHD-treated to confirm the treatment effect. To examine the potential normalization effects of stimulants in patients, we also computed the mean functional connectivity of those clusters in HC and performed two independent sample t-tests to compare ADHD-baseline with HCs and ADHD-treated with HCs. To examine the association of changes in brain functional connectivity and clinical improvement, we correlated change scores (Δ = ADHD-treated − ADHD-baseline) of functional connectivity for each cluster with those in ADHD-RS (total and subset scores) and ASRS (total score).
Results
Clinical rating scales
As expected, MAS and LDX treatments significantly improved patients' ADHD symptoms on the ADHD-RS [total score: t (15) = 6.99, p = 0.000004; mean reduction relative to baseline: 37.4% ± 22.1%; inattentive score: t (15) = 6.29, p = 0.00002; mean reduction: 43.9% ± 26.3%; and hyperactive/impulsive score: t (15) = 2.96, p = 0.01; mean reduction: 24.7% ± 39.7%] and ASRS [total score: t (15) = 4.71, p = 0.0003; mean reduction: 32.3% ± 25.7%] (Fig. 2). These clinical responses are representative as they were similar to the magnitude of responses observed in the overall group of patients (Adler et al. 2011, 2013).
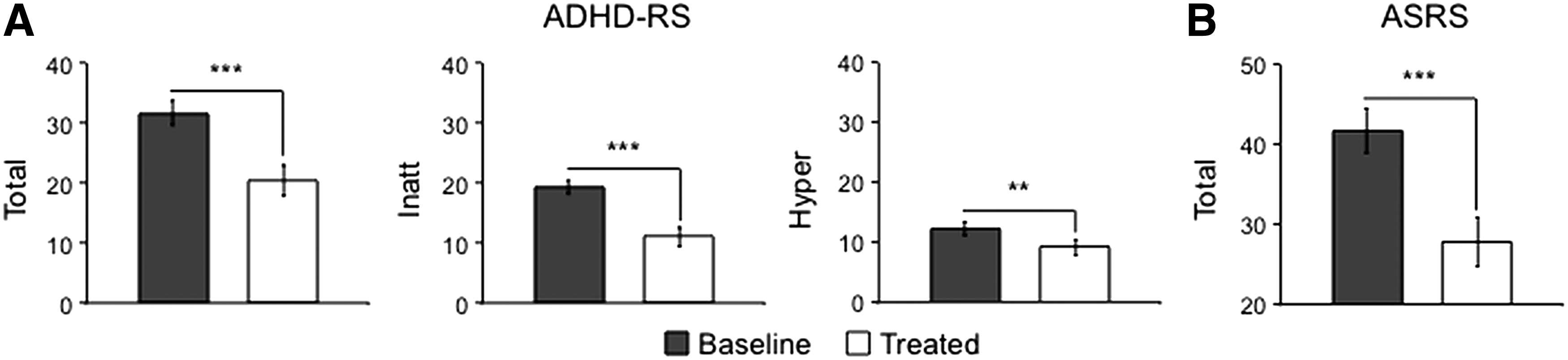
Stimulants significantly reduced ADHD symptoms measured using clinician-administered
Functional connectivity
MDMR results revealed that stimulants altered the multivariate connectivity pattern of the bilateral medial prefrontal cortex (MPFC)/paracingulate gyrus, as well as the left dorsolateral prefrontal cortex (DLPFC) (Fig. 3A and Table 1).
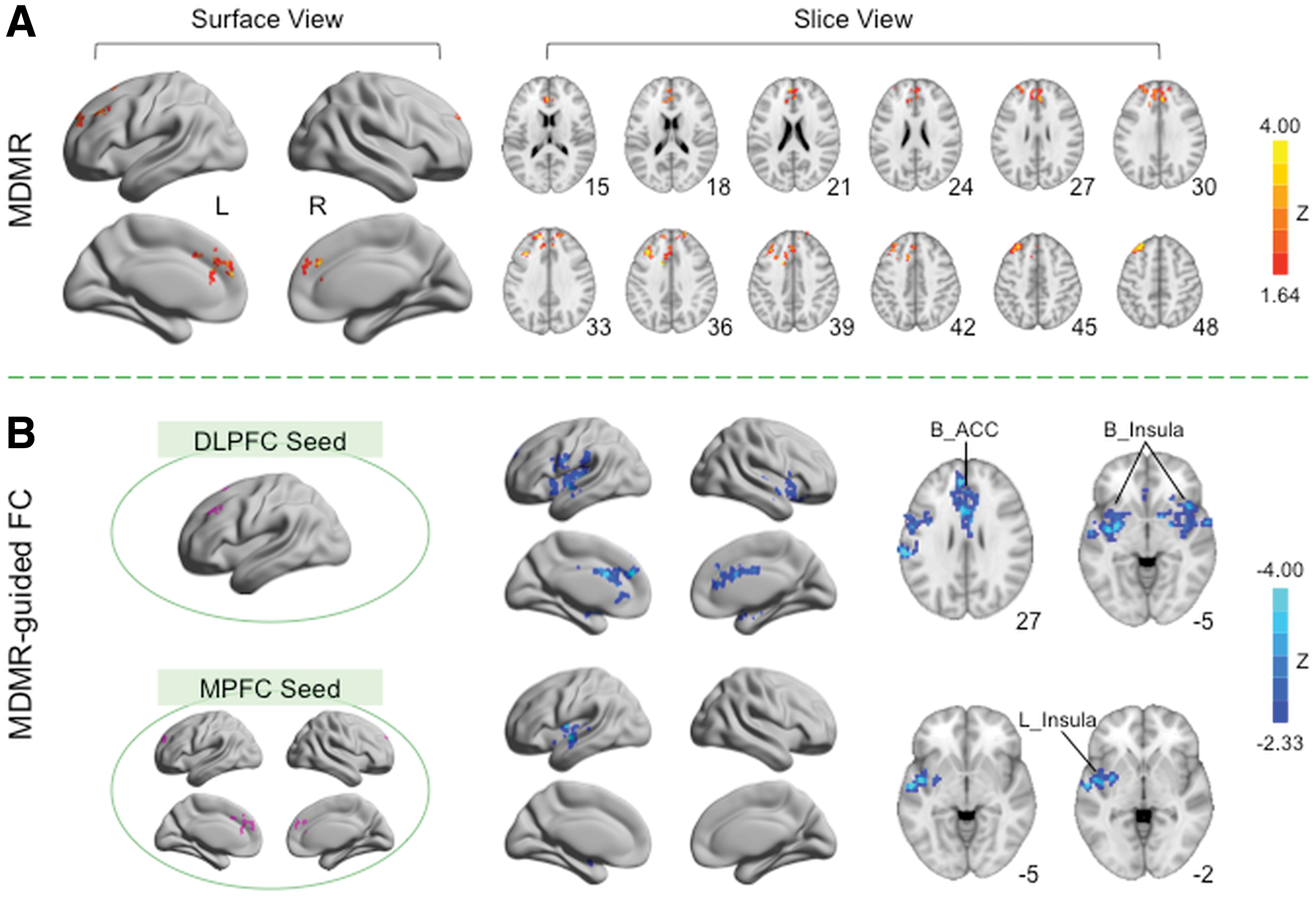
MDMR analyses demonstrated that stimulants altered the whole-brain connectivity of areas within the medial and dorsolateral prefrontal cortices (MPFC and DLPFC). The Z scores for areas exhibiting significant stimulant treatment effect in MDMR analysis are plotted on surface and slice maps
B, bilateral; L, left; BA, Brodmann area; MNI, Montreal Neurological Institute.
Based on the clusters detected using MDMR, two regions of interest were defined for follow-up seed-based correlation analysis: one located within the left DLPFC and the other within the bilateral MPFC. Follow-up functional connectivity analyses revealed that stimulants reduced the positive functional connectivity between the left DLPFC and three clusters (Z > 2.33; p < 0.025) (Fig. 3B, top row, and Table 2): (1) bilateral dorsal ACC extending into caudate, (2) the right insula extending into orbital frontal cortex and putamen, and (3) the left insula extending into surrounding areas, including precentral gyrus, supramarginal gyrus, superior temporal gyrus, medial temporal lobe, and putamen. Furthermore, stimulants also decreased the positive connectivity between bilateral MPFC and a cluster centered at the left insula extending into superior temporal lobe and putamen (Fig. 3B, bottom row, and Table 2). When contrasted to the data obtained from HC, the reductions in functional connectivity associated with stimulant therapy are consistent with “normalization” of function (Fig. 4, middle column). Specifically, the positive functional connectivity between a given seed and a cluster was significantly higher for patients compared with controls at baseline (p < 0.001). After treatment, no significant differences were observed between patients and HCs (p > 0.20).
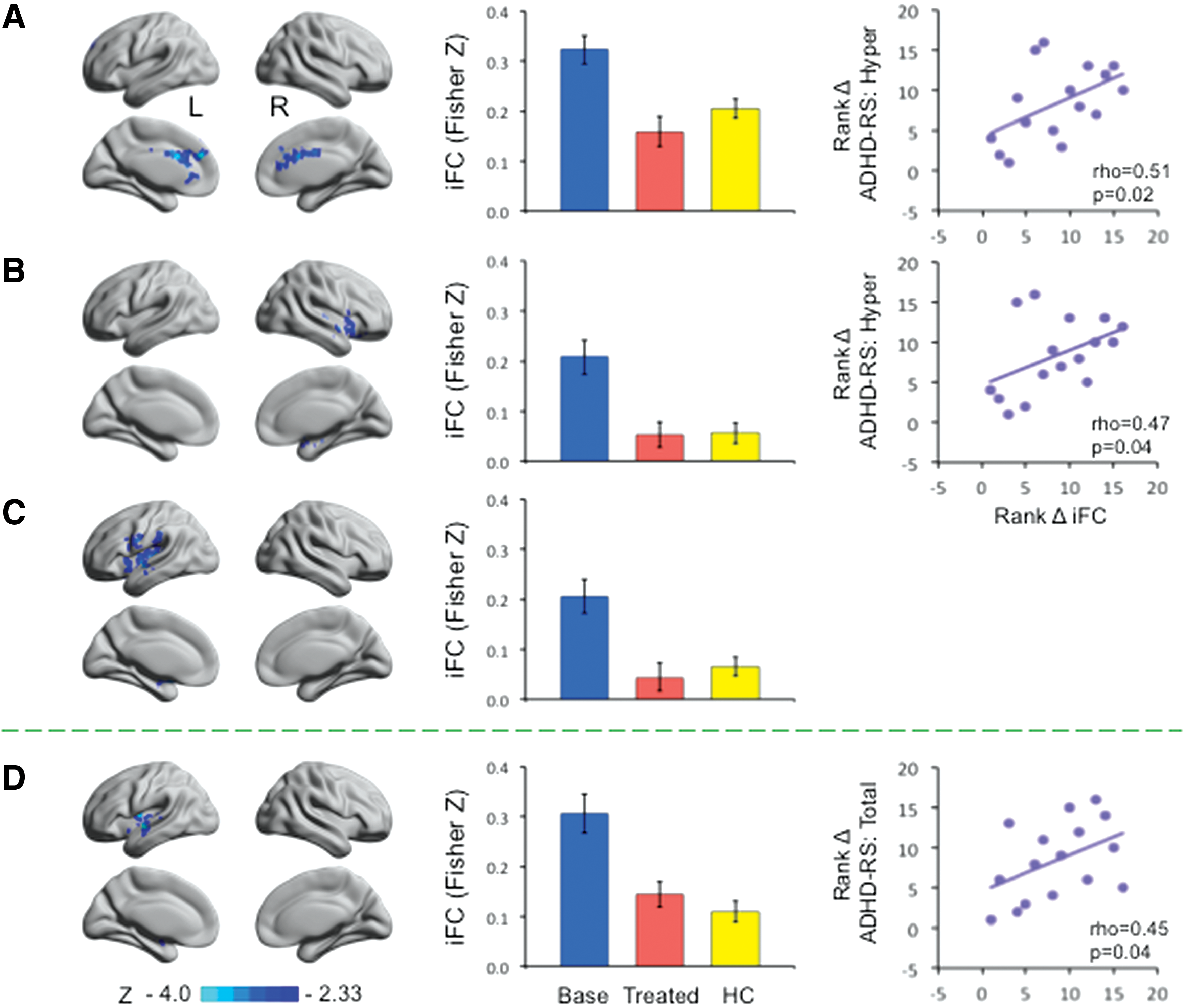
Stimulants to amphetamines reduced intrinsic functional connectivity (iFC) to normal levels and the changes in functional connectivity were associated with clinical symptom improvement. Left column: the MDMR-guided iFC analyses results shown in Figure 3B were split into clusters and represented here (DLPFC:
DLPFC, dorsolateral prefrontal cortex; MPFC, medial prefrontal cortex.
To examine the functional significance of these functional connectivity decreases, we correlated change scores of functional connectivity with those of clinical symptoms. We found that improvement in hyperactive/impulsive symptoms was significantly associated with reduction in functional connectivity (i.e., increased functional segregation) between left DLPFC and bilateral ACC (rho = 0.51, p = 0.02; one-tailed, uncorrected) and between left DLPFC and right insula (rho = 0.47, p = 0.04) (Fig. 4A, B, right column). In addition, changes in functional connectivity between bilateral MPFC and left insula/superior temporal lobe/putamen areas were significantly correlated with those in ADHD-RS total score, with a greater reduction in functional connectivity associated with a greater decrease in overall symptoms (rho = 0.45, p = 0.04; Fig. 4D, right column).
Discussion
Using a fully data-driven multivariate analytic approach, we examined the impact of 3 weeks of amphetamine administration on intrinsic connectome-wide connectivity patterns in 16 adults with ADHD participating in single-blind treatment trials. MDMR-based analyses revealed that improvements in clinical symptoms were accompanied by alterations in whole-brain connectivity patterns centered on the left DLPFC, a key cognitive control region, and MPFC, the anterior core of default network. Follow-up analyses suggested the following: (1) ADHD is associated with increases in functional synchronization or decreases in the functional segregation of these regions with key brain areas (i.e., DLPFC–insula, DLPFC–ACC, and MPFC–insula), and (2) clinical improvements in response to stimulant therapy are associated with apparent normalization of intrinsic functional connectivity in these circuits (i.e., increased functional segregation). Taken together, these findings suggest that the clinical benefits produced by amphetamine treatment may modulate functional interactions among these brain regions.
The MDMR approach used in the current work has several advantages. Unlike hypothesis-driven approaches (e.g., seed-based correlation analyses), this data-driven approach does not require a priori defined ROIs. Additionally, as a multivariate approach, it simultaneously evaluates the contributions of all possible connections of a given ROI to the treatment effect; this avoids the massive numbers of statistical tests inherent to univariate methodologies and the stringent multiple comparison corrections they entail. Unlike other data-driven multivariate approaches (e.g., ICA and graph theory-based approaches), the MDMR-based approach has minimal requirements for parameter selection (Shehzad et al. 2014) and does not require a prespecification of parameters, such as the dimensionality of the data or graph construction parameters. Thus, it effectively provides an unbiased characterization of the distributed pharmacological effects in the entire functional connectome.
In accord with previous task-based literature showing that the most consistent effect of psychostimulants on ADHD is on frontostriatal circuitry (Spencer et al. 2013; Rubia et al. 2014), we found that stimulants restored the multivariate functional connectivity pattern centered on dorsal and medial PFC. We speculate that these distributed effects may be driven by stimulant-induced increase in catecholamine levels within prefrontal neural circuits. This hypothesis is supported by neuroimaging evidence that functional connectivity involving frontal cortex is strongly modulated by catecholamines (Cole et al. 2013; Sripada et al. 2013; Mueller et al. 2014; Schrantee et al. 2015). Although our prefrontal findings are consistent with previous work, other brain regions are discrepant. For example, unlike a recent resting-state study (An et al. 2013), we did not detect stimulant effects in superior parietal, sensorimotor, or visual areas. This may be due to a limited statistical power. Alternatively, subject (e.g., age: children vs. adults), experimental (e.g., stimulants administered: MPH vs. amphetamines and treatment duration: single dose vs. 3 weeks of treatment), or methodological (e.g., ReHo vs. MDMR) factors could account for this discrepancy. Thus, these factors should be considered when unwrapping the neural substrates of psychostimulants.
Our follow-up functional connectivity analysis highlights the potential impact of stimulants on functional segregation (or functional decoupling) and the critical role of functional segregation in symptom improvement. Specifically, the abnormally enhanced functional connectivity between PFC and key regions of the executive–salience networks (i.e., ACC and insula) in patients appeared to normalize after treatment. Importantly, the magnitude of functional connectivity reduction was linked to the degree of amelioration of ADHD symptoms. These results are consistent with recent R-fMRI studies in healthy adults showing that stimulants reduced within- and between-network functional connectivity in central executive, salience, and default networks (Cole et al. 2013; Sripada et al. 2013; Mueller et al. 2014; Schrantee et al. 2015). These results are also in line with recent resting-state studies examining stimulant effects in children and adolescents with ADHD, which demonstrated that good responders to MPH were characterized by significantly reduced frontostriatal functional connectivity compared with poor responders (Hong et al. 2015).
According to the saliency mapping deficit model of psychopathology, enhanced salience–central executive and salience–default network coupling may reflect deficits in context-dependent engagement and disengagement of cognitive systems important for attending to salient external stimuli or internal mental events (Menon 2015). This is possibly due to the role of the salience network in mediating the switch between the lateral frontoparietal central executive network and the medial default network to keep attention focused on task-relevant goals. Abnormality in network interacts between salience, central executive control, and default networks were shown as a reproducible feature of childhood ADHD (Cai et al. 2015). Our findings provide further support for the saliency mapping deficit model and suggested that functional segregation may serve as a key neurobiological signature of psychostimulant therapy. The persistence of abnormal functional synchronization between these key brain areas may be an important factor in determining whether ADHD symptoms persist.
Consistent with previous work showing unique patterns of atypical connectivity for ADHD subtypes (Fair et al. 2012), we found the following: (1) decreased functional connectivity of DLPFC is associated with improved hyperactivity/impulsivity symptoms, and (2) decreased connectivity of MPFC is associated with improvement of overall symptoms (hyperactivity/impulsivity+inattentiveness). Functional segregation of DLPFC and ACC/insula may allow DLPFC to more effectively maintain representations of task demands with less influence from externally driven distracting stimuli. This increased cognitive control in turn can enhance one's ability to suppress responsiveness to irrelevant sensory stimuli in the environment and thus impulsivity. This explanation is supported by task-based fMRI studies showing that stimulants elicited compensatory activation in the PFC of children with ADHD during a rewarded continuous performance task (Rubia et al. 2009). Magnetic resonance spectroscopy studies also showed that stimulants induced significant neurochemical changes in the PFC of children with ADHD, which underpinned functional improvement (Husarova et al. 2014).
In contrast, overall symptom improvement was linked to reductions in MPFC–insula connectivity. This is consistent with previous work showing that the atypical connectivity pattern for ADHD-combined type was most prominent in midline default network components, as well as insular cortex (Fair et al. 2012). Our results are also in accord with previous resting-state fMRI studies showing stimulant-induced functional segregation between default and salience networks (Sripada et al. 2013). Consistent with the default network dysregulation hypothesis of ADHD (Mattfeld et al. 2014), decreased synchronization between these networks may represent decreased interference by default network processes in task-positive operations.
This study has several limitations. First, we used a convenience sample from two single-blind clinical trials (MAS XR and LDX); the lack of a placebo condition limits the interpretation and generalization of the results. However, the inclusion of an HC group and the observed relationship between brain responses and symptom improvement support our interpretations of the results. Second, our sample size was moderate (n = 16), and the sample was composed of participants with different ADHD subtypes (8 combined, 7 inattentive, and 1 missing information) with different treatment histories. Future studies should take into consideration these and other factors to unravel complex medication effects (Schweren et al. 2013). Third, MAS XR and LDX may have subtle differences in their impact on the brain, beyond the rate of delivery of active ingredients; this also warrants further exploration.
Conclusion
Via applying a fully data-driven multivariate analytic approach (i.e., MDMR) to R-fMRI data, the current work provided an unbiased and a comprehensive characterization of the distributed pharmacological effects of psychostimulants in the entire functional connectome. Our results suggested that stimulants reversed the abnormally decreased functional segregation between core regions of cognitive control (i.e., dorsolateral prefrontal cortex), default (i.e., medial prefrontal cortex), and salience (i.e., insula) network in ADHD. The identified brain-symptom associations suggest that brain mechanisms by which stimulants change behavior are possibly via increasing brain functional segregation.
Clinical Significance
Our connectome-wide association study provides novel insights on the neural consequences of psychostimulant therapy at the macroscopic level in adult patients with ADHD. The links observed between clinical benefits and brain functional connectivity changes are promising, suggesting the potential utility of unbiased resting-state brain measures as neural indices of treatment responses in ADHD. These preliminary data provide the groundwork for randomized, crossover placebo-controlled, as well as case-controlled clinical trials to further validate the predictive power of these intrinsic brain indices and confirm their specificity to ADHD. This knowledge can advance our understanding of action mechanisms of psychostimulants and help explain interindividual differences in treatment response, which can guide the administration of medication and help to improve treatment efficacy.
Footnotes
Acknowledgment
We thank Krishna Somandepalli for his help with transferring the data.
Disclosures
Zhen Yang, Clare Kelly, Francisco, X. Castellanos, Terry Leon, and Michael P. Milham declared no conflicts of interest in relation to this work for the past 3 years.
Three years of disclosures for Lenard A. Adler are shown below:
Grant/research: Sunovion Pharmaceuticals, Lundbeck, Purdue Pharmaceuticals, Enzymotec, Shire Pharmaceuticals, Theravance, Department of Veterans Affairs, Eli Lilly and Company, and APSARD/Pound Foundation.
Consultant and/or Advisory Board: Sunovion Pharmaceutical, Enzymotec, Alcobra Pharmaceuticals, Theravance, National Football League, Major League Baseball, Shire Pharmaceuticals, and Neurovance.
Speakers' Bureau: None.
Major stockholder: None.
Other financial or material support: Royalty payments (as inventor) from NYU for license of adult ADHD scales and training materials since 2004.