
Abstract
Objectives:
Fragile X syndrome (FXS) is the most common inherited cause of intellectual disability, in addition to being the commonest diagnosable cause of autism. The identification of the biochemical mechanism underlying this disorder has provided amenable targets for therapy. This review aims to provide an overview of investigational drug therapies for FXS.
Methods:
The authors carried out a search of clinical and preclinical trials for FXS in PubMed and on the U.S. National Institutes of Health index of clinical trials (
Results:
The review of the preclinical, Phase I, and Phase II trials of agents with therapeutic potential in FXS revolves around an understanding of the putative pathways in the pathogenesis of FXS. While there is significant overlap between some of these pathways, the agents can be categorized as modulators of the metabotropic glutamate receptor system, GABAergic agents, and miscellaneous modulators affecting other pathways.
Conclusion:
As trials involving agents targeting different aspects of the molecular biology proceed, common themes have emerged. With the great hope came great disappointment as the initial trials failed to demonstrate sufficient significance. In particular, the differences in outcome between the animal models and humans have highlighted the unique challenges of carrying out trials in these cognitively and behaviorally challenged individuals, as well as a dearth of clinically relevant outcome measures for use in medication trials. However, in reviewing and reframing the studies of the last decade, many important lessons have been learned, which will ultimately have a greater impact on therapeutic research in the field of developmental delay as a whole.
Introduction
F
Premutation carriers are defined as individuals who possess between 55 and 200 repeats. Premutation carriers can have milder cognitive and psychiatric difficulties throughout their lives as well as increased risk for late-onset disorders, such as fragile X-associated tremor ataxia syndrome (Brown and Stanfield 2015) and, in women, premature ovarian insufficiency (Allingham-Hawkins et al. 1999). As the number of repeats increases, there is genetic anticipation and potential repeat expansion in subsequent generations. As the number of repeats comes to exceed approximately 200, the gene is typically methylated, transcription is shutoff, and no FMRP is produced from that copy of the gene. This leads to the manifestations of the FXS (Pieretti et al. 1991). Females are usually, but not always, less severely affected due to X inactivation, when either the normal or the mutated X chromosome is randomly silenced, resulting in a mosaic pattern of FMRP production.
FMRP is necessary for neuronal plasticity: it is involved in RNA targeting to specialized regions for local translation of the messenger RNA (mRNA) that codify proteins involved in dendrite formation and synaptogenesis (Kao et al. 2010; Saldarriaga et al. 2014). Without FMRP, the mRNA translation stimulated by activation of metabotropic glutamate receptors (mGluR), including mGluR1 and particularly mGluR5, is altered, and synapses are unable to form and function normally (Bassell and Warren 2008). The GABAergic inhibitory system, in particular, has been shown to be downregulated in the absence of FMRP (D'Hulst and Kooy 2009).
Studies have also shown that FMRP acts to modulate the function of mRNA and miRNA (microRNA, a noncoding RNA that posttranscriptionally facilitates regulation of gene expression). Not only does it act on multiple targets, but it also has an effect in different tissue types, including brain and ovarian tissues (Ascano et al. 2012). Specifically, FMRP is involved through interaction with miRNAs in the regulation of N-methyl-D-aspartate receptor (NMDAR) translation at the synapse (Edbauer et al. 2010). FMRP also acts to stabilize certain proteins, several of which have been associated with autism phenotypes (including PTEN and SHANK3, among many others) (Betancur et al. 2009). Matrix metallopeptidase-9 (MMP-9), a protein that maintains the flexibility of synapses, has been shown to be elevated in FXS (Janusz et al. 2013).
There is currently no cure for FXS, and treatment is largely symptomatic, with medications most commonly targeted to anxiety, inattention, hyperactivity, irritability, and aggression. However, the advances in our understanding of deficits at the cellular and molecular level in FXS have led to several preclinical and clinical trials for this syndrome (Gross et al. 2015). For the purpose of this article, we have limited the review to preclinical and animal trials, Phase I, and Phase II clinical trials.
Methodology
The authors carried out a search of clinical and preclinical trials for “Fragile X” in PubMed and on the U.S. National Institutes of Health index of clinical trials (
As can be seen in Figure 1, treatment trials for FXS have targeted pathways apparently affected by FMRP. While the primary actionable consequence of FMRP deficit appears to be a perturbation in the balance of the excitatory glutamatergic and inhibitory GABAergic systems, there appear to be additional roles as well as various paths involved in producing the pathogenic effects. We have classified the studies based on the mechanism of action of the individual agents, specifically which pathways or neurotransmitter system they act on.
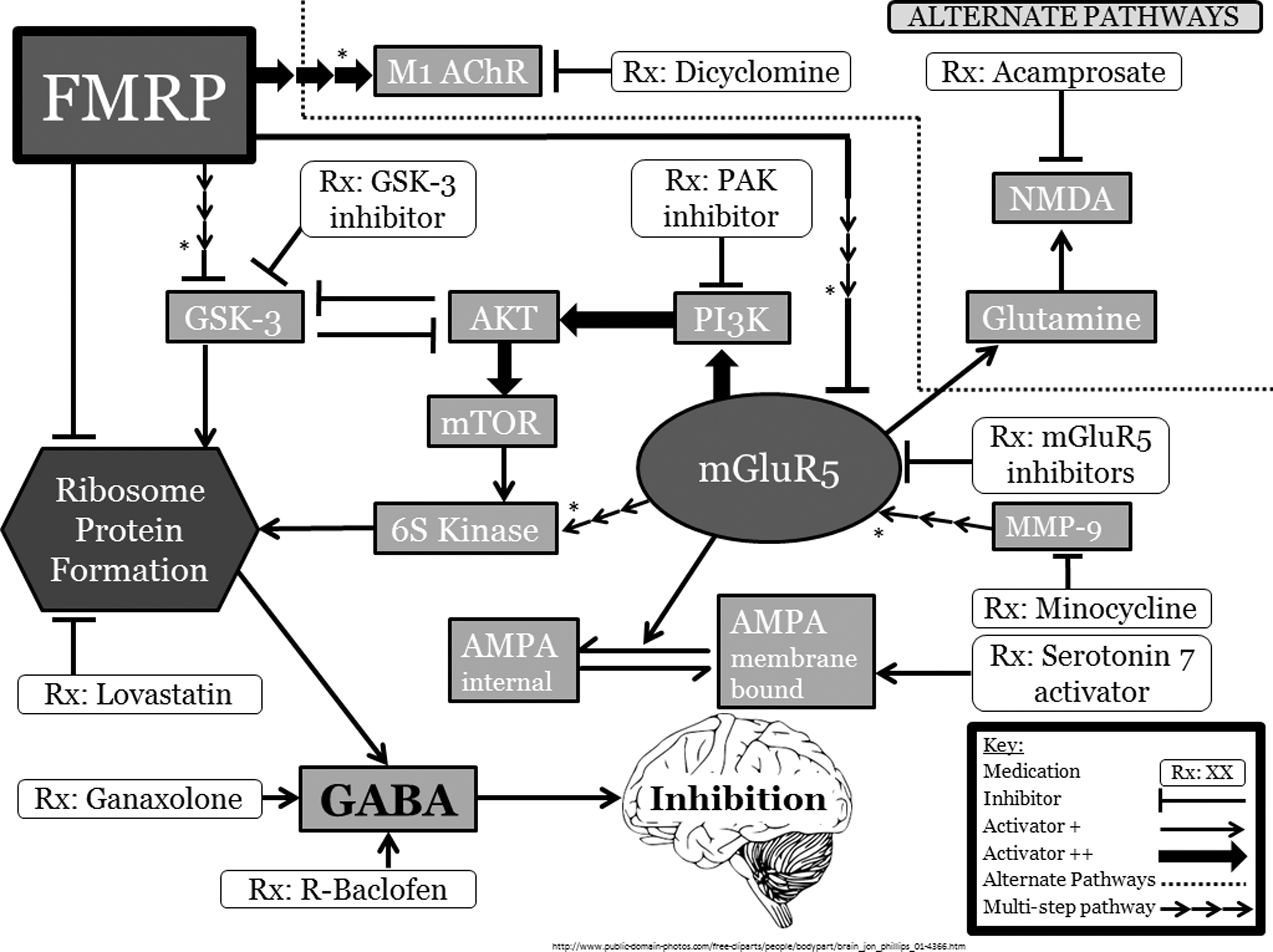
An overview of the pathways involved in FMRP expression. Please see text for complete citations and pathway explanations. *Please note that these are indirect effects; further discussion of these can be found in the text. FMRP, fragile X mental retardation protein; M1 AChR, muscarinic acetylcholine receptor 1; GSK-3, glycogen synthase kinase 3; PAK, p21-activated kinase; NMDA, N-methyl-D-aspartate receptor; AKT, protein kinase B; PI3K, phosphoinositide 3-kinase; mTOR, mammalian target of rapamycin; mGluR5, metabotropic glutamate receptor 5; MMP-9, matrix metallopeptidase-9; 6S kinase, Ribsomal S6 kinase; GABA, gamma-aminobutyric acid; AMPA, α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid.
Current Evidence
Described below is the current evidence based on preclinical and clinical studies performed, categorized by major pathways as described in a previously published review (Braat and Kooy 2014). Details, including outcome measures and specific findings, of all studies discussed in this review are listed in Tables 1 and 2.
A list of previous studies of FXS performed in mice, with the medication used, proposed mechanism of action, original publication, mouse type, and study parameters and results. For a full explanation of pathways and acronyms, please refer to the text.
FXS, Fragile X syndrome; ICSS, intracranial self-stimulation; IGF-1, insulin-like growth factor-1; mGluR5, metabotropic glutamate receptor 5; mRNA, messenger RNA.
A list of previous studies of FXS performed with human participants, with the medication used, proposed mechanism of action, original publication, age, gender, and methylation status of participants, and study parameters and results. For a full explanation of pathways and acronyms, including study questionnaires, please refer to the text.
ABC-C, Aberrant Behavior Checklist–Community Edition; ADHD-RS, attention-deficit/hyperactivity disorder rating scale; CGI-I, Clinical Global Impressions–Improvement; CGI-S, Clinical Global Impressions–Severity; CYBOCS-PDD, Children's Yale–Brown Obsessive Compulsive Scale modified for pervasive developmental disorders; EL, expressive language; FXS, fragile X syndrome; mGluR5, metabotropic glutamate receptor 5; MMP-9, matrix metallopeptidase-9; MSEL, Mullen Scales of Early Learning; NMDAR, N-methyl-D-aspartate receptor; PARS, pediatric anxiety rating scale; SRS, social responsiveness scale; SSRI, selective serotonin re-uptake inhibitor.
Modulators of the mGluR system and related pathways
Direct targeting of group 1 mGluRs
The mGlu5 receptor appears to have a central role in the pathology of FXS. In healthy individuals, FMRP regulates the activity of the mGluR pathway, driving the formation of dendritic growth and synaptogenesis in a regulated manner consistent with appropriate neuroplastic response to sensorial input. In fragile X, this controlled regulation is lost, and unregulated mGluR5 activity theoretically leads to inappropriate growth and formation of dendritic spines and synapses, and abnormal neoplastic remodeling in response to environmental stimuli (Bear et al. 2004).
MPEP, a model mGluR5 antagonist, demonstrated improvement in Fmr1-knockout mice (Fmr1 KO, a fragile X mouse model) after acute administration in young and adult mice, as evidenced by reduced audiogenic seizures and locomotor activity, as well as fewer errors and less impulsivity when tackling maze tasks (Yan et al. 2005; Gandhi et al. 2014). There are also other animal studies suggesting that treatment with MPEP reverses behavioral phenotypes in Fmr1 KO mice, as assessed by an improvement in a measure of motor learning and elimination of the manifestation of audiogenic seizures (Thomas et al. 2012; Fish et al. 2013).
In 2009, an open-label pilot study in individuals with FXS (without a control group) of fenobam (NPL2009), an mGluR5 receptor antagonist, was conducted. Twelve participants (age range 18.7–30.7 years) with FXS underwent one-time dosing of 50–150 mg of fenobam and pharmacokinetic testing. Outcome measures of prepulse inhibition and continuous performance test (CPT) were obtained pre- and postdosing to examine fenobam effects on sensory gating, attention, and inhibition. Improvement in prepulse inhibition (a marker of sensorineural gating that is impacted by fragile X) was seen in 6 participants after 1 dose, and there was clinical improvement (characterized by observations at designated time points by the principal investigator) in eye contact and/or social interaction in 8 of the 12 patients. There was no improvement in CPT with fenobam. There were no significant adverse effects to the fenobam treatment (Berry-Kravis et al. 2009).
CTEP, another mGluR5 antagonist, has a duration of action of 18 hours, longer than either fenobam or MPEP (Lindemann et al. 2011). Using this longer-acting mGluR5 antagonist in Fmr-1 KO mice, a study of behavior and physiological measures showed normalization of some brain activity and recovery of learning problems (Michalon et al. 2012). This study demonstrated correction of elevated hippocampal long-term depression, protein synthesis, and audiogenic seizures with acute CTEP treatment, as well as rescue of cognitive deficits, auditory hypersensitivity, and aberrant dendritic spine density with chronic treatment.
This was followed by the Novartis-sponsored randomized controlled trials (RCTs) of the noncompetitive, mGluR5 antagonist AFQ056 (Mavoglurant) in adolescents and adults with FXS (Vranesic et al. 2014). The initial Phase II RCT in the European Union on fully methylated individuals showed promise, because, while there was no improvement of behavioral symptoms of FXS on the primary outcome measure of the Aberrant Behavior Checklist–Community Edition (ABC-C), an exploratory analysis showed improvement on this measure in 7 patients with full FMR1 promoter methylation and no response in 18 patients with partial promoter methylation (Jacquemont et al. 2011).
Based on these findings, there followed two American and European Union RCTs involving fully and partially methylated individuals with FXS, one in adults and one in adolescents (Berry-Kravis et al. 2016). However, in both these RCTs, the difference in outcomes between mavoglurant and placebo did not reach significance, as measured by the primary end point of improvement on behavioral symptoms measured by the ABC-C using the FXS-specific algorithm. This led to the cancelation of the open-label extension study and the child study.
Overlapping in time with the initial AFQ056 study, Roche sponsored an RCT of the novel mGluR5 antagonist, R04917523 (basimglurant) in FXS, also including fully and partially methylated subjects (Hoffmann-La Roche 2012). Results again demonstrated no significant benefit from basimglurant compared with placebo. These failures, compared with the apparent success of mGluR5 inhibitors in rescuing cognitive function and behavior in mouse models, raised questions about both the validity of the mouse model and the validity of the clinical outcome measures used in the human trials. It also raised questions about when individuals with developmental disorders are most likely to respond to treatments, particularly given that the majority of subjects in the trials were post-adolescence and beyond (Gomez-Mancilla et al. 2014).
mGluR1 is also a group 1 mGluR, like mGluR5, and also has promise as a therapeutic target. DL-AP3, an mGluR1 antagonist, in combination with SKF81297, a dopaminergic (D1) agonist, has been studied as a potential drug therapy for FXS, by Xu et al. (2012), who describe the utility of combining these two agents. They report that, “mGluR1 inhibition is a useful strategy to recover D1 receptor signaling in the Fmr1 KO mice, and combination of Grp1 mGluR antagonist and D1 agonist is a potential drug therapy for the FXS”.
Nondirect targets including those downstream of group 1 mGluRs
As is typical for biological processes, FMRP impacts many interconnected pathways. Within the core mGluR–FMRP processes are a number of potential targets (Fig. 1). These include the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor pathway and the glycogen synthase kinase-3 (GSK) pathway.
Glycogen synthase kinase (GSK) is one of the neuromodulators involved in the regulation of dendritic transcript translation. In the hippocampus, FMRP indirectly regulates GSK3β, which may affect adult learning and neuronal development (Guo et al. 2012). Given the function of FMRP as an indirect regulator of GSK3 translation, supplemental inhibition of GSK3 in an Fmr-1 KO mouse was attempted in an effort to regulate neurogenesis in the hippocampus. The GSK3 inhibitor SB216763 led to improvements in learning in domains regulated by the hippocampus (Guo et al. 2012). A similar mechanism, inhibition of GSK3, has been proposed for the functional efficacy of lithium treatment in animal models of FXS. Mouse and fruit fly models have shown behavioral and physiological recovery from several fragile X phenotypes with lithium treatment (Choi et al. 2010; King and Jope 2013; Liu and Smith 2014). One open-label safety study of lithium use in adults with FXS showed some improvements in functional behavior. However, to this point, no randomized controlled trial of lithium has been carried out in individuals with FXS (Liu and Smith 2014).
Research on synaptic defects in FXS has shown insufficiency of AMPA-receptor activity (Berry-Kravis et al. 2006). As seen in Figure 1, absence of FMRP leads to enhanced mGluR-mediated dendritic translation. This, in turn, causes excessive internalization of AMPA receptors, and immature, elongated dendritic processes, affecting ribosomal protein formation (Bear et al. 2004). Given this, it was believed that AMPAkine compounds might provide a mechanism for treatment of FXS, and the AMPAkine CX516 was tested as a treatment for memory issues, which may contribute to the learning and attentional problems seen in FXS. A four-week Phase II clinical trial, however, showed no improvement to the primary outcome of memory or to the secondary outcomes, including global functioning, behavior, attention, and language. The authors pointed out that inadequate dosage, as well as a lack of appropriate end points, may have played a role in the failure of this trial, and did not feel that this negative trial ruled out the possibility of AMPA modulation as a treatment strategy for FXS (Berry-Kravis et al. 2006).
Activation of serotonin receptor 7 (5-HT7) is thought to reverse mGluR-induced ionotropic AMPA receptor internalization and long-term depression (LTD) of hippocampal synaptic plasticity, in both wild-type and Fmr-1 KO mice. Therefore, selective activation of 5-HT7 receptors was proposed as a novel strategy in the therapy of FXS (Costa et al. 2012). Osterweil et al. (2012) further suggest that the 5-HT pathway functionally interacts with the mGluR pathway and that 5-HT7 agonists can correct pathologic changes in FXS by this mechanism, as well as enhance memory formation.
Bryostatin-1, a protein kinase Cɛ activator, encourages maturation of dendrites and synapses through phosphorylation of PSD-95 (postsynaptic density protein 95). This is associated with an increase in the number of synapses and increased amplitudes of excitatory postsynaptic potentials in rat hippocampus as well as restoration of cognition in adult fragile X mice, showing an effect even after brain maturation (Sun et al. 2014; Sen et al. 2016).
GABAergic agents
Gamma aminobutyric acid (GABA) is the major neuroinhibitory transmitter; therefore, the downregulation of the GABAergic system seen in FXS leaves patients prone to seizures, anxiety, and oversensitivity, as well as behavioral and attentional problems and autistic symptoms (Watanabe et al. 2002; Darnell and Klann 2013; Lozano et al. 2014). Both GABA-A and GABA-B receptors have been investigated in animal and human studies and are described below.
GABA-A acts on a chloride ion channel receptor to reduce the neuronal firing rate when ligand is bound, by hyperpolarizing the cell. Recent work by Braat et al. (2015) supports a downregulation of GABA-A transcription in FXS, as well as evidence for the direct binding of FMRP to the mRNA molecules of the GABA-A receptor subunits. This work also demonstrated an improvement in repetitive and perseverative behavior (as assessed by digging behavior on the marble-burying assay) in Fmr1-KO mice with ganaxolone, a GABA-A-positive allosteric modulator. In previous studies that investigated the GABAergic system, Fmr-1 KO mice treated with the GABA-A agonists, diazepam and alphaxalone, showed improved motor coordination (using the rotarod test), and overall improved performance on maze tests (due to an anxiolytic effect); treatment with ganaxolone showed prevention of seizure activity induced by the audiogenic seizure tests in Fmr-1 KO mice (Heulens et al. 2012). Unfortunately, however, the picture is not clear; taurine, another GABA-A agonist and modulator, has not shown efficacy in improving the processing capabilities of the hippocampus of Fmr-1 KO mice, as a function of increasing interstimulus intervals (ISI). This may be because the Fmr-1 KO mice have fewer binding sites for taurine on the GABA-A receptors compared with the wild-type mice (El Idrissi et al. 2010).
Based on the successes with some modulators, Heulens et al. (2010) concluded that GABA-A was a potential therapeutic target for FXS, and additional compounds are being considered for investigation (Tranfaglia 2015). Currently, there is a Phase II trial of the GABA-A agonist ganaxolone in children and adolescents with FXS, investigating its efficacy for the treatment of attention problems and anxiety (Marinus Pharmaceuticals; Lozano et al. 2014). This proof-of-concept, double-blind, randomized, placebo-controlled crossover study also aims to assess the safety, tolerability, and efficacy of ganaxolone and is currently recruiting subjects with FXS aged 6–17 years. The study uses several neuropsychological and psychometric outcome measures, including the Clinical Global Impressions–Improvement (CGI-I) scale, Pediatric Anxiety Rating Scale, and KiTAP (test of attentional performance for children).
GABA-B receptors, unlike GABA-A, are G protein-coupled receptors, with a slower cell hyperpolarization than GABA-A type, because they act through potassium channels. GABA-B agonists, such as R-baclofen and GS-39783, were shown to reduce induced audiogenic seizures in Fmr-1 KO mice (Pacey et al. 2009). In Fmr-1 KO mice, R-baclofen rescued the increased susceptibility to seizures and corrected the excessive dendritic spine density and protein synthesis believed to be pathogenic in FXS (Henderson et al. 2012).
This led to an investigation of arbaclofen (a GABA-B agonist) and its effect on neurobehavioral function, including irritability, in individuals with FXS. In a randomized, double-blind, placebo-controlled crossover study of 63 individuals (55 males and 8 females) aged 6–39 years with full mutation fragile X, no effect was found on the primary end point of irritability, as assessed using the ABC-C Irritability subscale. However, global measures and post hoc analyses showed behavioral benefits, and the drug was well tolerated (Berry-Kravis et al. 2012). On post hoc analysis with the ABC-C Social Avoidance scale, the authors reported a significant beneficial treatment effect in the full study population. Based on these findings, the authors concluded that in spite of not showing a benefit on irritability, arbaclofen may potentially improve social function and behavior in individuals with this condition. Of note, a more recent animal study demonstrated reduced deficits on a test of social behavior in Fmr1 KO mice (Qin et al. 2015). The drug manufacturer began Phase III trials for FXS with social withdrawal as the primary outcome measure, but the results of those trials are not published, to the best of our knowledge. It should be noted that later trials were stopped by Seaside Therapeutics due to resource limitations before the end points were reached.
Given animal research implicating glutamate and GABA receptors in the symptomatology of FXS, riluzole emerged as a medication of interest. It is used to treat amylotrophic lateral sclerosis because of its hypothesized attenuation of excitotoxicity induced by glutamate (Obrenovitch 1998). The reduction in excitatory glutamate effects and the enhancement of GABA-A effects are theoretically desirable in treating FXS. However, in a small open-label pilot study of adults with FXS, only one of six subjects demonstrated a clinical response on outcome measures of the CGI-I and Child Yale–Brown Obsessive Compulsive Scale (CYBOCS), which measured reductions in repetitive checking, questioning, touching, and picking (Erickson et al. 2011b).
Memantine has activity at a number of receptors, notably glutamate, and has possible theoretical efficacy in the treatment of FXS (Chen et al. 1992). A retrospective chart review of six individuals with FXS and a comorbid autism spectrum disorder, who were taking memantine, showed a global benefit for four participants. The remaining two participants stopped treatment due to irritable behavior (Erickson et al. 2009).
Acamprosate, a medication approved for maintenance treatment of alcohol use disorder through possible glutamate and GABA modulation, may have a beneficial effect for FXS through a complex and controversial mode of action. This agent is thought to act at the NMDAR through unclear mechanisms (Berton et al. 1998), act as a weak GABA-A agonist (Mann et al. 2008), and possibly even have mGluR5 antagonism (Erickson et al. 2010). An open-label trial by Erickson et al. (2013) showed social improvement and reduction in inattention and hyperactivity over a 10-week period in children and adolescents aged 6–17 years with FXS. Nine of 12 subjects in the study, after treatment with acamprosate (mean dose: 1054 ± 422 mg/day), showed treatment response, as defined by CGI-I scale score of “very much improved” or “much improved.” Outcome measures included the CGI-I, ABC-C, Social Responsiveness Scale, compulsion scale of Children's Yale–Brown Obsessive Compulsive Scale modified for pervasive developmental disorders (CYBOCS-PDD), Clinical Global Impressions–Severity, and ADHD rating scale. Acamprosate was well tolerated with no significant adverse effects. The study also identified brain-derived neurotrophic factor (BDNF) as a potential biomarker reflecting efficacy of the treatment (Erickson et al. 2013).
Miscellaneous modulators affecting other pathways
While the main research focus for investigational drugs for FXS is on effectors of the abovementioned pathways, there has also been interest in agents affecting alternate pathways. These pathways have been postulated as a result of animal and human studies detailed below (included are those agents for which preclinical, Phase I, or Phase II trials have been conducted) as well as extrapolations from studies mentioned earlier, indicating overlapping pathways.
Antioxidants
The prevention of oxidative stress by antioxidants has been proposed as an approach for clinical trials for the treatment of FXS. Antioxidants, including melatonin, vitamin C, and vitamin E, have been tested as possible treatments for FXS based on data showing high levels of free radical production in the brain of Fmr-1 KO mice (el Bekay et al. 2007). When treated chronically with melatonin, Fmr-1 KO mice show positive behavioral changes and improved learning. The mice had reduced glutathione levels and lower rates of lipid peroxidation compared with untreated Fmr-1 KO mice (Romero-Zerbo et al. 2009). Fmr1-KO mice treated with alpha-tocopherol had reversals of many of the classic findings of FXS, including macroorchidism, and behavior and learning deficits (de Diego-Otero et al. 2009). In a Phase II pilot RCT of ascorbic acid and alpha-tocopherol, adolescent males with FXS had behavioral and learning improvements, as well as reduced hyperactivity, mirroring the behavioral changes seen in mouse models (de Diego-Otero et al. 2014).
Muscarinic acetylcholine receptors
Another group of neurotransmitters affected by FMRP levels are the muscarinic acetylcholine receptors (mAChR), found not only in the brain but also throughout the central nervous system. Studies of the Fmr-1 KO mice have shown overactive M1 receptors, lending themselves as a potential treatment target in FXS (Veeraragavan et al. 2011). In a study of Fmr-1 KO mice being treated with dicyclomine, an M1 antagonist, mice were found to have fewer perseverative behaviors and fewer auditory-evoked seizures, without overall sedative effects (Veeraragavan et al. 2011). The authors concluded that pathogenesis of FXS may involve the cholinergic system. However, trihexyphenidyl, another M1 antagonist, was less beneficial in Fmr-1 mice than in the wild type, and motor stimulation was greatly increased (Fish et al. 2013). Similarly, donepezil, which acts to inhibit acetylcholinesterase and therefore prolongs the effects of acetylcholinergic neurons, failed to show increased IQ scores or improved behavior over a 12-week period in an RCT of 20 boys with FXS (Sahu et al. 2013).
Catecholamine stimulation
Based on the hypothesis that hyperactivity and reduced attention/cognitive function lead to impaired performance in Fmr-1 KO mice, Ventura et al. (2004) conducted a study of amphetamine (a drug that increases the release of dopamine and norepinephrine), which showed improved Fmr-1 KO mouse performance on cognitive tasks without effects on the motor activity levels of the mice. They concluded that prefrontal cortical dopamine plays a role in cognitive impairment in FXS.
Aripiprazole (D2, 5-HT 1A agonist; 5-HT 2A antagonist) is an atypical antipsychotic that is FDA approved for the treatment of irritability associated with autism spectrum disorders in the pediatric population. It acts on multiple neurotransmitter systems, as a partial agonist at the D2 and 5-HT 1A receptors, and an antagonist of 5-HT 2A receptors. In a Phase II open-label trial of aripiprazole in FXS, positive treatment response of lessened irritable behavior was seen in 10 of 12 individuals. Two individuals discontinued the study because of side effects or adverse events. To date, no placebo-controlled trials have been undertaken (Erickson et al. 2011a).
A retrospective chart review gives some evidence that early treatment with sertraline may have a positive effect on language development in children younger than 5 years (Indah Winarni et al. 2012). To the best of our knowledge, the only clinical trial of selective serotonin reuptake inhibitors (SSRIs) for the treatment of FXS is a Phase II controlled trial at the University of California, Davis, of sertraline as a modality to improve language development in young children (Greiss Hess et al. 2016). The study investigators evaluated the efficacy of low-dose sertraline in an RCT of 52 children aged 2 to 6 years. While there was no difference between sertraline and placebo on the primary outcome measures of the Mullen Scales of Early Learning expressive language age equivalent and the CGI-I, there was modest but nominal significant improvement shown on the analysis of some secondary measures (including social participation and motor and visual perceptual abilities). Sertraline was well tolerated. The authors concluded that these results warrant further studies of sertraline in young children with FXS.
Lovastatin
Although lovastatin is widely known for its competitive inhibition of cholesterol synthesis, it may also be used to regulate protein translation in the hippocampus. In Fmr-1 KO mice, lovastatin has been shown to prevent seizure onset by limiting excessive protein production (Osterweil et al. 2013). Based on the results from animal studies, there is currently interest in carrying out human trials in individuals with FXS, and one open-label study suggested possible benefit (Caku et al. 2014).
PAK inhibitors
Aberrant dendritic spine growth is a prominent neurocellular phenotype in FXS. Therefore, a logical target for pharmacological intervention is a protein regulator of spine proliferation, namely, p21-activated kinase or PAK. After only one administration of the PAK-inhibitor FRAX486, Dolan et al. (2013) were able to restore normal spine growth, eliminate seizure activities, and induce positive behavioral changes in adult Fmr-1 KO mice.
Minocycline
Minocycline has been shown to lower levels of MMP-9, previously known to be elevated in Fmr-1 KO mice (Bilousova et al. 2009). MMP-9 mRNA has been identified as an FMRP target (Dziembowska and Wlodarczyk 2012; Sidhu et al. 2014). Bilousova et al. (2006, 2009) reported that MMP-9 dysregulation may contribute to FXS-associated deficits. After mouse studies of the antibiotic minocycline showing improved behavior and fewer of the synaptic abnormalities associated with FXS, an open-label study was conducted in 20 individuals aged 13–32 with FXS and seemed to show some individual benefit (Paribello et al. 2010). As a result, an RCT was undertaken in 66 children and adolescents (aged 3.5 to 16 years)with FXS. Treatment for 3 months showed no serious side effects, and global improvement was seen; however, results may be confounded because of early unblinding of subjects, preliminary data analysis, and results being shared with investigators (Leigh et al. 2013). It also is unclear if the benefits of minocycline are due to its inhibitory effects on MMP-9 or its antibiotic properties (Sidhu et al. 2014).
NNZ-2566
An insulin-like growth factor-1 (IGF-1) analog, NNZ-2566, was tested by Deacon et al. (2015) in Fmr-1 KO mice based on the hypothesis that an intervention modulating neuroinflammation, as well as astrocyte and glial function, could rescue the abnormalities in the phosphoinositide 3-kinase–protein kinase B–mammalian target of rapamycin and ras GTPase–mitogen-activated protein kinase–signaling pathways in the Fmr-1 KO mouse. These mice showed improvements in behavioral features, such as activity level, learning, and memory, as well as physical phenotypes, including dendritic spine density and macroorchidism, potentially implicating the IGF-1 molecular pathway in the pathogenesis of FXS (Deacon et al. 2015). This has led to a Phase II, randomized, double-blind, placebo-controlled, parallel-group fixed-dose study of NNZ-2566 in human males aged 12–45 years with FXS (Neuren Pharmaceuticals Ltd.). This study has been completed, but the results are pending.
MG01CI/metadoxine
Metadoxine is a drug approved in many countries for the treatment of alcohol intoxication and withdrawal. The precise mechanism of action is unknown and is not related to the monoamine pathways (Manor et al. 2014). It has been studied to improve attention as it has cognitive enhancement characteristics and has demonstrated efficacy in a Phase IIb trial for adult ADHD (Manor et al. 2012). A Phase II RCT examining the effects of metadoxine for the treatment of inattention in FXS has been completed, but the results are pending. Previous studies have shown significant improvement in selective attention in adults with inattentive ADHD symptoms (Manor et al. 2014).
Oxytocin
Oxytocin has previously been shown to reduce anxiety, including social anxiety (Kirsch et al. 2005). Given that anxiety is a well-known problem in FXS, Hall et al. (2012) conducted a Phase II, double-blind, placebo-controlled single-dose trial in 10 individuals with FXS. Social anxiety, as measured by eye gaze frequency, improved significantly with oxytocin. The authors concluded that oxytocin may be helpful for social anxiety in FXS and recommended further placebo-controlled trials with oxytocin to be conducted in combination with behavioral treatment programs.
Epigallocatechin-3-gallate
Estrogens targeting estrogen receptor beta (ER-B) can be neuroprotective by inducing synaptic plasticity. Flavonoids, such as epigallocatechin-3-gallate (EGCG), interact with membrane endoplasmic reticulum. More commonly known as green tea extract, the estrogen receptor-beta target EGCG, a dietary supplement, may have protective effects on dendritic growth and plasticity. Animal models of FXS treated with EGCG showed improved recognition (de la Torre and Dierssen 2012). Therefore, a Phase II trial of EGCG for FXS has been undertaken, but the results are still pending (Parc de Salut Mar).
Conclusion
The review of the preclinical, Phase I, and Phase II trials of agents with therapeutic potential in FXS revolves around an understanding of the putative pathways in the pathogenesis of FXS. As seen in Figure 1, while there is significant overlap between some of these pathways, the agents can be categorized as mGluR antagonists, GABAergic agents, modulators of the mGluR-AMPA-GSK pathway, and miscellaneous modulators. Of these, most of the focus has been for the mGluR antagonists and the GABAergic agents, based on recent theories for etiopathogenesis of FXS holding the most promise (Bear et al. 2004). However, so far, clinical trials for these agents have not been able to demonstrate significant efficacy in treating FXS with the outcome measures utilized.
For the last decade, FXS has held the promise that disorders of intellect can be amenable to therapeutic interventions. As a result, there has been an explosion of research in opportune directions, with rapid translation into human studies. While unfortunately this has not led to a major clinically relevant breakthrough, critical insights as to the biology of neurodevelopmental disorders, the role of animal models, and the challenges of carrying out clinical trials on intellectually and behaviorally challenged individuals have been obtained.
Early studies focused on partial correction of perturbed neurosignaling, particularly at the AMPA receptor. The lack of efficacy raised the questions of drug specificity and potency in a system with many perturbed pathways. This heightened the excitement for more direct therapies, particularly the proof-of-concept Phase I trial of the mGluR5 inhibitor fenobam, as it was suggested that redressing the direct upstream biochemical loss of attenuation in the central mGluR5 pathway would have a cascading effect on downstream intracellular molecular targets and thus improve many of the consequent symptoms. Minocycline, as a weak indirect effector of the mGluR pathways, appeared to give some support to this hypothesis, in an open-label trial that showed a proportion of individuals seemed to benefit from treatment.
Around the same time, there was similar interest surrounding the controlled and blinded Phase II trial of arbaclofen. This drug did not aim to restore biochemical equilibrium, but instead to target the primary neurophysiological deficits in GABAergic activity that result and therefore to have downstream effects at the electrochemical level. The outcome measures were not able to confirm this to the necessary degree, but family and investigator interest in clinical trials remained high as the first Phase II trials of mGluR5 inhibitors, AFQ056 and R04917523, got under way.
While outcome data from these trials were also not compelling, data suggested that there was a basis to the theory and solidified the emerging realization that failure of these trials was due, in part, to the challenge of assessing outcomes in human subjects. This is important because interest in pursuing target molecules that interface with the mGluR pathway remains strong and pharmaceutical companies are actively involved in developing trials to pursue this opportunity. For example, AMO pharma has developed an inhibitor of the Ras-ERK pathway upstream of 6S kinase. In preclinical efficacy studies, AMO-01, the trial agent, has been shown to rescue the neuronal phenotype of the FMR1 knockout in transgenic mouse models of FXS. Another example is Ovid Therapeutics' drug gaboxadol, a GABAA receptor agonist, that appears to target the disruption of tonic inhibition involved in the regulation of excitatory and inhibitory signals that are aberrant in fragile X.
Less targeted trials, for example those of riluzole (Erickson et al. 2011b) and donepezil (Sahu et al. 2013), also suffered from a lack of efficacy; although in those trials, it was less clear if the failure was due to lack of efficacy or inadequate outcome measures.
The solidity of the biochemical models being assessed in each of these trials supports the ever-increasing call for improved methods of assessing outcome in humans. Unlike in the animal models, there is no readily ascertainable, objective standardized measure of drug efficacy in people. This shortfall, coupled with the phenotypic variability and symptomatic fluctuations of affected individuals, has further complicated the ability to describe meaningful changes to the most essential functional variables with current outcome measures.
The result is that these trials, while superficially disappointing, have allowed a maturation of the field of therapeutic trial assessment in the intellectually disabled population and have strengthened commitments to improving the assessment and outcome measures for these individuals. This is consistent with the goals of the NIMH's research domain criteria aim to refocus clinical research to better align cognitive and biological science with outcome measures that not only generate measurements but also probe the underlying mechanisms of the disorder. To do this, new and modified outcome measures need to be developed through a rigorous process to insure that they are clinically relevant to the population under study and should avoid fitting standardized nonspecific criteria.
Current trials have accordingly evolved with different outcome emphasis and an additional focus on concomitant outcome measure assessment, integrated into the protocol of a traditional drug trial. For example, a recent study used transcranial magnetic stimulation to assess cortical plasticity and excitability in individuals with autism and FXS (Oberman et al. 2010). Using new measures to examine previously assessed drugs affords the opportunity to accurately explore their merit, a strategy that is now being implemented. As Thomas Insel, former director of NIMH, stated “trials will test for target engagement itself as a potential mechanism of disease.
These studies, with either biomedical or psychosocial interventions, should prove informative even when the intervention does not prove efficacious because the goal of the trial is to learn about the disorder and not simply to demonstrate efficacy of the intervention” (Insel and Gogtay 2014). Examples that may be particularly pertinent to fragile X should comprise both neurophysiological measures and psychological assays that measure functional competency. These could include measures of appropriate sensory responsiveness, such as eye tracking, prepulse inhibition, and assays of stimulated evoked potentials such as transcranial magnetic stimulation, all of which have been noted to have differential responses in individuals with fragile X (Zingerevich et al. 2009; Oberman et al. 2010; Hall et al. 2015).
Similarly, and perhaps in parallel, psychological and other functional assays may need to be measured more naturalistically (Hall et al. 2015). Ideally, this would take place in the home or other familiar environmental settings, where test artifact has less influence and achievement of measurable outcomes can be more directly assessed through video or other modalities (O'Keefe et al. 2014; Kurtz et al. 2015). While biochemical biomarkers, such as ERK (extracellular signal-regulated kinase), offer some interesting possibilities, the challenge of confirming that plasma effects correlate with the brain remains an issue to resolve. Unsuccessful trials have also provided valuable lessons in working with affected individuals and their families, as disappointments have impacted the relationship between the community and professionals.
While the optimism of the early days in FXS drug development has been tempered, a new considered approach allows for a more systematic and stepwise development to therapeutic studies. Fragile X remains in the vanguard of treatable intellectual disabilities and continues to provide invaluable lessons that may be applied to a wide range of conditions related to intellectual disability and autism spectrum disorders.
Clinical Significance
There has recently been an explosion of research into drug development for FXS, as this disorder has held the promise that disorders of intellect can be amenable to therapeutic interventions. While this research has not yet resulted in any major breakthrough, our review of the preclinical and Phase I and Phase II clinical trials for FXS provides critical insights into the biology of neurodevelopmental disorders, which can help guide innovative clinicians in the use of pharmacological treatment agents for these conditions and their psychiatric comorbidities, while also providing researchers insights into the development of agents for this condition.
Footnotes
Disclosures
K.M.: No financial disclosures. Has been a Site Investigator for pharmaceutical trials sponsored by Johnson & Johnson, GSK, Novartis, and Alcobra. K.P.: None. J.G.-H.: In the past three years, J.G.-H. has received grant support from the Tommy Fuss Fund and the Al Rashed Family. He has equity in and is Founding Head of the Scientific Advisory Board for Neuro'motion, Inc., a company that is developing technology-based games to foster development of emotional regulation skills. In previous years, he has served as a consultant to Abbott Laboratories, Pfizer Inc, Johnson & Johnson (Janssen, McNeil Consumer Health), Novartis, Parke–Davis, GlaxoSmithKline, AstraZeneca, and Seaside Therapeutics; has been a speaker for Abbott Laboratories, Pfizer Inc, Novartis, Bristol-Meyers Squibb; and has received grant support from Abbott Laboratories, Pfizer Inc, Johnson & Johnson (Janssen, McNeil Consumer Health), GlaxoSmithKline, Akzo Nobel/Organon, and the NIMH. J.D.P.: Founder and Chief Scientific Officer with equity in Global Gene Corporation, a company focused on developing genomics data in the developing world. In the past three years, J.D.P. has been Local Co-PI on Novartis AFQ056 Phase II Trial for FXS; Local PI on Seaside Therapeutics Arbaclofen Phase II Trial in FXS; Member of Data Safety Monitoring Board for Roche RO4917523 Phase II Trial in patients with FXS.