
Abstract
Objectives:
Key neurobiological factors contribute to vulnerability to nonsuicidal self-injury (NSSI) among adolescents and how they respond to treatment targeted to reduce such behaviors. This study aims to examine differences in intrinsic functional connectivity between adolescents with NSSI and healthy controls (HCs) and to identify baseline connectivity markers that predict improvements in NSSI after psychotherapy.
Methods:
Adolescents aged 12–17 (n = 24) with repetitive NSSI along with demographically similar HCs (n = 16) underwent resting-state functional MRI scanning after which patients received up to 4 months of psychological treatment. A seed-based approach was used to examine baseline between-group differences in intrinsic functional connectivity of the amygdala and the medial prefrontal cortex (mPFC). Further analyses examined the associations between intrinsic functional connectivity at baseline and improvement in NSSI after psychological treatment.
Results:
Compared with HCs, adolescents with NSSI showed significantly reduced connectivity between the amygdala and the anterior cingulate cortex, subcallosal cortex, and paracingulate gyrus, as well as between the amygdala and a cluster encompassing the right planum temporale and right insula. Adolescents with NSSI, compared with HCs, also showed reduced connectivity between the mPFC and two clusters: one located in the precentral and postcentral gyri and another in the left insula. After treatment, 50% of patients reported fewer NSSI episodes compared to baseline, which was considered as improvement. Stronger negative amygdala-prefrontal connectivity was associated with greater posttreatment improvement in NSSI.
Conclusions:
Adolescents with NSSI may have aberrant amygdala and mPFC connectivity compared with HCs. Furthermore, stronger baseline negative amygdala-prefrontal connectivity may predict greater improvement in NSSI after psychological intervention. Given that no prior study has used resting-state functional connectivity to predict response to psychological treatment in adolescents with NSSI, replication of these findings is needed.
Introduction
Nonsuicidal self-injury (NSSI) is defined as self-injurious behavior in the absence of any observable intent to die (Nock 2010; Brent et al. 2013). Clinical and scientific interest in NSSI among adolescents has increased dramatically in the last decade due to its rise in prevalence worldwide. Based on community studies, between 13% and 45% of adolescents have engaged in NSSI sometime in their lifetime (Nock 2010; Muehlenkamp et al. 2012). These rates are as high as between 40% and 60% among pediatric clinical samples. Since NSSI is a risk factor for future suicide attempts (Asarnow et al. 2011; Chesin et al. 2017), understanding the neurobiological basis of NSSI could help in developing earlier interventions to prevent suicide attempts and subsequently completed suicides.
NSSI is principally associated with dysfunctional emotional regulation (Klonsky et al. 2011; Nock 2010) and may represent a maladaptive means to regulate extreme emotional reactivity and to compensate for deficits in social problem-solving skills (Nock and Prinstein 2005; Nock and Mendes 2008; Edmondson et al. 2016).
Connectivity within fronto-limbic brain circuitry has been demonstrated to be involved in the regulation of emotion (Banks et al. 2007; Etkin et al. 2011). In particular, the amygdala is considered a central hub in a network of brain regions that plays a key role in emotional reactivity (Drevets 2003; Weir et al. 2012). Although there are few studies that examine the neural circuitry associated with NSSI (Martin et al. 2015), preliminary results in small samples of adolescents point to a high activation in the amygdala during emotion processing (Plener et al. 2012). These results are consistent with studies in adults with borderline personality disorder and NSSI showing amygdala hyperactivity with negative effect (Niedtfeld et al. 2010), and with physiological research which found high levels of distress in adolescent self-injurers (Nock and Mendes 2008).
Aberrant amygdala connectivity with frontal and occipital regions, supplementary motor area, and dorsal anterior cingulate has also been demonstrated in adolescents with NSSI (Schreiner et al. 2017). Specifically, the amygdala has connections to prefrontal subregions that downregulate emotional reactivity, such as the medial prefrontal cortex (mPFC) (Banks et al. 2007). The mPFC is involved in social rejection and self-referential processing (Eisenberger et al. 2011), as well as in impulsive and decision-making process (Bechara et al. 1994; Ingvar 1994; Domsalla et al. 2014), which are impaired in self-injurious individuals. Indeed, NSSI may arise as a need to compensate for deficits in social problem-solving skills (Nock and Mendes 2008), as evidenced by hyperactivation in the mPFC following a social exclusion task in depressed adolescents with NSSI (Groschwitz et al. 2016).
In conclusion, these studies suggest that amygdala and mPFC hyperactivity and aberrant connectivity may be risk factors that predispose individuals who engage in NSSI to be more sensitive to stressful stimuli and to react more negatively to stress. Although these results provide some evidence for emotion-regulation deficits in the developing brain of adolescents with NSSI, more research is needed.
Importantly, both the amygdala and mPFC are highly sensitive to the effects of psychological treatments (Hölzel et al. 2009; Ritchey et al. 2011). Psychological interventions may lead to improved impulse control, decision-making, planning, and regulating of emotions, which may be a consequence of enhanced fronto-limbic connectivity. Some psychological interventions have shown to be effective in preventing NSSI (Ougrin et al. 2015); however, little is known about the brain-based characteristics that predict improvement following these interventions.
Considering these previous findings, the aims of this study were twofold: (1) to examine differences in intrinsic functional connectivity between fronto-limbic regions in adolescents with NSSI compared with healthy controls (HCs) and (2) to identify baseline intrinsic neural connectivity patterns associated with response to psychological interventions. To these aims, resting-state functional connectivity (RSFC), was used. The amygdala and mPFC were chosen as priori regions of interest (ROIs) based on the extant literature. We hypothesized that youth with NSSI would show impaired amygdala-mPFC connectivity compared with HC and that evidence of more adaptive functional connectivity in this network would be associated to response to psychological intervention.
Methods
Design overview
The study was approved by the Ethics Committee of the institution. All participants and their parents or legal guardians provided written informed consent and assent. Participants received a small monetary compensation for participating in each assessment.
Structural and functional MRI data were collected from all participants (patients and HCs) at baseline. Adolescents with repetitive NSSI were treated with psychological intervention for 16 sessions. The HC participants did not participate in any experimental intervention during the 16-week interval. The frequency of NSSI was assessed at baseline and at the end of treatment. Pretreatment RSFC maps were analyzed to identify predictors of improvement in pre- and posttreatment NSSI.
Participants
Twenty-eight patients were enrolled from an outpatient clinic of the Department of Child and Adolescent Psychiatry and Psychology at the Hospital Clinic in Barcelona. Eligible patients met the following inclusion criteria: (1) age between 12 years 0 months and 17 years 11 months, (2) presence of repetitive NSSI over the last 12 months, and (3) at least one parent or guardian willing to participate in sessions of psychological treatment. Exclusion criteria included the following: (1) intelligence quotient (IQ) below 70 on the Wechsler intelligence test (Wechsler 1997, 2003), (2) acute psychopathology requiring inpatient treatment at the time of screening, (3) low weight anorexia nervosa as determined following criteria Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision (DSM-IV-TR) (American Psychiatric Association 2000), (4) substance dependence (although concurrent substance abuse was not exclusionary) as determined by DSM-IV-TR (American Psychiatric Association 2000), and (5) orthodontic braces. Four participants withdrew during the study due to the following reasons: one moved to another city making it challenging to attend treatment, another was admitted to an inpatient mental health facility and could no longer participate, a third traveled to another country for more than a month, and the fourth withdrew for reasons unknown to the research team. Hence, the group of patients was composed of 24 participants.
Seventeen HC subjects were enrolled for study participation from community advertisements and local schools. Control subjects were matched by age, sex, and socioeconomic status to the patient group. Exclusion criteria included the following: (1) past or current psychopathology, (2) IQ below 70 on the Wechsler intelligence test (Wechsler 1997, 2003), (3) substance dependence (although concurrent substance abuse was not exclusionary) as determined by DSM-IV-TR (American Psychiatric Association 2000), and (4) orthodontic braces. One participant was excluded for unusable MRI data due to excessive motion artifact. The final sample was comprised by 16 HC participants.
All the participants were assessed using the same clinical diagnostic assessment (described below) and MRI procedures in the recruitment time period.
Clinical assessment
Patients and HCs were assessed by an independent and trained interviewer at baseline (M.P.). The assessment consisted of the structured Kiddie Schedule for Affective Disorders and Schizophrenia, Present and Lifetime version (K-SADS-PL) (Kaufman et al. 1997) (Spanish validated adaptation) interview to establish group status, obtain demographics, and DSM-IV-TR Axis I diagnoses. Parents and children were interviewed separately about the child.
NSSI was defined as self-injurious behavior in the absence of any observable intent to die (Nock 2010; Brent et al. 2013). The specific items assessing NSSI of the Columbia Suicide Severity and Rating Scale (C-SSRS) (Posner et al. 2011) were used to determine this behavior, and its frequency was rated by the patient's clinician using a scale designed specifically for this study. The scale to assess the frequency of NSSI consisted of a 6-point scale from 1 to 6 based on the type of responses used by the Suicidal Ideation Questionnaire-Junior (SIQ-Jr) (Reynolds 1987) and the C-SSRS to assess suicidal ideation: (1) No episodes of NSSI, (2) once a month, (3) couple of times a month, (4) once a week, (5) two to five times per week, and (6) daily. The frequency of NSSI at baseline was measured using this scale according to the information reported by patients and parents at each individual session and documented in the medical record in the 4 weeks before treatment. Current NSSI was assessed by the clinician every 4 weeks after the beginning of the study, based on the information reported by each participant and parents at each individual session. Although the scale was answered every 4 weeks, score improvement in NSSI was defined as the difference in NSSI recorded at baseline (covering the first 4 weeks before treatment) minus NSSI at 16 weeks (covering the last 4 weeks of treatment). Therefore, higher scores indicated better outcomes.
Other measures used were severity of suicidal ideation, measured by the 15-item self-report SIQ-Jr (suicidal thoughts rated on a 7-point scale from “I never had this thought” to “almost every day”) (Reynolds 1987); the level of depressive symptoms, assessed by the 21-item self-report Beck Depression Inventory-II (BDI-II) (Beck et al. 1996); and global level of functioning, measured by the Children's Global Assessment Scale (C-GAS) (Shaffer et al. 1983).
To ensure the reliability of the diagnostic assessment, all interviews were performed by the same psychologist (M.P.). The reliability of the diagnosis was evaluated by the clinicians in team meetings to build consensus. An interrater reliability analysis using the Kappa statistic was performed to determine consistency among raters. The interrater reliability for all symptoms rated with K-SADS was κ = 0.70 (p < 0.001), 95% CI (0.48–0.89).
Psychological interventions
Participants received one of two psychosocial interventions with no other concurrent psychological treatment: either components of Dialectical Behavior Therapy for adolescents (DBT-A) or Treatment-as-Usual (TAU) were administered over 16 weeks. Both treatments included at least biweekly individual sessions, 16 group sessions (1 session every week) for adolescents and parents attended separately, and medication management as needed. Components of DBT-A were conducted by clinicians trained in DBT who followed the manualized intervention protocol (Rathus and Miller 2014). TAU is defined as routine care usually provided by community-based adolescent mental health services. In addition, group sessions (where the needs of the participants were discussed) were added to TAU to control the intensity of the sessions of both treatments. Both treatments are commonly used in our outpatient clinic, and study therapists were specifically trained in both types of therapies.
Patients were randomized to either treatment, so 12 patients received DBT and the other 12 patients were treated using TAU and group sessions. Both treatment arms had similar responses and attrition patterns. To maximize power to investigate RSFC connectivity correlates of psychological treatment for NSSI, treatment arms were combined in our analysis.
Neuroimaging acquisition
Magnetic resonance imaging was performed on a 3-Tesla Siemens Magnetom Trio Tim (Siemens Medical Systems, Germany) scanner equipped with an 8-channel head coil. A high-resolution T1-weighted three-dimensional magnetization-prepared rapid sequence was acquired with the following sequence: 240 sagittal slices; repetition time (TR) = 2300 ms; echo time (TE) = 3.01 ms; slice thickness = 1 mm; field of view (FOV) = 240 × 240 mm; flip angle (FA) = 9°; and voxel size: 1 × 1 × 1. Functional images were collected with the following parameters: 32 slices; TR = 2000 ms; TE = 29 ms; slice thickness = 4 mm; FOV = 240 × 240 mm; FA = 80°; and voxel size: 3 × 3 × 3. The total scan time was 8 minutes 14 seconds. During the resting state-fMRI scans, all the subjects were instructed to remain as still as possible for the duration of the scanning session. A technician was engaged in conversation with the participant before and after the resting-state session to guarantee that they did not fall asleep.
Preprocessing
Preprocessing of resting-state data was carried out using FMRI Expert Analysis Tool (FEAT) Version 6.00 within FSL (FMRIBs Software Library). Each participant's 240-volume functional dataset was realigned to compensate for small head movements using Motion Correction using FMRIBs Linear Image Registration Tool (MCFLIRT) (Jenkinson et al. 2002) skull-stripped using Brain Extraction Tool (Smith 2002) spatially smoothed using a Gaussian kernel of 5 mm full width at half maximum, intensity normalized by a single multiplicative factor, and band-pass filtered to correct for baseline drift and high frequency noise (high-pass temporal filter: Gaussian-weighted least-squares straight line fitting, with sigma = 50.0 seconds; low-pass temporal filter: Gaussian with sigma = 2.8 seconds). Functional images were registered to participants' corresponding high-resolution T1-weighted structural images and then normalized to Montreal Neurological Institute (MNI) space using a 12-parameter transformation. Masks of white matter and cerebrospinal fluid (CSF) generated from each subject's anatomical images were applied to the functional data to extract white matter and CSF time series. These time series were used together with six motion parameters as nuisance regressors in a voxel-wise regression of the fMRI data. Data scrubbing was additionally performed following the method of Power et al. (2013) excluding any volume in which either the value for DVARS (the root mean squared change in blood-oxygen-level dependent (BOLD) signal from the prior volume) or the value for framewise displacement exceeded the upper boxplot threshold (the 75th percentile plus 1.5 times the interquartile range), along with the previous volume and the two following volumes. If at least 33% of the volumes were removed, participants would be excluded from analyses (no participants were excluded from the analyses for this reason). The number of censored volumes was not significantly different between the patient (M = 40.00, SD = 17.93) and control groups (M = 37.69, SD = 13.98) as indicated by t-tests [patient/control: t (38) = −0.435, p = 0.666]. Residuals of the voxel-wise regression were used in subsequent seed-based connectivity analyses.
Functional connectivity analysis: within subjects
Functional connectivity was examined using a whole-brain seed-based approach with two ROIs, the bilateral amygdala and the mPFC. The amygdala seed region was anatomically defined using a probabilistic map from the Harvard-Oxford Subcortical Structural Atlas, incorporating voxels that had 25% or greater probability of being labeled as the amygdala (left: 3456 mm3, right: 3936 mm3). The mPFC seed was defined as a 8mm-diameter sphere centered on previously published coordinates (mPFC: 0 52 − 6) as described by Van Dijk et al. (2010). The seed ROIs were registered to each participant's preprocessed fMRI data, and the mean time series of voxels in each ROI were extracted for each subject for use as primary regressors in a general linear model (GLM) analysis of all other voxel time series, resulting in individual whole-brain amygdala RSFC maps.
Functional connectivity analysis: between groups
Twenty-four patients and 16 HCs were included in the analysis. Although we collected functional connectivity data for the participants that withdrew from the study, they were not included in the functional connectivity between-group analyses. We focused only on the participants who continued with the treatment to analyze the relationship between functional connectivity at baseline and the change in frequency of NSSI at the end of the study. Group-level differences in RSFC for each seed were determined by voxelwise unpaired t-tests, covarying for age and sex. Resulting statistical maps were thresholded using a height threshold of Z > 2.0 and an extent threshold of p < 0.025, corrected using Gaussian random field theory. To avoid multiple testing concerns, all results were corrected for multiple comparisons using a Bonferroni-adjusted alpha level of significance of p = 0.025 for the two seeds: amygdala and mPFC. Parameter estimates (proportional to fMRI signal change) of BOLD signal response were extracted separately for each cluster and for every participant using featquery. Parameter estimates from the emergent clusters were fed into separate multivariate GLMs in SPSS (v.22) that modeled the parameter estimates as the dependent variables and age, sex, and group as independent variables.
Exploratory analyses
Continuous variables were compared between the two groups using independent-sample t-tests, whereas categorical variables were analyzed using the chi-square test. To test the normality of the sample distribution, the Kolmogorov–Smirnov test was used, together with the Levene test to assess the equality of variances.
Within the patient group, intersubject differences in functional connectivity were investigated using a voxel-wise GLM analysis that modeled each individual's change in NSSI as a covariate of interest and age and sex as covariates of no interest. Gaussian random field theory was used to correct for multiple comparisons with a cluster threshold of Z > 2.0 and p < 0.025. These higher level analyses generated thresholded Z-statistic maps of those voxels exhibiting significant NSSI change-related variation in coactivation with the amygdala and with the mPFC.
Results
Participant characteristics
Participants were composed of 24 adolescents with repetitive NSSI and 16 HCs. There were no significant group differences in age, gender, or socioeconomic status (see Table 1 for participant characteristics). The mean frequency of NSSI at baseline for patient participants based on a 6-point scale was 2.75 (SD = 1.2). A significant difference was found in IQ [t (36) = 3.780, p = 0.001] among groups; HC youth showed higher average IQ (M = 113.57, SD = 10.93) compared with patients with NSSI (M = 98.88, SD = 11.90) (see Table 1 for participant characteristics). However, covarying for IQ did not significantly change our primary results.
Baseline Demographic and Clinical Variables
Seventeen participants were on more than one medication.
Statistic computed using ANOVA.
Statistic computed using χ2 test.
Any anxiety disorder, including panic disorder, phobias, generalized anxiety disorder, and obsessive-compulsive disorder. Antipsychotic: quetiapine (n = 13), risperidone (n = 2), and aripiprazole (n = 3). Antidepressant: fluoxetine (n = 8), sertraline (n = 4), trazodone (n = 1), clomipramine (n = 1), and citalopram (n = 1).
ADHD, attention-deficit/hyperactivity disorder; BDI-II, Beck Depression Inventory-II; C-GAS, Children's Global Assessment Scale; DSM-IV, Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition; HC, healthy control; NSSI, nonsuicidal self-injury; PTSD, post traumatic stress disorder; SIQ-Jr, Suicidal Ideation Questionnaire-Junior; SES, socioeconomic status.
Differences in RSFC between patients and HCs
Our seed-based functional connectivity analyses revealed significant group differences in connectivity between the amygdala and a collective bilateral cluster composed of the anterior cingulate cortex (ACC), subcallosal cortex, and paracingulate gyrus (k = 1198 voxels, peak x/y/z MNI coordinates = 6/30/−2). Groups were also found to differ in functional connectivity between the amygdala and a cluster encompassing the right planum temporale and right insula (k = 703 voxels, peak x/y/z MNI coordinates = 56/−26/14, Fig. 1). Differences between patient and HC groups in mPFC connectivity were found in two clusters: one located in the precentral and postcentral gyri (k = 1022 voxels, peak x/y/z MNI coordinates = 14/−16/56) and another in the left insula (k = 1022 voxels, peak x/y/z MNI coordinates = −34/−8/12, Table 2 and Fig. 2).
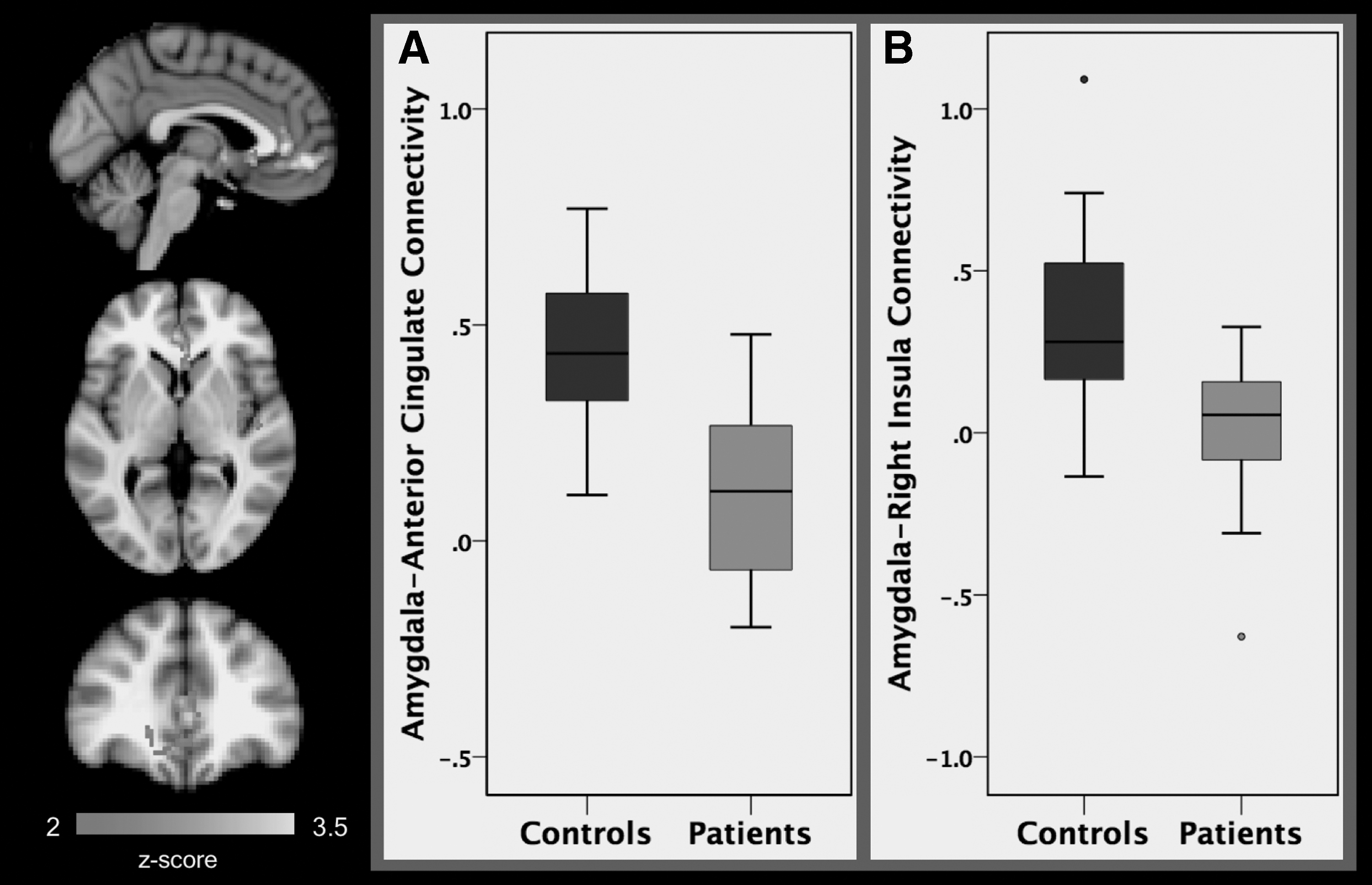
A comparison of patients (N = 24) and healthy controls (N = 16) found significant differences in average resting-state functional connectivity between the amygdala seed and the ACC, subcallosal cortex, and paracingulate gyrus
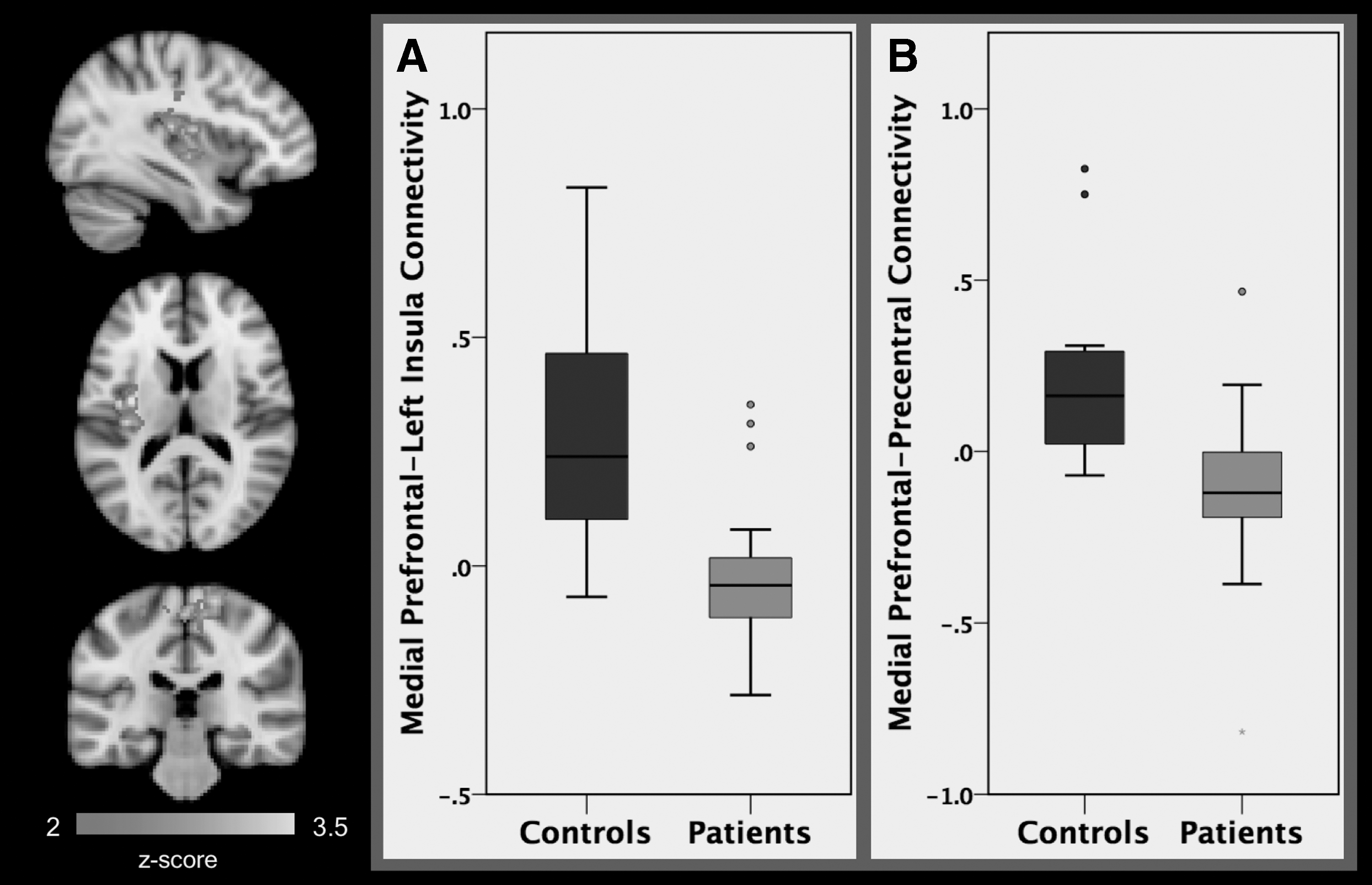
A comparison of patients (N = 24) and healthy controls (N = 16) found significant differences in average resting-state functional connectivity between the mPFC seed and the left insula
Differences in Functional Connectivity Between Patients and Healthy Control at Baseline
ACC, anterior cingulate cortex; MNI, Montreal Neurological Institute; mPFC, medial prefrontal cortex; ROI, region of interest.
Extracted connectivity estimates were analyzed using general linear models in SPSS, including age and sex as covariates. Higher connectivity was found between the amygdala and the ACC, subcallosal cortex, and paracingulate gyrus among HCs compared with patients. Higher connectivity between the amygdala and the right planum temporale and the right insula was also seen in the HC group compared with the patient group (Fig. 1). Higher positive connectivity was furthermore found between the mPFC and the precentral and postcentral gyri, as well as between the mPFC and the insula found in HCs relative to patients (Table 2 and Fig. 2).
In addition, GLM analyses were repeated with the outlier participants removed to test whether the group differences in RSFC were being driven by outliers. All between-group differences in RSFC remained significant (p < 0.025).
RSFC at baseline associated with change in NSSI
All 24 patients who participated in the study had experienced repetitive NSSI during the 12 months before treatment. Of those, 12 (50%) reported fewer NSSI episodes at the end of the treatment compared with baseline, which we consider as improvement. Ten (41.6%) participants displayed no change in NSSI frequency throughout the study and, thus, received an NSSI score of zero. Of these 10 participants, 6 reported no NSSI at baseline or at the end of the study. Four of the 10 participants who displayed no change had reported NSSI at baseline and maintained the same frequency of episodes throughout the treatment. Finally, two participants (8.3%) worsened during the treatment.
Improvements in NSSI, as quantified by changes in frequency of NSSI from pre- to posttreatment, were associated with variation in resting-state connectivity between the amygdala and the ACC, subcallosal cortex, paracingulate gyrus, and frontal medial cortex (k = 910 voxels, peak x/y/z MNI coordinates = −4/32/−4), as well as with variations in connectivity between the amygdala and a cluster in the brainstem and in the parahippocampal gyrus (k = 764 voxels, peak x/y/z MNI coordinates = 26/−30/−38). Improvements in NSSI were associated with greater mPFC connectivity with a cluster encompassing portions of the bilateral amygdala and striatum (k = 693 voxels, peak x/y/z MNI coordinates = 20/−4/−20) and with the bilateral intracalcarine/occipital cortex (k = 1781 voxels, peak x/y/z MNI coordinates = 6/−84/4).
When age- and sex-adjusted amygdala and mPFC connectivity estimates were extracted from the emergent clusters and analyzed using Spearman's correlation coefficients, they were found to correlate significantly with improvements in NSSI (r s = −0.642, p = 0.001; Fig. 3). In addition, adjusting for IQ and the number of individual and group sessions attended did not change the significance of these results. Greater negative or anticorrelation of RSFC between the amygdala and ACC connectivity at baseline was associated with significantly greater improvements in NSSI from pre- to posttreatment (r s = −0.642, p = 0.001). Conversely, positively correlated activity between the amygdala and ACC at baseline was associated with less improvement in NSSI. Greater positive amygdala-brainstem connectivity was associated with greater improvements in NSSI (r s = 0.747, p < 0.001). Reduced positive connectivity and greater negative connectivity between the mPFC and amygdala were associated with greater improvements in NSSI (r s = −0.630, p = 0.001). In addition, reduced positive and greater negative connectivity between the mPFC and the intracalcarine cortex was associated with greater improvements in NSSI (r s = −0.471, p = 0.02) (Fig. 3).
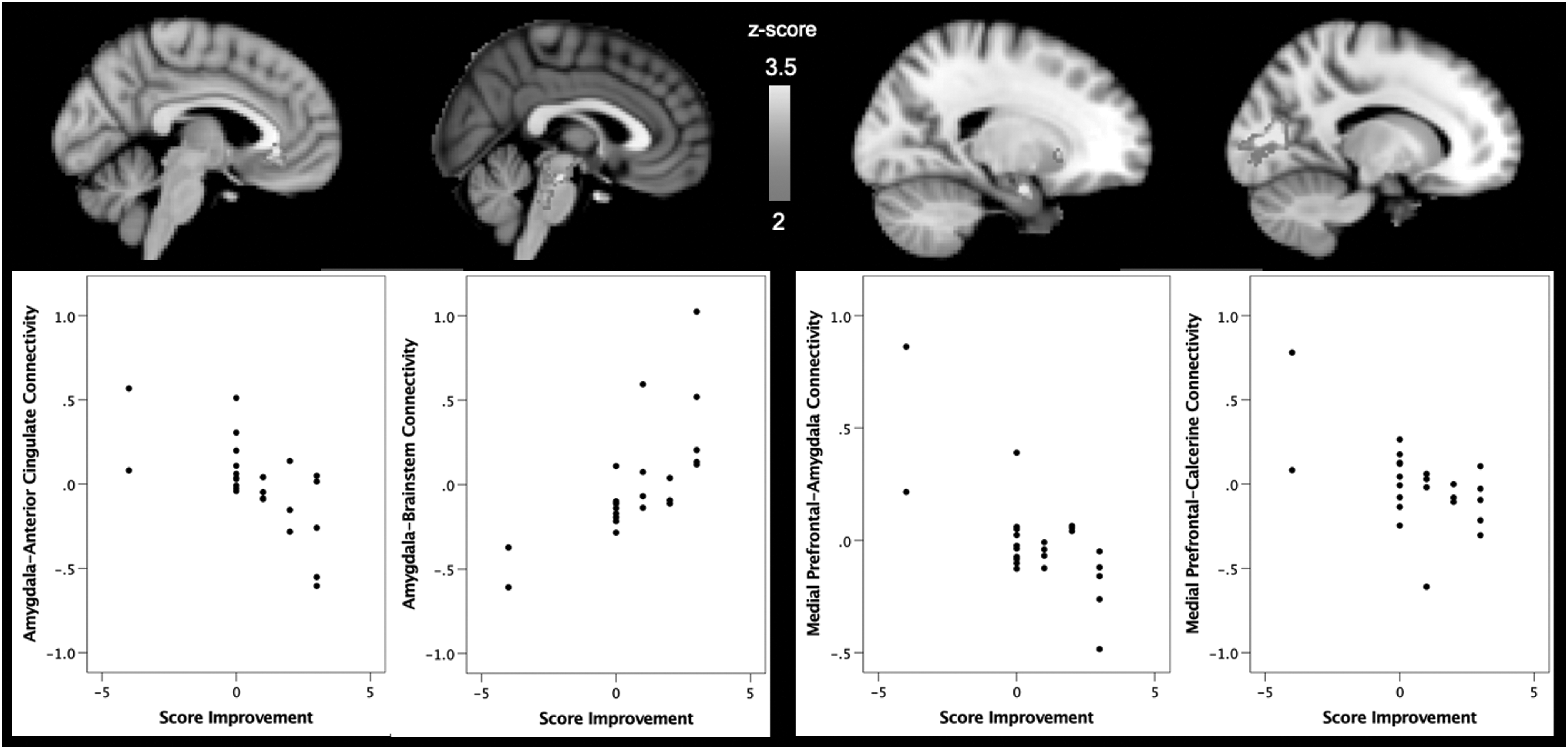
Within the patient group (N = 24), interindividual differences in improvement after treatment (based on the change in self-harming behavior from pre- to posttreatment) were associated with variation in intrinsic connectivity of the amygdala and the mPFC. Analyses controlled for age and sex.
Differences in RSFC and depression severity
BDI-II questionnaires were given to all of the participants. However, two participants did not return the questionnaire to the assessor at the beginning, and two others did not do so at the end of treatment. Consequently, pre- and posttreatment BDI-II scores were analyzed from 20 of the 24 patients. Change in depression severity was quantified as the difference between pre- and posttreatment BDI-II scores. The relationship between depression severity and connectivity was assessed in the patient group using Spearman's rho correlations with age- and sex-adjusted RSFC estimates and BDI-II change scores. No associations were found between amygdala or mPFC connectivity estimates and changes in depression scores: amygdala-ACC connectivity (rs = −0.196, p = 0.409), amygdala-brainstem/parahippocampal connectivity (rs = −0.046, p = 0.848), mPFC-amygdala connectivity (rs = −0.018, p = 0.940), or mPFC-intracalcarine connectivity (rs = −0.041, p = 0.865).
Differences in RSFC and suicidal behavior
Pre- and posttreatment SIQ and C-SSRS (suicidal behavior) were collected, and change in SIQ and C-SSRS was quantified as the difference between pre- and posttreatment scores. Spearman's rho was used to explore the relationship between age- and sex-adjusted RSFC estimates and changes in SIQ scores. Medial PFC-amygdala connectivity was found to correlate with SIQ change score (r s = −0.541, p = 0.011); no other correlations were found with RSFC. Using also Spearman's rho, no correlations were found between RSFC and changes in C-SSRS (all ps > 0.05).
SIQ was lower at the end of the treatment for the patient group; however, improvement on the SIQ was not significant between baseline and end of treatment. In contrast, C-SSRS quantified suicide severity was significantly improved from baseline to the end of the treatment [t (22) = 2.659, p = 0.014].
Effects of medications on NSSI
Eighteen patients were taking medication before the beginning of the study. Patients maintained stable dosing of medications during the study, and this did not significantly contribute to changes in scores or baseline connectivity estimates. Specifically, we ran analyses to evaluate the potential role of medication (antipsychotics or antidepressants) in connectivity and NSSI change separately. There were no statistically significant differences in groups that were exposed versus were not exposed to either antidepressants or antipsychotics (all p-values >0.23). We analyzed the distribution of medication (number of psychiatric medications, dosage of antipsychotics and antidepressants) among patients who received DBT (N = 9) and TAU (N = 9). There were no differences between both groups of treatment at baseline (all p-values >0.73).
Discussion
This is the first study conducted to identify brain connectivity predictors of treatment response using RSFC in adolescents with NSSI. As hypothesized, adolescents with NSSI showed significantly varying connectivity patterns in amygdala and prefrontal networks compared with HCs. In addition, stronger negative amygdala-prefrontal connectivity at baseline was associated with greater posttreatment improvement in NSSI. These findings provide new insights on how baseline amygdala-prefrontal network connectivities may predict psychological treatment outcomes.
Our findings are consistent with previous studies which have identified aberrant structural and functional changes in amygdala and regulatory prefrontal networks that are associated with the pathophysiology of NSSI in both adolescents (Groschwitz and Plener 2012; Plener et al. 2012; Schreiner et al. 2017) and adults (Niedtfeld et al. 2010). In particular, our study showed decreased connectivity between the amygdala and the ACC among adolescents with NSSI. This result could reflect that the ACC has a diminished capacity to inhibit amygdala activity (Sotres-Bayon et al. 2004; Pezawas et al. 2005). This is congruent with the fact that adolescents with NSSI often present high hypersensitivity to emotional stimuli (Plener et al. 2012). Impaired amygdala-prefrontal connectivity has also been found in adolescents with emotional difficulties related to NSSI, such as in depression (Connolly et al. 2017) or suicidal behavior (Cox Lippard et al. 2014; Johnston et al. 2017). Hence, our findings confirm the importance of the amygdala-prefrontal neural circuit in the expression and regulation of emotional behavior (Weir et al. 2012).
Our data also showed aberrant connectivity between amygdala and insula in adolescents with NSSI. The insula, specifically, plays a key role transmitting information about the salience of internal and external stimuli, including pain perception (Schreiner et al. 2015). It has been suggested that patients who engage in NSSI have altered pain perception (Ballard et al. 2010). Aberrant amygdala-insula connectivity may indicate deficits in attending to salient external stimuli, such as the physical pain inflicted from NSSI. This finding is consistent with a previous study that observed dysfunction of the insula in young adults with NSSI (Bonenberger et al. 2015), suggesting that amygdala-insula connectivity may be an important biomarker of NSSI in adolescents.
Youth with NSSI, compared with HCs, also showed decreased connectivity between mPFC and precentral and postcentral gyri. To our knowledge, these data have not previously been reported in the NSSI literature. Specifically, the precentral gyrus consists of the primary motor cortex, which is responsible for voluntary movement (Exner et al. 2002), cognitive processing and emotion regulation (Seo et al. 2014), and avoidance of emotional suffering (Li et al. 2017). The decreased mPFC precentral gyrus connectivity could represent a voluntary behavior that allows these youths to reduce and modulate their emotional responses and avoid the suffering associated with negative events. With regards to the mPFC postcentral gyrus network, the postcentral gyrus is the key receptor of general bodily sensation (Webb 2017). Decreased mPFC postcentral gyrus connectivity may reflect aberrant perception of bodily sensations associated with NSSI. Further studies are required to confirm this interpretation.
We also found that patients who engage in NSSI showed decreased mPFC left insula connectivity compared with HCs. This connectivity has been implicated in pain processing (Zhang et al. 2014). Similar to the aberrant amygdala-insula connectivity mentioned above, altered mPFC-insula connectivity may contribute to impaired pain perception in adolescents with NSSI.
In our second objective, we set out to identify baseline connectivity patterns that would predict response to psychological intervention for NSSI. Our results confirm a bidirectional relationship between the amygdala and mPFC, in association with improvement in NSSI after psychotherapy. These results suggest that the decreased bidirectional amygdala-prefrontal connectivity at baseline may predict response to psychotherapy in adolescents who engage in NSSI. To our knowledge, this is the first study to analyze baseline connectivity using functional neuroimaging in adolescents with NSSI who received psychotherapy. Frontal regions, such as the mPFC, are involved in control functions (Bechara et al. 1994; Ingvar 1994; Domsalla et al. 2014). A negative functional connectivity between frontal regions and the amygdala was described in adolescents with difficulties regulating emotions, such as in depression (Connolly et al. 2017) or suicidal behavior (Johnston et al. 2017). This finding was interpreted according to the hypothesis of “top-down” regulation of amygdala activity by these control regions (Huang et al. 2014). In addition, reduced connectivity between the amygdala and frontal regions was associated with poor ability to regulate emotions (Luking et al. 2011). Consequently, our results suggest that psychotherapy may enhance skills associated with mPFC and increase the ability to regulate emotions, which is in accordance with the “top-down” emotion regulation mechanism. These results were confirmed in a sample of adults with depression, where an increase in mPFC activity was found after receiving psychotherapy (Ritchey et al. 2011).
In addition, the magnitude of functional connectivity from the mPFC to the amygdala was negatively associated with a change in suicidal ideation. This is congruent with findings obtained in a previous study with adolescents (Johnston et al. 2017), where a reduction in amygdala-prefrontal connectivity was associated with greater severity of suicidal ideation. An increase in suicidal ideation was observed in adults with dysfunction in cortical midline structures, which include the mPFC (Marchand 2012). These regions are also implicated in self-referential thinking and emotional regulation (Gusnard et al. 2001). Maladaptive skills in regulating emotions increase the likelihood of suicidal ideation and behavior, and adaptive skills decrease suicidal thoughts and acts (Tamás et al. 2007). Given that PFC regions are involved in the development of adaptive regulatory responses (Luna et al. 2010), emotion dysregulation may likely be associated with dysfunction in prefrontal connectivity and may be a cause of suicidal ideation. As suggested above, enhancing skills associated with mPFC using psychotherapy could develop more adaptive abilities to cope with stressful events and may reduce both suicidal ideation and NSSI.
Interestingly, we also found that greater positive baseline amygdala-brainstem connectivity was associated with NSSI improvement. The amygdala-brainstem network has been implicated in the processing of visceral information in relation to emotional stimuli and is important for arousal (Price 2003). To our knowledge, these data have not previously been reported in the NSSI literature. One prior study has shown decreased connectivity between both brain regions in adolescents with depression, which may be associated with vegetative symptoms (Cullen et al. 2014). In the current study, greater positive connectivity between the amygdala and brainstem may relate to a hyperarousal response observed in young self-injurers, following stressful events, which may also be sensitive to psychological intervention.
Finally, it should be noted that the majority of youth in our sample were diagnosed with a major mood disorder. Although depressive symptoms improved with psychosocial intervention, this improvement did not correlate with amygdala or mPFC connectivity patterns, as was observed with changes in NSSI. This may be due to the fact that improvements in NSSI are more specific to these connectivity patterns, whereas depression, broadly defined, is a heterogeneous construct (Goldberg 2011; Lieblich et al. 2015). Despite the close relationship between depression and NSSI, our results point to the potential importance of targeting phenotypes of treatment response and their specific underlying neural circuitry.
The findings of this study should be interpreted in the context of some limitations. First, the sample size was small, which significantly constrains the generalizability of our findings and the ability to investigate differential effects of various psychological treatments. Larger studies will be needed to confirm our findings. Second, given that treatment adherence ratings are unlikely to be part of routine clinical settings, as the present study was conducted, treatment fidelity was not formally assessed. DBT-A has been recently implemented in Spain, and at the time of the design of the study, there were no official assessors to assess treatment adherence systematically for DBT-A. TAU did not follow any specific manualized protocol; therefore, it was not systematically assessed, as in previous studies (Rossouw and Fonagy 2012; Mehlum et al. 2014). Despite the lack of a formal assessment of treatment fidelity, improvement in NSSI was observed. Finally, due to the cross-sectional nature of our imaging data acquisition, we were not able to determine whether response to treatment was associated with changes in functional connectivity between baseline and the end of treatment. We will aim to pursue this hypothesis in a future study to determine if response to treatment corresponds to improvement in functional connectivity in the areas targeted in the current study.
Conclusion
In conclusion, this study advances the understanding of baseline intrinsic brain network connectivity characteristics in adolescents with NSSI and provides data to support the concept that specific connectivity patterns may predict response to psychological intervention. To our knowledge, this is the first study conducted to identify amygdala-prefrontal connectivity predictors of treatment response using RSFC in adolescents with NSSI.
Clinical Significance
The ability to predict an individual's response to treatment would help clinicians to assign patients to more effective treatments and, consequently, optimize resources. The current study provides a step forward toward personalized treatment in adolescents with NSSI. In addition, it increases knowledge of the underlying mechanisms that are involved in NSSI.
Given the potential for neuroplasticity during adolescence, more research focused on this stage could aid in the development of novel interventions to promote adaptive emotional regulation.
Disclosures
Dr. Singh receives research support from Stanford's Child Health Research Institute, National Institute of Mental Health, National Institute of Aging, Neuronetics, Johnson and Johnson, and the Brain and Behavior Foundation. She is on the advisory board for Sunovion. Dr. Sugranyes has received research funding from the Brain and Behavior Foundation, Instituto de Salud Carlos III, and Catalonia Regional Government and travel support from Otsuka Pharmaceuticals, Janssen, and Adamed Farma. There are no conflicts of interest to disclose for the rest of the authors.
Footnotes
Acknowledgment
The authors are grateful to all the participants of this study.