
Abstract
Objectives:
Prior studies have shown that youth with bipolar disorder demonstrate neurofunctional changes in key prefrontal and subcortical brain regions implicated in emotional regulation following treatment with pharmacological agents. We recently reported a large response rate (>60%) to quetiapine (QUET) for treating depressive symptoms in adolescents with bipolar depression. This study investigates the neurofunctional effects of QUET using functional magnetic resonance imaging (fMRI).
Methods:
Thirty-three unmedicated subjects, 10–17 years of age, with a current depressive episode (Children's Depression Rating Scale-Revised [CDRS-R] > 40) associated with bipolar I or II disorder were recruited in a two-site randomized, placebo (PBO)-controlled trial of QUET monotherapy for treatment of bipolar depression in adolescents. Twenty-three of these participants (nine male) underwent an MRI scan at baseline, then were randomized to QUET or PBO, followed for 8 weeks, and at the end of their study participation underwent another MRI scan. During the fMRI scan, subjects viewed negative and neutral pictures and rated the valence of each picture.
Results:
Sixteen subjects had usable data at both time points: 10 subjects randomized to QUET, and 6 randomized to PBO. For QUET subjects, lower baseline activation in the left dorsolateral prefrontal cortex (p < 0.005) and higher baseline activation in the left ventrolateral prefrontal cortex (p = 0.0024) predicted greater improvement in CDRS-R scores from baseline to follow-up. When QUET and PBO groups were combined (n = 16), region-of-interest activation did not significantly predict change in CDRS-R.
Conclusions:
Baseline activation patterns in dorsal and ventral portions of the prefrontal cortex that are critical for the regulation of emotion-predicted response, but only within the QUET group. Thus, specific medications may be more effective in the context of specific prefrontal activation patterns in youth with bipolar depression. Larger studies of these youth would help to clarify the effects of QUET on brain activation.
Introduction
B
Quetiapine (QUET) has been found to be effective for treating bipolar depression in adults (Calabrese et al. 2005; Thase et al. 2006) and acute mania in youth (Findling et al. 2013; Pathak et al. 2013). We previously reported that QUET monotherapy was no different than placebo (PBO) in treating depressive symptoms in adolescents with bipolar depression (DelBello et al. 2009). Furthermore, in a larger study of 193 adolescents with bipolar depression, extended-release QUET was similarly no more effective than PBO (Findling et al. 2014). Nonetheless, because both studies observed a large response rate for adolescents taking QUET (>60%), and because of the potential for QUET to alter neurofunctional responses to negative stimuli (Fahim et al. 2005; Davis et al. 2015), this trial provided an opportunity to investigate changes in brain function associated with clinical improvement in depression associated with BD.
QUET is an atypical antipsychotic that is believed to exert its effects by blocking D2 dopamine and 5-HT2 serotonin receptors (Sanford and Keating 2012). However, its effects on brain activation, measured by changes in blood oxygenation level-dependent signal, have only been studied in adults with schizophrenia (Fahim et al. 2005; Nejad et al. 2013) and in adults with mania (Davis et al. 2015). These studies suggest that QUET treatment is associated with increases in prefrontal activation, particularly in dorsolateral and ventrolateral subregions of the prefrontal cortex (Jones et al. 2004; Stip et al. 2005; Meisenzahl et al. 2006; Davis et al. 2015). To our knowledge, no studies have directly examined the neurofunctional effects of QUET in pediatric populations or in any patients with depression associated with BD.
Several studies in youth with BD have now demonstrated that prefrontal-limbic neural circuitry, important for emotional regulation, may be highly responsive to a variety of interventions (Mayanil et al. 2011). We reported that youth with bipolar depression treated with lamotrigine had less amygdalar activation while viewing negative stimuli as depressive symptoms improved (Chang et al. 2008). Indeed, youth with BD across mood states have demonstrated a consistent pattern of increased functional activity in ventrolateral and medial prefrontal regions with lamotrigine (Pavuluri et al. 2010; Passarotti et al. 2011), divalproex (Pavuluri et al. 2012a, 2012b), and risperidone treatment (Pavuluri et al. 2011). We have also examined the neural correlates of clinical improvement in youths at high risk for BD, and have found that dorsolateral prefrontal cortex (DLPFC) activation increases as symptoms improve from a family-focused intervention (Garrett et al. 2015). Altogether, these studies suggest that the prefrontal cortex may be a promising target for treatment in pediatric bipolar depression.
We now report on functional magnetic resonance imaging (fMRI) findings in adolescents treated with QUET or PBO for 8 weeks. We hypothesized that response of depressive symptoms in all subjects would be associated with relative increases in prefrontal activation, and that relative deficits in prefrontal activation would predict response to QUET.
Methods
Study design
Children and adolescents (12–18 years of age) with a current major depressive episode associated with bipolar I or II disorder were recruited from outpatient clinics at Stanford University and inpatient units and outpatient clinics at the University of Cincinnati (UC) to participate in this 8-week study. To be included in this study, potential subjects had a screening and baseline CDRS-R score >40 and Young Mania Rating Scale (YMRS) score <20, and were not taking antidepressants, mood stabilizers, or antipsychotics for at least 72 hours before baseline assessments. Additionally, the use of concomitant psychotropic medications was prohibited during study participation. After informed assent and consent were obtained from the subject and their legal guardian, respectively, subjects were randomized to QUET or PBO, which was titrated to 300–600 mg/d over 3–6 days. Randomization was stratified by gender and the presence of psychosis.
Assessments and analyses
The primary outcome measure, defined a priori, was change from baseline to endpoint in CDRS-R score. Secondary outcome measures were change from baseline to endpoint in Hamilton Anxiety Rating Scale (HAM-A), YMRS, Clinical Global Impression-Severity (CGI-S), and laboratory and safety measures, including extrapyramidal symptom ratings. Response (defined by a ≥50% improvement from baseline in CDRS-R) and remission (defined by CDRS-R ≤28 and Clinical Global Impression-Improvement [CGI-I] ≤2) rates in each group were calculated. Study visits with efficacy and safety ratings were performed at screening, baseline, and weekly, thereafter for 8 weeks. All patients, caregivers, and investigational staff who performed efficacy and tolerability ratings were blinded to patient treatment group. For details about the clinical trial, please see DelBello et al. (2009)
Neuroimaging subsample
Twenty-three subjects participated in the neuroimaging aspect of the study, consisting of baseline and posttreatment follow-up MRI scans after on average 57 days. Of these participants, 9 were scanned at Stanford University and 14 at UC. During both magnetic resonance imaging sessions, the same fMRI task was administered. Six subjects were excluded due to excessive motion and one for poor image acquisition. Thus, 10 subjects (5 girls) on an active dose of QUET were included in the final analysis. Six of the 10 subjects were scanned at UC. The post hoc exploratory analysis included an additional 6 PBO subjects (4 scanned at UC) contributing to a total of 16 subjects.
fMRI acquisition
At Stanford University's Lucas Center, images were acquired on a 3T GE Signa scanner using a custom-built head coil. Functional magnetic resonance images were collected using a spiral pulse sequence with the following parameters: repetition time (TR) = 2000 mseconds, excitation time (TE) = 30 mseconds, flip angle = 80°, field of view (FOV) = 220.0 mm, 30 slices (4 mm thick, 0.1 mm skip) along the AC-PC axis, 64 × 64 matrix, and 1 interleave. To reduce field inhomogeneities, an automated high-order shim was applied (Kim et al. 2002). At UC College of Medicine's Center for Imaging Research (CIR) images were acquired on a 4.0 Tesla Varian Unity INOVA Whole-Body MRI/MRS system (Varian, Inc., Palo Alto, CA) using a TEM (Transverse ElectroMagnetic) 1H headcoil. Functional magnetic resonance images used a T2-weighted gradient-echo echoplanar imaging pulse sequence with the following parameters: TR = 2000 mseconds, TE = 30 mseconds, flip angle = 75, FOV = 220.0 mm, 30 slices (4 mm thick, 0.1 mm skip), 64 × 64 matrix.
Affective pictures task
Negative (e.g., a mutilated dog), positive (e.g., puppies), and neutral (e.g., a plate) pictures that were deemed appropriate for a pediatric population were selected from the International Affective Picture System (IAPS) (Lang et al. 2005). The three types of pictures were presented in blocks, each block containing six pictures, with each picture presented for 4500 mseconds with a 500 mseconds interstimulus interval. Subjects were asked to indicate how each picture made them feel by pressing one of three buttons corresponding to “negatively,” “neutrally,” and “positively.” Eprime software was used to present images, and a hand-held button box was used to collect responses (Psychology Software Tools, Inc.). At Stanford University, stimuli were projected to the subjects from the foot of the scanner onto a projection screen attached to the head coil. Subjects viewed the stimuli by looking directly up at a mirror that reflected the projection screen. At the UC, stimuli were presented to the subject using nonferromagnetic goggles. At Stanford, responses and response times were collected by Eprime. Although subjects at UC were observed to competently respond to stimuli presented during fMRI, operator error resulted in no recordings of behavioral responses to the task. Therefore, all IAPS task performance statistics were calculated from data collected at Stanford University.
Image preprocessing and statistical analysis
Functional data were preprocessed and statistically analyzed using SPM5 (
fMRI regions of interest analysis
The following spherical regions of interest (ROIs) were placed, by a trained research assistant, on the peak loci of activation from a previous study in which pediatric subjects with BD demonstrated greater activation compared with healthy controls when performing the same task (Chang et al. 2004). ROIs included 7 mm spherical ROIs in the left DLPFC (MNI coordinates: −48, 12, 28) and right DLPFC (MNI coordinates: 48, 16, 22), and a 5 mm spherical ROI in the left ventrolateral prefrontal cortex (VLPFC; MNI coordinates: −32, 20, 12). Based on the finding of abnormal metabolite concentrations in the anterior cingulate cortex (ACC) of adolescents with BD (Patel et al. 2008; Singh et al. 2010), an additional spherical ROI with a 8 mm radius was placed in the pregenual ACC (MNI coordinates: 0, 47, 0). ROI radii were determined to maximize gray matter coverage while not extending beyond the boundaries of the region. MarsBar (
Correlations between fMRI and clinical symptom scores
Independent t-tests were used to compare demographic and clinical characteristics of the QUET and PBO groups. Linear regression was used to determine whether baseline activation within the selected ROIs could predict treatment response, (change in CDRS score). Change in activation was correlated with change in CDRS score to assess the neural correlates of symptom improvement. All analyses controlled for site (Stanford or Cincinnati) and baseline CDRS scores.
Results
Clinical measures
Of the 23 subjects who were scanned, 16 had usable baseline imaging data. Seven subjects were excluded due to either poor image acquisition or rapid motion (>0.5 mm/TR) exceeding 25% of the task collected during the baseline. Of the 16 participants with usable scan data, 10 received QUET (5 female, 6 scanned at Cincinnati and 4 scanned at Stanford). QUET subjects had mean age of 16.1 ± 2.02 years and mean CDRS scores of 54.4 ± 5.9 and 31.4 ± 11.6 at baseline and follow-up, respectively. PBO subjects had mean age of 15.33 ± 1.75 years and mean CDRS scores of 57.8 ± 7.8 and 23.4 ± 5.8 at baseline and follow-up, respectively. Six of the subjects received PBO (four female, four Cincinnati and two Stanford). Seven of 10 subjects receiving QUET and 4 of 6 receiving PBO were considered responders by CDRS-R criteria. Thus, there were a total of 11 responders and 5 nonresponders. Three in the PBO group and five in the QUET group were considered remitters.
Affective pictures task performance
Ratings for negative, neutral, and positive pictures were averaged and compared across conditions. As expected, there was a significant effect of valence, indicating that all subjects rated the positive, negative, and neutral pictures significantly differently (baseline, F = 36.7, p < 0.001; follow up, F = 69.5, p < 0.001). Ratings did not change from baseline to follow-up [paired t-test, two-tailed; negative pictures, t(5) = −0.22, p = 0.84; neutral, t(5) = 0.40, p = 0.71; positive, t(5) = −0.57, p = 0.59]. Similarly, average reaction times within blocks of the same valence did not change from baseline to follow-up [paired t-test: negative pictures: t(5) = 0.78, p = 0.47; neutral, t(5) = 0.41, p = 0.70; positive, t(5) = 0.62, p = 0.57].
Using the negative pictures to neutral picture contrast, within the QUET group at baseline, a significant correlation was found between higher CDRS scores and lower left DLPFC activation, r = −.775, p = 0.008. Baseline activation in the left VLPFC, right DLPFC, and ACC ROIs were not significantly correlated with baseline CDRS scores. Furthermore, at follow-up, ROI activations were not correlated with CDRS scores.
However, using a linear regression, baseline activation in the left DLPFC and left VLPFC significantly predicted change in CDRS from baseline to follow-up (Table 1) in the QUET group. Additionally, lower baseline left DLPFC and higher baseline left VLPFC predicted greater CDRS reductions from baseline to follow-up (Figs. 1 and 2). Right DLPFC and ACC ROIs did not significantly predict changes in CDRS.
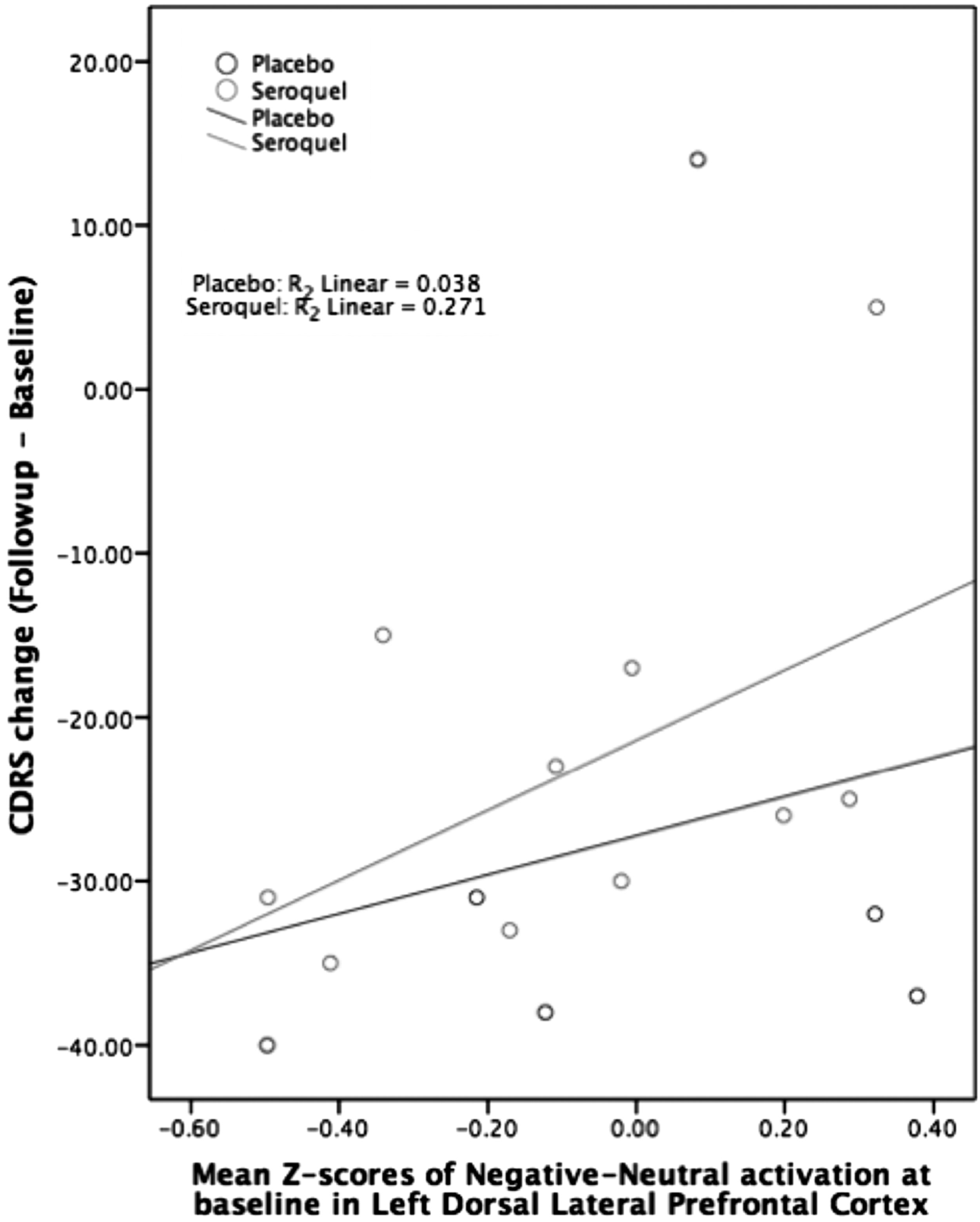
Left DLPFC activation plotted with CDRS change. CDRS, Children's Depression Rating Scale; DLPFC, dorsolateral prefrontal cortex.

Left VLPFC activation plotted with CDRS change. VLPFC, ventrolateral prefrontal cortex.
ACC, anterior cingulate cortex; CDRS, Children's Depression Rating Scale; LDLPFC, left dorsolateral prefrontal cortex; LVLPFC, left ventrolateral prefrontal cortex.
When the PBO and QUET groups were combined, baseline ROI activation was not significantly associated with change in CDRS. CDRS change plotted with left VLPFC and left DLPFC z-scores did not show an association between QUET and PBO groups (Figs. 1 and 2).
Across all subjects from both treatment conditions who had a decrease in CDRS scores (n = 14) from baseline to follow-up, the left VLPFC baseline activation was significantly associated with the left DLPFC baseline activation (Table 2, Fig. 3). With the addition of two subjects (n = 16) who had increased CDRS scores from baseline to follow-up the association between left VLPFC and left DLPFC was no longer significant.
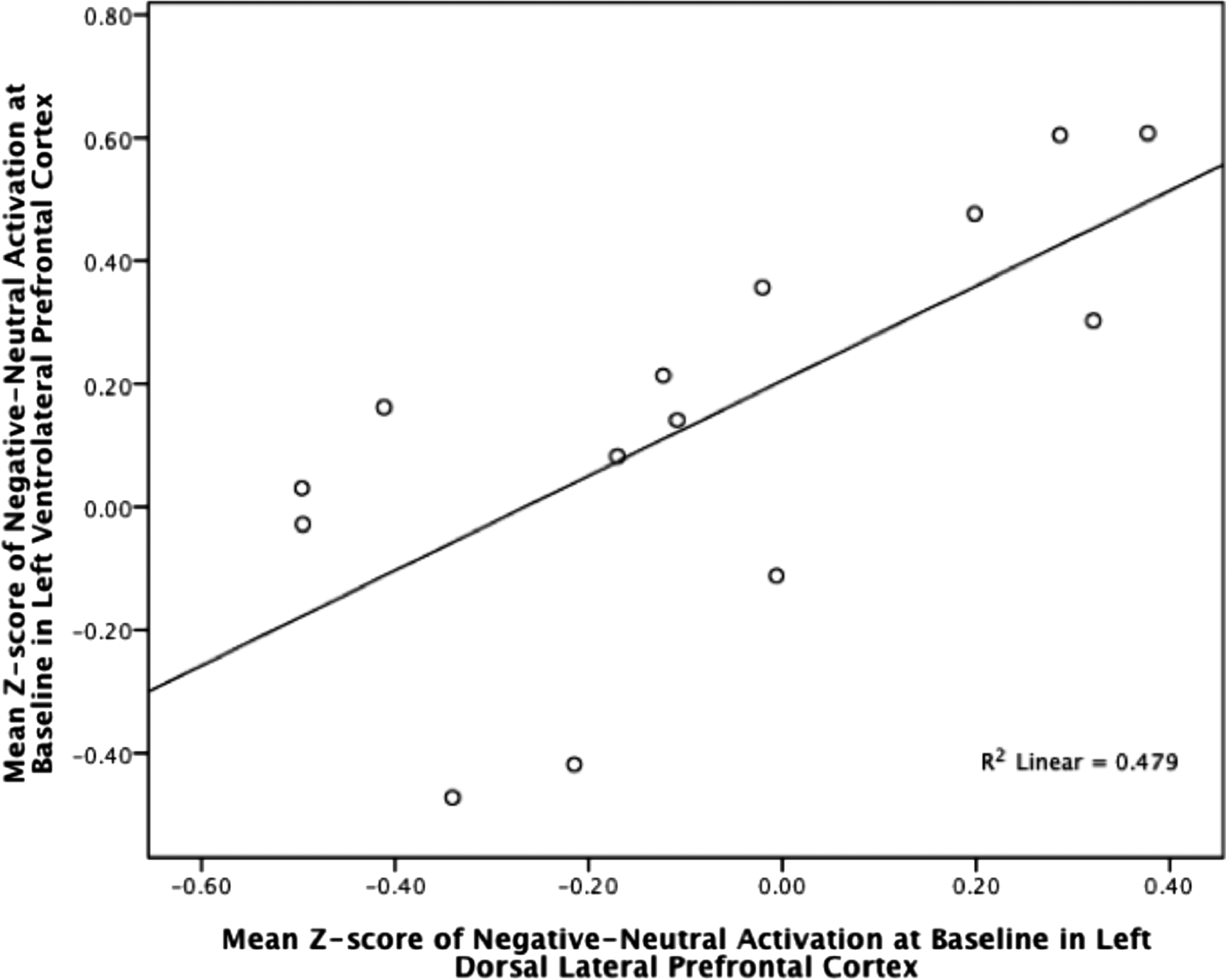
Left VLPFC activation plotted with left DLPFC activation across subjects with reduced CDRS scores. DLPFC, dorsolateral prefrontal cortex.
ACC, anterior cingulate cortex; LDLPFC, left dorsolateral prefrontal cortex.
Change in activation was not significantly correlated with change in CDRS for any ROI within the QUET group, across all subjects.
Discussion
This is the first study using fMRI to investigate the neural correlates of response to QUET in youth with bipolar depression. Contrary to our predictions, we did not find that increases in prefrontal activation were correlated with response to QUET treatment. However, decreased baseline activation of left DLPFC and increased baseline activation of left VLPFC when performing an affective task predicted greater response in youth treated with QUET. In subjects who responded positively to either QUET or PBO, these two regions were inversely correlated in their activation. Thus, lower DLPFC and higher VLPFC activation at baseline during an emotion task may indicate a failure to engage higher cognitive control centers critical for emotional regulation. Potentially less efficient prefrontal ventral structures (VLPFC) may then be relatively overactivated instead of DLPFC, which mirrors the increase in functional connectivity between amygdala and VLPFC reported in youth with BD (Hafeman et al. 2017). Importantly, our study suggests that these same regions may be associated with depressive states that are more responsive to treatment.
Our results are consistent with functional neuroimaging studies in adults with bipolar depression that suggest core impairments in brain regions critical for emotional regulation. fMRI studies in adults with bipolar depression have reported reduced activations in dorsal and ventral portions of the prefrontal cortex (Altshuler et al. 2008; Townsend et al. 2010; Liu et al. 2012), both limbic hypoactivity (Vizueta et al. 2012) and hyperactivity (Perlman et al. 2012), and aberrant connectivities between prefrontal-limbic circuits (Versace et al. 2010; Cerullo et al. 2012; Vizueta et al. 2012). Importantly, prefrontal hypoactivity inversely correlates with depression symptom severity, and individuals with bipolar depression can simultaneously show both hypoactivity in the DLPFC and failure to deactivate the medial prefrontal cortex, suggesting important functional distinctions in prefrontal subregions during emotional processing (Fernández-Corcuera et al. 2013). These studies provide a neural signature for bipolar depression, emphasizing the importance of considering mood state in the neural evaluation of individuals with BD. Specifically, researchers have demonstrated through longitudinal observations of individuals with BD that the transition from a manic state to a depressive state can lead to a change in amgydala connectivity shifting away from the VLPFC to the insula (Cerullo et al. 2012). This finding suggests that unique markers of mood state may be found within individuals with BD across manic and depressive episodes.
Most fMRI studies in pediatric BD were designed without explicit consideration of mood state, except in post hoc analyses (Rich et al. 2008; Singh et al. 2010). Two recent fMRI studies were reported in youth with bipolar depression, who were treated naturalistically with multiple pharmacological interventions and combinations (Diler et al. 2013a, 2013b). Youth in these studies were scanned at baseline and then again 6 weeks after naturalistic treatment and were presented with emotion processing (Diler et al. 2013a) and response inhibition (Diler et al. 2013b) fMRI tasks. Ten adolescents were studied and had an average of 57% improvement in depressive symptoms over the course of 6 weeks. During emotion processing, these youth demonstrated increased right VLPFC, insula, cerebellar activations that corresponded with reduced negative emotion processing, and increased positive emotion processing after treatment. Furthermore, higher activity in ventral ACC to mild happy faces found at baseline was associated with greater improvement in depression. During response inhibition, these youth demonstrated baseline hyperactivity in the VLPFC that was not responsive to treatment. Instead, these youth had increased activity in the hippocampus and thalamus after treatment. Lower left thalamus activity at baseline was correlated with higher depression scores. These latter findings are contrary to our findings, but may be due to differences related to task demand and nonspecificity of treatment targets.
We should note a number of limitations of this study. First, the cohort of subjects with useable MRI data was small in sample size with fewer subjects randomized to PBO. These small cell sizes limited our ability to evaluate whether neural effects of QUET could be distinguished from those found in youth randomized to PBO, or if the PBO was working through an alternative mechanism. Second, to increase power, we combined fMRI data collected from two different scanners at two different sites. However, covarying for scanner did not significantly change our results. Moreover, we conducted a traveling subjects study and found no between-scanner differences in fMRI activation using the same task used in the study. Future studies with larger number of subjects are needed to clarify the effects of QUET in this patient population.
Conclusions and Clinical Significance
In summary, our preliminary data suggest that youth with bipolar depression may show alterations in baseline prefrontal neural function. These dorsal and ventral prefrontal subregions may be promising candidates for biological markers of treatment response in bipolar depression. In this study, we found preliminary evidence that youth with bipolar depression who have higher severity of depressive symptoms have lower prefrontal activation that may be more predisposed to neurofunctionally respond to quetiapine than youth who have lower severity of depressive symptoms and higher prefrontal activation. Understanding how these brain regions respond to different treatments may eventually help guide more personalized treatments for youth with mood disorders. Larger studies would further help to clarify the effects of quetiapine on brain activation in youth with bipolar depression.
Footnotes
Disclosures
Dr. Chang is a consultant for Sunovion. In the past 3 years he has been a consultant for Octapharma, GlaxoSmithKline, Actavis, and Janssen. Dr. Delbello has received Research Support from Otsuka, Lundbeck, Purdue, Sunovion, Pfizer, Johnson and Johnson, Supernus, Amarex, and AssureRx, and is on the Consulting/Advisory Board/or has received Honoraria from Pfizer, Lundbeck, Sunovion, Supernus, Takeda, Johnson and Johnson, Neuronetics, and Akili. Dr. Adler is a consultant for Neurocrine Biosciences and receives research grants from Merck and Forest. Dr. Strakowski is a consultant for Sunovion. Dr. Singh receives research support from Neuronetics and Johnson and Johnson. The other authors have no disclosures.