
Abstract
Objectives:
Environmental factors may interact with genetic factors via the epigenetic process, and this interaction can contribute to inter-individual variability in the treatment response. The purpose of this study was to investigate the interaction effects between dopamine receptor D4 (DRD4) methylation and prenatal maternal stress on the methylphenidate (MPH) response of youth with attention-deficit/hyperactivity disorder (ADHD).
Methods:
This study was an 8-week open-label trial of MPH that included 74 ADHD youth. We investigated the associations between MPH treatment response, which was defined as a score ≤2 on the Clinical Global Impressions-Improvement (CGI-I) scale, and the methylation of 28 cytosine-guanine dinucleotide (CpG) sites of DRD4. Additionally, the interaction effects between DRD4 methylation and prenatal maternal stress on changes in Continuous Performance Test (CPT) scores after MPH treatment were investigated.
Results:
Although there were no significant sites that showed significant association with treatment response, there was a significant interaction effect of the methylation of CpG7 and prenatal maternal stress on changes in omission errors of the CPT following treatment (p = 0.0001).
Conclusions:
The present findings indicate that the interaction between methylation of CpG7 of DRD4 and prenatal maternal stress may be predictive of the treatment response to MPH in youth with ADHD.
Introduction
A
Methylphenidate (MPH) is a first-line treatment for ADHD, and 65%–75% of patients treated with this drug show a response (Santosh and Taylor 2000); the reported effect size is between 0.54 and 0.78 (Schachter et al. 2001). The major molecular mechanism of MPH involves the blockade of dopamine transporters, which subsequently increases synaptic dopamine concentration and dopaminergic transmission (Wilens 2008). Another important factor related to the efficacy of dopaminergic transmission is the sensitivity of dopamine receptors (Seeger et al. 2001). Of the various dopamine receptor-related genes, the dopamine receptor D4 (DRD4) exon III 48-base-pair variable number tandem repeats (VNTR) polymorphism has frequently been the candidate gene in pharmacogenetic studies (Hamarman et al. 2004; Kieling et al. 2010).
The 7-repeat (7R) allele of this gene variant has been implicated in the MPH response in Caucasians (Froehlich et al. 2011), but this allele is rare in Asian populations, including Koreans (Lichter et al. 1993); thus, genetic research investigating DRD4 has tended to focus on the 4R allele. However, one study reported a significant association between the 4R allele and the MPH treatment response, whereas another reported negative results (Cheon et al. 2007; Kim et al. 2009). Another study found significant interaction effects between the DRD4 VNTR genoytpes and those of noradrenergic gene polymorphisms (Hong et al. 2012). The overall findings regarding the predictive value of DRD4 variants (4R and 7R) for the MPH response have been inconsistent (Contini et al. 2012; Soo et al. 2013), and this may be due to variability among the environmental factors that interact with genetic makeup (Domschke et al. 2014).
Several studies have shown that prenatal maternal stress is an important environmental risk factor for ADHD (Van den Bergh and Marcoen 2004; Rodriguez and Bohlin 2005). Animal studies have found that prenatal stress increases D2-like receptors in limbic areas and/or stimulates the release of dopamine in the nucleus accumbens while decreasing its release in cortical areas (Berger et al. 2002; Baier et al. 2012). Furthermore, human studies have reported positive interaction effects between maternal stress during pregnancy and the 7R allele of DRD4 on ADHD symptomatology, including externalizing behavior and aggression levels (Grizenko et al. 2012; Buchmann et al. 2014; Zohsel et al. 2014). However, the effects of the interaction between prenatal maternal stress and DRD4 on the MPH response have yet to be investigated.
DNA methylation, which is one of the most frequently studied epigenetic markers, can mediate gene–environment interaction effects on the treatment response in patients with psychiatric disorders (Elia et al. 2012). Methylation typically occurs at cytosine-guanine dinucleotide (CpG) sites and acts to repress gene activity (Goll and Bestor 2005). The utility of measuring methylation profiles when predicting the treatment response has been evaluated in patients with a variety of psychiatric disorders, including depression (Menke et al. 2012; Frieling and Tadic 2013), schizophrenia (Tang et al. 2014), and bipolar disorder (Houtepen et al. 2016).
Only one study has investigated the association between methylation and the MPH treatment response in ADHD. Ding et al. (2017) reported that methylation of the dopamine transporter 1 gene (DAT1) is negatively correlated with improvements in ADHD symptoms after 6 weeks of MPH treatment but that the treatment response did not differ according to one's DRD4 methylation status. However, a major limitation of this study was that it assessed the mean percentage of methylation across the CpG island rather than methylation at each individual CpG site. Moreover, this study did not consider the interaction between DNA methylation and environmental factors. Because various environmental risk factors for ADHD cause changes in DNA methylation, this process should be taken into account along with environmental risk factors when conducting a genetic study (Han et al. 2015).
Thus, the primary goals of this study were as follows: (1) to investigate the associations between the methylation of CpG sites in a promoter region of DRD4 and MPH treatment response; (2) to evaluate the effects of the interaction of DRD4 methylation sites and prenatal maternal stress on the MPH response; and (3) to evaluate the effects of the interaction of the DRD4 4R allele genotype and prenatal maternal stress on the MPH response. Because the methylation of DRD4 affects the extent of gene expression, it was hypothesized that the MPH responses of ADHD patients would differ according to the interaction of DRD4 methylation and the level of prenatal stress but not according to the interaction between the DRD4 4R genotype and level of prenatal maternal stress.
Materials and Methods
Participants
This study included 83 children and adolescents with ADHD (mean age: 9.5 ± 2.6 years, 65 boys [78.3%]) who were recruited between August 2010 and February 2013 from the Seoul National University Hospital Child and Adolescent Psychiatry outpatient clinic. A detailed explanation of the protocol has been previously reported (Hong et al. 2015; Kim et al. 2015).
All ADHD participants in the present study were medication-naïve and their ADHD diagnoses were confirmed according to the criteria of the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition (DSM-IV), by board-certified child and adolescent psychiatrists using the Kiddie Schedule for Affective Disorders and Schizophrenia-Present and Lifetime version (K-SADS-PL) (Kaufman et al. 1997; Kim et al. 2004). Potential participants were excluded if they had an IQ <70, a past or ongoing history of tic disorder, obsessive compulsive disorder, language disorder, learning disorder, convulsive disorder, pervasive developmental disorder, schizophrenia, bipolar disorder, or brain damage; a history of taking stimulants or atomoxetine for longer than 6 months; and/or a recent history (within the past 4 weeks) of taking stimulants or atomoxetine.
IQ was measured using the abbreviated form of the Korean Educational Developmental Institute's Wechsler Intelligence Scale for Children (Park et al. 1996). The presence of prenatal maternal stress was determined by asking mothers of the participants whether they experienced severe stress during pregnancy or not (Park et al. 2014).
Written informed consent was obtained from the adolescent participants (verbal assent in the case of child participants) and their parents before enrollment after sufficient explanation of the study. This study protocol was approved by the Institutional Review Board of Seoul National University Hospital.
Determination of DNA methylation
Genomic DNA was extracted from whole blood using an Intron_G-DEX™ IIb Genomic DNA Extraction kit (Intron, South Korea). A bisulfite sequencing procedure was performed to determine the CpG methylation profiles located upstream of the DRD4 coding regions; the experiment was carried out as previously described (Xu et al. 2015) with some modifications.
The bisulfite conversion was performed with the EpiMark Biosulfite conversion kit (New England Biolabs, Inc., Ipswich, MA) according to the manufacturer's instructions. The bisulfite conversion reaction was performed in a volume of 140 μL that contained 1 μg of genomic DNA isolated from blood and 130 μL of bisulfite mix, which was prepared by adding 650 μL of nuclease-free water and 250 μL of Solubilization Buffer. The steps of the bisulfite conversion reaction consisted of denaturation at 95°C for 5 minutes, incubation at 65°C for 30 minutes, denaturation at 95°C for 5 minutes, incubation at 65°C for 60 minutes, denaturation at 95°C for 5 minutes, incubation at 65°C for 30 minutes, and final incubation at 18°C–20°C for up to 12 hours.
Following the completion of the conversion reaction, desulfonation reaction and sample clean-up procedures were carried out using the EpiMark spin column provided with the conversion kit. First, the entire sample was loaded onto the EpiMark spin column and washed with 500 μL of Wash Buffer. Next, the desulfonation reaction was performed by adding 500 μL of desulfonation reaction buffer to the column and incubating it at room temperature for 15 minutes. After the incubation, the column was washed with 500 μL of Wash Buffer and the samples were eluted into 20 μL of Elution Buffer.
The bisulfite-treated DNA was then subjected to a polymerase chain reaction (PCR) procedure in which the nested PCR method was utilized to amplify the CpG island regions of DRD4. The following primers were used: for the first PCR of the DRD4 gene, primers DRD4_F1 (TAGGTTATTTTTTTTTGGTGAAGA) and DRD4_R1 (TCACCCTAATCCACCTAATATCT); and for the second PCR of the DRD4 gene, primers DRD4_F2 (TTGTTTAGGGTTAGAGGGG) and DRD4_R2 (ATCCACCTAATATCTAACAAAACC).
The first-step PCR was performed in a 25-μL volume containing 20–100 ng of bisulfite-treated DNA, 1 × PCR buffer, 0.2 mM of each deoxynucleotide triphosphate (dNTP), 0.2 μM of each primer for the first PCR, and 0.625 U of EpiMark Taq polymerase (New England Biolabs, Inc.). The reaction steps consisted of denaturation at 95°C for 30 seconds, followed by 40 cycles of 95°C for 30 seconds, 55°C for 1 minutes, and 68°C for 1 minutes, and then final extension at 68°C for 5 minutes. The second-step PCR was carried out using 1 μL of a 1:10 dilution of the first-step PCR product in a total volume of 25 μL under the same conditions as the first-step procedure but using the second PCR primers and an annealing temperature of 50°C.
Following the PCR procedure, any unincorporated primers and dNTPs were removed by adding a 1/10 volume of Exo-Sap (ExoProStar 1; GE Healthcare, Little Chalfont, United Kingdom), incubating it for 15 minutes at 37°C, and then incubating it for 15 minutes at 85°C for enzyme inactivation.
The resulting PCR products were directly subjected to DNA sequencing and the methylation status of each CpG island was manually called by visual inspection. The percentage of methylation was calculated as the peak height of C versus the peak height of C plus the peak height of T for each CpG site as shown in the sequencing chromatogram. Sites between 0% and 10% were classified as unmethylated (0%, U/U), sites between 90% and 100% were classified as fully methylated (100%, M/M), and sites between 10% and 90% were classified as hemimethylated (50%, U/M).
MPH administration and treatment response
This study was an 8-week open-label trial of MPH during which the participants visited the clinic at baseline and weeks 2, 4, 6, and 8. The doses of MPH were adjusted until sufficient treatment effects were achieved and then that dose was maintained for the remainder of the trial period; all final doses were set within the range of 0.7–1.5 mg/kg.
The primary treatment outcome in this study was the Clinical Global Impressions-Improvement (CGI-I) scale score at 8 weeks of MPH treatment; this scale was administered by board-certified child psychiatrists (Guy 1976). The CGI-I is a widely used clinician-based rating scale that assesses symptom improvement and evaluates changes in symptom severity compared to the baseline Clinical Global Impressions-Severity (CGI-S) score; scores are rated on a scale ranging from 1 (very much improved) to 7 (very much worse) (Guy 1976). All raters were blind to the methylation status of each subject; subjects with a CGI-I score ≤2 were classified as good responders and those with scores of 3–7 were classified as poor responders.
Before and after 8 weeks of MPH treatment, the Continuous Performance Test (CPT) was administered to assess behavioral changes following MPH administration (Shin et al. 2000). A computerized version of the CPT that utilizes visual stimuli was administered to the participants to measure impulsivity and inattention (Shin et al. 2000). The level of performance for each participant is represented using three variables: (1) omission errors (failures to respond), as a measure of inattention; (2) commission errors (false responses), as a measure of impulsivity; and (3) response time variability (the standard deviation of the response times of correct responses), as a measure of sustained attention. All data were automatically transformed into T-scores adjusted for age relative to a normal population of 847 children between 5 and 15 years of age (Shin et al. 2000); lower T-scores indicate better performance.
Statistical analysis
The demographic and clinical characteristics of the participants were compared between the good responder and poor responder groups using independent t-tests for continuous variables and chi-square or Fisher's exact tests for categorical variables. Due to the rarity of the M/M group at some sites, the U/M and M/M groups were combined into a single methylated group for the methylation analyses (Copoglu et al. 2016). Thus, the methylation status of each CpG site was categorized into two groups: unmethylated (U/U) and methylated (U/M+M/M). Changes in CPT scores following MPH treatment were compared between the methylation groups of each CpG site using independent t-tests or Mann–Whitney U tests.
The interaction effects between the methylation groups of each CpG site and prenatal maternal stress (positive or negative) on the treatment response, as defined by the CGI-I, were assessed using hierarchical multivariable logistic regression models, with methylation group and prenatal maternal stress included in the first block and the interaction between methylation and prenatal maternal stress included in the second block.
The interaction effects between the methylation of each CpG site and prenatal maternal stress (positive or negative) on score changes on the CPT were evaluated using hierarchical multivariable linear regression models, with methylation group and prenatal maternal stress included in the first block and the interaction between methylation and prenatal maternal stress included in the second block. For the CpG sites that exhibited a significant interaction, post hoc analyses were performed to determine the effects of methylation on treatment response or change in CPT score in the stress-positive and stress-negative groups independently.
All of the same analyses were conducted for the DRD4 4R allele for purposes of comparison. All statistical analyses were performed with SPSS ver. 22.0 software (SPSS, Inc.; Chicago, IL) and two-tailed p-values <0.0002 (0.05/28 CpG sites × 4 variables × 2 [for interaction]) were considered to indicate statistical significance.
Results
Of the initial 83 participants included in this study, 5 dropped out and 78 completed the 8-week trial; methylation data were available for 74 of these 78 participants. The demographic and clinical characteristics of the participants according to treatment response (45 good responders and 29 poor responders) are presented in Table 1. There were no significant differences between the two groups except that there were significantly more participants with comorbid oppositional defiant disorder in the poor responder group. The allele and genotype distibutions of the DRD4 polymorphism are presented in Supplementary Table S1 (Supplementary Data are available online at
ADHD, attention-deficit/hyperactivity disorder; CGI-S, Clinical Global Impressions-Severity; CPT, Continuous Performance Test; MPH, methylphenidate; NOS, not otherwise specified; ODD, oppositional defiant disorder; SD, standard deviation.
Table 3 presents the associations between treatment response, defined by the CGI-I, and methylation groups at each CpG site. There was no methylation site that showed a significant association with treatment response. Data for the pre- and post-treatment CPT scores were available for 69 patients. There were no differences in CPT scores following MPH treatment according to the methylation status of any CpG site.
Adjusted for age, gender, IQ, baseline CGI-S score, baseline ADHD-RS total score, final MPH dose.
ADHD-RS, Attention-Deficit/Hyperactivity Disorder Rating Scale-IV; AOR, adjusted odds ratio; CI, confidence interval; CGI-S, Clinical Global Impressions-Severity; MPH, methylphenidate.
There was a significant interaction effect between methylation at CpG7 and prenatal maternal stress level in terms of changes in omission errors on the CPT following MPH treatment (p = 1.6 × 10−4; Table 4). The post hoc power analyses revealed a power of 99.0%. The post hoc analyses revealed no difference in treatment response according to the methylation status of CpG7 in the stress-positive group; however, in the stress-negative group, the unmethylated group exhibited a nominally significant larger change in omission errors in the post-treatment testing than did the methylated group (p = 0.001; Fig. 1).
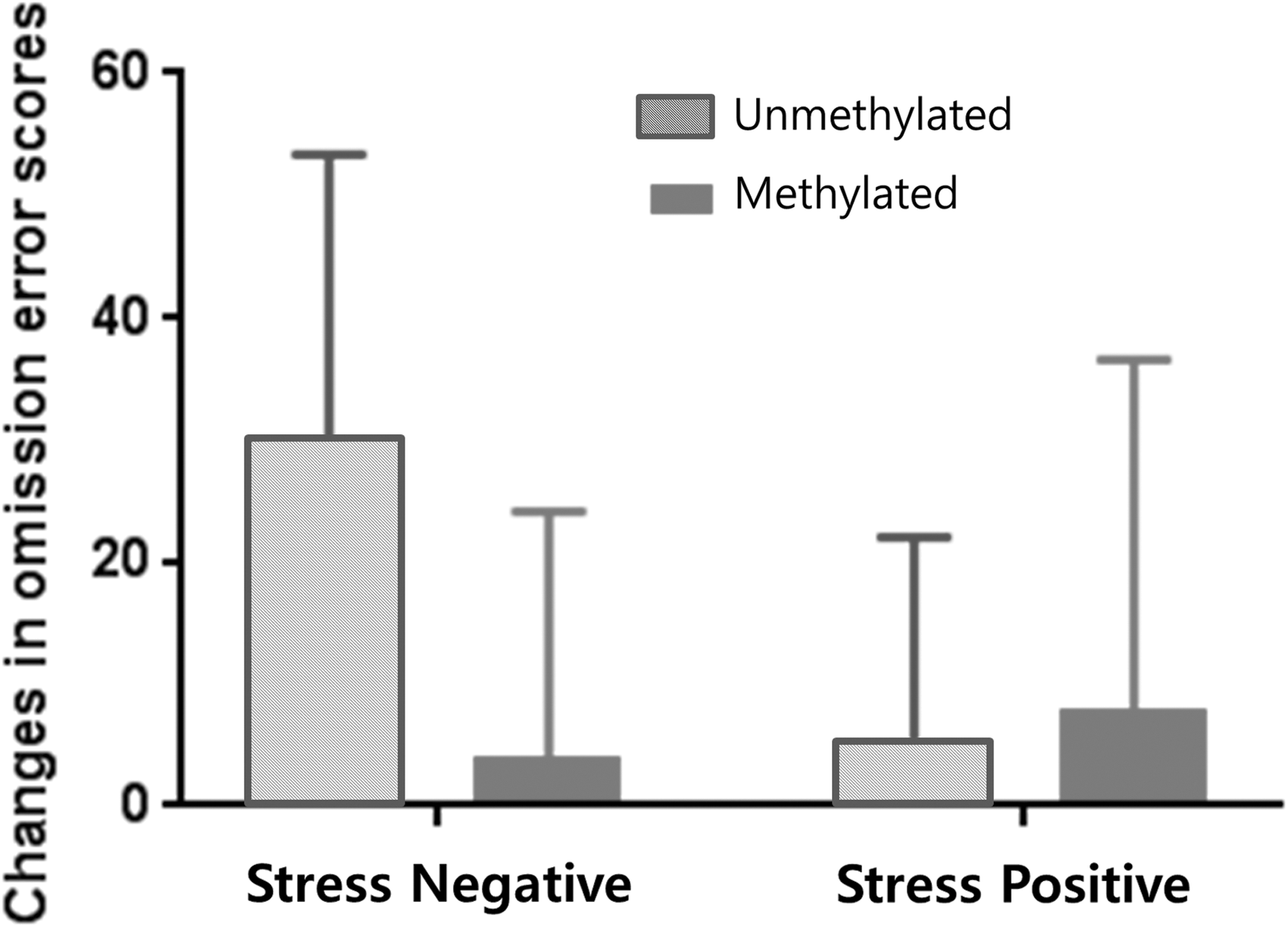
Changes in Continuous Performance Test omission error scores post-treatment according to CpG7 methylation in stress negative and stress positive group.
p-Value <0.0002.
CI, confidence interval.
Finally, the 4R-homozygote group of the DRD4 variant was not significantly associated with treatment response or change in score on the CPT (Supplementary Table S2). Additionally, there were no significant interactions between the DRD4 4R genotype and prenatal maternal stress for any treatment response variable.
Discussion
To our knowledge, this is the first pharmaco-epigenetic study to investigate the effects of the interaction between methylation status and environmental factors on the MPH treatment response in ADHD youth. There was a significant interaction effect between the methylation status of CpG7 and prenatal maternal stress on change in omission errors on the CPT. Contrary to the results reported by Cheon et al. (2007) based on the investigation of 83 ADHD patients, the present study did not find a significant association between the DRD4 4R allele and treatment response. These results emphasize the importance of considering epigenetic processes rather than genotype per se when predicting the treatment response of a patient.
Methylation processes can affect gene expression and mediate the effects of gene–environment interactions (Jones and Baylin 2002). The increased methylation of the DRD4 promoter region typically leads to the suppression of gene transcription; therefore, a decrease in the density of D4 receptors could alter the sensitivity to MPH (Goll and Bestor 2005; Anier et al. 2010).
However, because functional analyses of individual CpG sites were not conducted in the present study, it is difficult to determine how methylation influenced gene expression. A study conducted by Xu et al. (2015) found that children with methylated CpG1 exhibited a decrease in the extent of DRD4 expression but the expression levels of other CpG sites according to methylation were not examined in that study. As it is thought that the methylation status of critical CpG sites, rather than the entire CpG island, is involved in gene regulation, further studies on the role of each CpG site on gene expression are needed to clarify this relationship (McGowan et al. 2009; Murgatroyd et al. 2009).
The mechanisms underlying the interaction between prenatal maternal stress and DRD4 methylation are not yet well understood. An increasing amount of evidence indicates that stress causes a downregulation of 11β-hydroxysteroid dehydrogenase type 2, which is an enzyme responsible for the metabolization of cortisol in the placenta (O'Donnell et al. 2012), which results in high glucocorticoid levels in the fetus. Because the developing brain is vulnerable to adverse intrauterine conditions, an altered endocrine milieu may result in the reprogramming of the hypothalamic–pituitary–adrenal (HPA) axis via methylation of DNA encoding for glucocorticoid receptors (Zucchi et al. 2013). Animal studies have demonstrated that glucocorticoid receptors act as transcription factors for dopaminergic genes and directly influence the plasticity of dopaminergic pathways (Hensleigh and Pritchard 2013).
Among the epigenetic processes, alterations in CpG methylation appear to have the longest duration (Oh et al. 2013). As the methylation profiles are established early in life and then maintained during differentiation (Guo et al. 2011), they likely represent the effects of early life adversity, such as an adverse maternal environment (Oberlander et al. 2008). As one's methylation profile is relatively stable, it was postulated that prenatal maternal stress and DRD4 methylation status during childhood or adolescence may interact despite the difference in time (Walton et al. 2017). However, environmental risk factors, such as parenting style, parental psychiatric disorders, and home environment, often co-occur with maternal stress (Grizenko et al. 2012); therefore, future studies must consider the effects of co-occurring prenatal risk factors.
Studies investigating the effects of environmental factors on the MPH treatment response in ADHD populations are scarce. A study by Grizenko et al. (2008) did not find any difference in MPH response status according to the severity of prenatal maternal stress, which suggests that it may not be related to early environmental factors. However, this study was only a 2-week trial and may have been too short to achieve a sufficient treatment response in ADHD participants. Pagerols et al. (2017) identified a strong association between prenatal smoking and MPH treatment outcome and significant interaction effects between prenatal smoking and dopamine-related genes, including the dopamine receptor D3 (DRD3), dopamine beta-hydroxylase (DBH), and tyrosine hydroxylase (TH) genes, on MPH response.
However, neither of these studies considered the interaction of prenatal maternal stress with genetic or epigenetic factors. Future studies investigating the treatment response to MPH should consider the effects of various environmental factors that increase susceptibility to ADHD (e.g., prenatal smoking and obstetrical complications) and the interaction of environmental and genetic factors (Ben Amor et al. 2005).
Few epigenetic studies have focused on ADHD; moreover, studies investigating the relationship between the methylation of DRD4 and ADHD have produced inconsistent results. For example, Van Mil et al. (2014) reported that lower DNA methylation levels are associated with more ADHD symptoms but not with formal ADHD diagnoses, whereas Xu et al. (2015) demonstrated that higher methylation levels increase the risk of ADHD susceptibility. Furthermore, Dadds et al. (2015) showed that higher methylation levels are related to a greater degree of inattention. These discrepant results may be due to differences in sample size, sampling periods, tools used to measure symptoms, characteristics of the participants, and use of varied tissue sources (Dadds et al., 2015). Cautious interpretation of these results is required as methylation may differ according to environmental exposure and ethnicity.
This study has several limitations that should be considered. The small sample size may have underpowered the results of the statistical analyses and further replication studies with larger sample sizes are needed. Additionally, this study included only patients with a Korean genetic background, which may limit the generalizability of the findings regarding ADHD to other ethnicities. Owing to the lack of a placebo arm, psychological factors that have an impact on treatment response could not be ruled out either. Because DNA methylation was assessed using peripheral blood DNA, differences in the epigenetic profiles according to tissue type could not be considered and confirmation of this study results using postmortem brain tissues may better illustrate the mechanisms that are involved.
Moreover, methylation status was treated as a categorical variable, whereas it is continuous in nature. The hemi-methylated and fully methylated groups were combined into a single group, as the number of fully methylated groups was too small for statistical analyses in several CpG sites. Therefore, it would be hard to determine the effects of methylation according to degree of methylation, and further studies that separately analyze methylation groups or measure methylation in a continuous matter (e.g., pyro-sequencing) are required. Additionally, prenatal maternal stress was retrospectively assessed based on a dichotomous question, which likely led to a recall bias and may have limited the assessment of stress severity.
Finally, the functional relevance of the methylation of each CpG site (e.g., mRNA expression or protein expression) was not investigated and limited insights were obtained regarding the neural mechanisms underlying the study results. Further replication studies with large sample sizes and randomized placebo-controlled designs that include patients of various ethnic backgrounds are needed to help determine the validity of the interaction between prenatal maternal stress and DRD4 methylation as a biological marker of MPH treatment response.
Conclusions
In conclusion, this study provides preliminary evidence that the interaction between prenatal maternal stress and methylation in a CpG island of the DRD4 promoter region was associated with improvements in inattention following MPH treatment. Further investigations of prenatal stress and the methylation profiles of the DRD4 promoter region, particularly CpG7, may contribute to improving the prediction of individual treatment responses to MPH.
Conclusions and Clinical Significance
The genetic makeup, environmental factors, as well as their interaction may contribute to the inter-individual variability in treatment response. This study investigates the interaction between the methylation of the dopamine receptor D4 (DRD4) and prenatal maternal stress on MPH response of youth with ADHD. There was a significant interaction between methylation of CpG7 and prenatal maternal stress on changes in inattention following MPH treatment. These results emphasize the importance of considering epigenetic processes rather than genotype per se when predicting the treatment response of a patient.
Footnotes
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning (NRF-2015R1A2A2A01004501 to J.-W.K.), by Promising-Pioneering Researcher Program through Seoul National University (SNU) in 2015 to J.-W.K., and by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (No. 2015M3C7A1028926 to B.-N.K).
Disclosures
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.