
Abstract
Background:
Indole tryptophan metabolites (ITMs), mainly produced at the gastrointestinal level, participate in bidirectional gut–brain communication and have been implicated in neuropsychiatric pathologies, including attention-deficit/hyperactivity disorder (ADHD).
Method:
A total of 179 children, 5–14 years of age, including a healthy control group (CG, n = 49), and 107 patients with ADHD participated in the study. The ADHD group was further subdivided into predominantly attention deficit (PAD) and predominantly hyperactive impulsive (PHI) subgroups. Blood samples were drawn at 20:00 and 09:00 hours, and urine was collected between blood draws, at baseline and after 4.63 ± 2.3 months of methylphenidate treatment in the ADHD group. Levels and daily fluctuations of ITM were measured by tandem mass spectrometer, and S100B (as a glial inflammatory marker) by enzyme-linked immunosorbent assay. Factorial analysis of variance (Stata 12.0) was performed with groups/subgroups, time (baseline/after treatment), hour of day (morning/evening), and presence of depressive symptoms (DS; no/yes) as factors.
Results:
Tryptamine and indoleacetic acid (IAA) showed no differences between the CG and ADHD groups. Tryptamine exhibited higher evening values (p < 0.0001) in both groups. No changes were associated with methylphenidate or DS. At baseline, in comparison with the rest of study sample, PHI with DS+ group showed among them much greater morning than evening IAA (p < 0.0001), with treatment causing a 50% decrease (p = 0.002). Concerning indolepropionic acid (IPA) MPH was associated with a morning IPA decrease and restored the daily profile observed in the CG. S100B protein showed greater morning than evening concentrations (p = 0.001) in both groups.
Conclusion:
Variations in ITM may reflect changes associated with the presence of DS, including improvement, among ADHD patients.
Introduction
The neurobiological basis of attention-deficit/hyperactivity disorder (ADHD) is based on two complementary models, which are both based on the dysregulation of interacting neural pathways, that is, the inhibitory noradrenergic frontocortical activity on dopaminergic striatal structures and the ascending dopamine circuits in the limbic system. Although ADHD has a multifactorial origin with a strong genetic component (Biederman and Faraone 2005), there are age-related changes in discrete brain areas and connectivity that parallel behavioral improvement and increased efficiency in cognitive task performance (Matthews et al. 2014). The development and differentiation of the central nervous system (CNS) are crucially influenced by serotonin systems, which are involved in mood, behavioral, and cognitive functions (O'Mahony et al. 2015). Through the orbitofrontal-striatal circuitry, serotonin may regulate behavioral domains of hyperactivity and impulsivity in ADHD (Banerjee and Nandagopal 2015).
In addition to genetic factors, social and environmental risk factors (associated with proinflammatory changes) affect ADHD, especially when they are present in prenatal and early postnatal periods during brain development (Sagiv et al. 2013). Bacterial colonization of the intestine has a major role in the postnatal development and maturation of the immune and endocrine systems and is integrated into the programming of CNS development (Julio-Pieper et al. 2013). Early life biological stress and psychological stress can change the gut microbiota composition, with a bidirectional microbial–neuroendocrine relationship (Bailey et al. 2011).
In addition to protein synthesis, tryptophan metabolism follows the following main branches. In the main branch (∼90% or more of tryptophan metabolism), the oxidation by indoleamine-2,3-dioxygenase (IDO) breaks the indole ring, resulting in the formation of kynurenine and metabolites (kynurenines), nicotinic acid, and nicotinamide adenine dinucleotide (NAD+) synthesis (Stone 2016). In the other branch, 3%–10% of tryptophan metabolism keeps the indole ring intact while producing chemical messengers such as the indoleamines, serotonin and melatonin, and the trace amine tryptamine and its derivatives. Approximately 95% of serotonin is synthesized by enterochromaffin cells within the gastrointestinal (GI) tract, and 5% is synthesized in the CNS.
Serotonin may interact with the DA system regulating impulsivity, sensitivity to expressed emotion, and academic, interpersonal, and recreational functioning domains in ADHD (Sonuga-Barke et al. 2009). Several amines, including serotonin and the ITM tryptamine, arise directly from gut bacteria or indirectly from the actions of bacteria on dietary components. The supply and availability of tryptophan are essential for this bidirectional brain–gut axis signaling (O'Mahony et al. 2015), regulating monoamine transmission (Khan and Nawaz 2016). In this way, changes in microbiota have been recently related to ADHD (Cenit et al. 2017; Richarte et al. 2018).
Intestinal bacterial colonization has a major role in the postnatal development and maturation of the immune and endocrine systems, which are integrated into the programming of brain development; thus, this process becomes essential for CNS homeostasis (Julio-Pieper et al. 2013). The gut microbiota modifies the expression of risk genes and participates in mechanisms that alter cognitive functions (Garcia et al. 2010). Germ-free male animals have a significant elevation in the hippocampal serotonin and (5-hydroxy-indoleacetic acid, 5HIAA); this alteration is sex specific unlike the immunological and neuroendocrine effects that are present in both sexes (Clarke et al. 2013). Early postnatal development is a sensitive period in that the gut microbiota could modulate ACTH levels, affecting normal brain development and behavioral functions (Diaz Heijtz et al. 2011).
Inflammatory states, which can be triggered by stress (Miller et al. 2009), are associated with a disrupted intestinal barrier that facilitates the access of both dietary antigens and pathogens, leading to innate immune activation with increased cytokine production. Conversely, indole tryptophan metabolites (ITMs) originating from gut microbiota may protect intestinal barrier integrity (Kelly et al. 2015). Protein S100B, which is a cytokine-related neurotrophin that largely derives from astrocytes and oligodendrocytes, is involved in the regulation of glutamate and calcium uptake, neuronal plasticity, and energy metabolism, especially in regulation involving the neurotrophic role of serotonin (Stroth and Svenningsson 2015). Several proinflammatory states are associated with the activation of kynurenine pathway metabolism (Erny et al. 2015) and increased protein S100B. Similar to kynurenine pathway metabolism (Notarangelo and Pocivavsek 2017), in the first 2 years of life, the concentration of serum S100β is higher in younger children (Arroyo Hernandez et al. 2016), remaining stable at later ages. Overproduction of S100B (micromolar levels) can be toxic and follow or accompany severe mental disorders (Schroeter et al. 2013).
Because ADHD is highly more frequent in children suffering early adverse events of physical (proinflammatory states) or psychological stress (social disadvantage), the aims of our study were to determine the levels and morning–evening variations of ITMs (which may have a protective role) in addition to serum cytokine S100B (putative glial inflammatory marker) in children with ADHD, in comparisons with healthy children, and their possible variations according to both the type of ADHD and the presence of depressive symptoms (DS), before and after chronic methylphenidate treatment.
Methods
Subjects
Participants included an ADHD group in which each patient was assessed twice at baseline and after treatment and a control group (CG) as a reference. A total of 107 patients who met the Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition, Text Revision (DSM-IV-TR)/criteria for ADHD were included after completing the clinical protocol to exclude the main comorbidities. The CG (n = 41) was mainly composed of siblings (n = 35) of patients with ADHD (recruited simultaneously with their siblings) or unrelated participants (n = 6) who were healthy children with adequate academic performance.
Assessment
Each child with ADHD was assessed at least twice. We obtained a personal medical history and physical examination and administered the following assessment instruments: (1) DSM-IV-TR criteria checklist, which was completed by the child's teacher; (2) evaluation of deficit of attention and hyperactivity (EDAH) scale (EDAH is a Spanish acronym) (Sánchez et al. 2010) in duplicate, one by a teacher and the other by the child's parents; (3) the Children's Depression Inventory (CDI), which was completed by participants ≥8 years of age; and (4) a sleep diary, which was completed for 1 week. The EDAH contains the main criteria of DSM-IV-TR to aid in identifying children with ADHD and/or conduct disorder (CD). The EDAH is a 20-item scale (Farré-Riba and Narbona 1997) that uses structured observation by teachers and is divided into two 10-item subscales for ADHD and CD, respectively. Based on EDAH scores (Table 1), the ADHD group was quantitatively subclassified into the following clinical subgroups: children who had predominantly attention deficit (PAD) and children with predominantly hyperactive–impulsive subtype with comorbid CD (predominantly hyperactive impulsive, PHI). Accordingly, of the 78 children included in the PHI group, 34 (44%) met the criteria for the diagnosis of HI without CD. Of the 44 children with symptoms of CD, 33 showed a predominance of HI symptoms over CD symptoms, whereas the remaining children (11/78; 14%) had a predominance of CD symptoms over HI symptoms. Of the 78 children of PHI group, 52 (67%) also had criteria for AD, and therefore they can be considered of a combined ADHD type. As indicated previously, because externalizing behavioral problems and aggression seem to be related to the hyperactive–impulsive ADHD symptom domain and the overall ADHD symptom severity (Connor and Ford 2012), for statistical analysis, we added these patients to the PHI group. The PAD group consisted of 29 children, none of them with CD.
Antropometrics, ADHD, CDI, and Intellectual Scores of the Control and the ADHD Groups
Data are expressed as mean ± SD.
“Negative Mood” and “Negative Self-Esteem” are subscales of the CDI; Z: z-value on the Wilcoxon signed-rank test.
KBIT, combined punctuation of the Kaufman abbreviated intelligence test. ADHD subgroups were classified as follows: Attention deficit (AD) if score >9. Hyperactivity-impulsivity (HI), if score <10, and Global deficit scores if <30. PHI, if AD <10; H (hyperactivity) >9; and/or total punctuation >29.
M, male; F, female; ADHD, attention-deficit/hyperactivity disorder; BMI, body mass index; CDI, childhood depression inventory; PHI, predominantly hyperactive impulsive.
The d2 Test (Brickenkamp 1997), as an attentional dimension (particularly visual attention), scores processing speed, rule compliance, and performance quality, allowing for a neuropsychological estimation of individual attention and concentration performance by quantification of the following scoring keys: errors of omission and errors of commission.
The CDI is a self-report assessment of depression for children (Kovacs 1992) whose two subscales (Negative Mood and Negative Self-Esteem) contain items that are most unique to depression and least related to anxiety. For defining subgroups, we considered the sum of both subscales, with a quantitative cutoff of >17 points considered pathological. The DSs were assessed through interviews with the parents at baseline and in the clinical follow-up; symptoms were quantified by the CDI score fulfilled by each child. All children were evaluated with an abbreviated intelligence test (Kaufman Brief Intelligence Test, KBIT) and completed the Spanish version of the Sleep Diary of the National Sleep Foundation for 1 week (data not shown). The ADHD group completed the diary again after treatment.
Written informed consent was obtained from all parents and children ≥12 years, and informed assent was obtained from all participants. No control participant was treated with any drug for ethical reasons, and they were assessed only once. The study design was approved by the Hospital Ethics Committee and the Health Research Fund of Spanish Ministry of Science and Innovation.
The exclusion criteria were as follows: (1) Low intelligence (KBIT <85), (2) preexisting or current epilepsy treatment, (3) other ADHD treatments or other pathological conditions, and (4) revocation of informed consent.
Treatment
The only drug used was prolonged release methylphenidate (OROS formulation), initially at 0.5 mg/(kg·d). The dosage was adjusted as a function of response and tolerance to treatment. The mean initial dose of methylphenidate was 25.8 ± 10.3 mg, and the final dose at the time of the second evaluation was 31.8 ± 10.7 mg. At inclusion, all patients were naive of any medication, and no other treatment (pharmacological or psychological) was administered before the conclusion of the protocol.
Measurements
None of the samples were obtained in the presence of an acute or severe illness. Blood samples were taken at 20:00 and 09:00 hours the following day. In the ADHD group, after 4.6 ± 2.3 months of daily methylphenidate, an identical study protocol was repeated. Serum was separated into 0.5 mL aliquots for freezing at −80°C until analysis.
Analytical method
We assessed levels of ITM (tryptamine, indoleacetic acid, and indolepropionic acid [IPA]) by liquid chromatography–tandem mass spectrometry (LC–MS/MS) (Fazio et al. 2015). Before analysis, serum samples were filtered with a filter nylon of 0.22 μm. One hundred microliters of serum samples was deproteinized using 100 μL of internal standard (IS) working solution (50 μM in TCA 4%). Samples were vortex-mixed and centrifuged at 14,000 rpm for 5 minutes. Fifty microliters of the clean upper layer was injected into the chromatographic system. UPLC™ analysis was performed using an Acquity UPLC class (Waters, Manchester, United Kingdom) with a mass-mass detector XEVO TQS, which included a quaternary pump, autosampler, solvent degasser, and column oven. The mass spectrometry method was performed on an XEVO TQS system equipped with a Turbo Ion Spray source. The detector was set in the positive and negative ion mode. The capillary and cone voltages were set at 0.6 kV and 40 V, respectively; the source temperature was 150°C; and the desolvation temperature was 500°C. The instrument was set in the multiple reaction monitoring mode. Data were acquired and processed by MassLynk v4.1 software. The chromatographic conditions were as follows: Channel A, water added with 0.01% formic acid and ammonium 0.05%; Channel B, acetonitrile; flow, 0.2 mL/min; gradient, t0: 5% B; t3: 70% B; t5: 5% B; and run time: 9 minutes. Acquity BEH C18 1.7 μm 2.1 × 50 mm was used as a chromatography column. Analytic parameters were estimated by two calibration lines, with one line from 1 to 20 μg/L and the other line from 20 to 500 μg/L.
Protein S100B was measured by enzyme-linked immunosorbent assay (ELISA). Human S100B ELISA (Cat. No. RD 192090100; BioVendor Laboratory Medicine, Inc., Brno, Czech Republic) and RD 192090100 Human S100B ELISA were used. The sensitivity was 5 pg/mL, the assay sensitivity was 20 pg/mL, the specificity for human S100B was 100%, the intra-assay variability was 4.5%, and the interassay variability was 5.2%.
Statistics
To achieve the study objectives, we conducted factorial analysis of variance (ANOVA) as described hereunder. ANOVAs were made after a Box–Cox transformation to ensure the normality of the variable and to ensure the hypothesis of homogeneity of variances between groups, in all the analyses that required it. For comparisons between EDAH and CDI scores, ANOVA within subjects (ordinal variables) and Wilcoxon signed-rank tests (paired samples) were used for inferential statistics. For comparisons between patients (cases) and each study variable, the factors in the factorial models were as follows: (1) groups with two categories: PAD and PHI groups; (2) patients nested in ADHD presentation types groups and depression subgroups (CDI); (3) hour, with two categories (morning and evening), and crossed with ADHD group and DS subgroups; and (4) time, with two levels before and after treatment. Time was a crossed factor with subgroups and hour. Subgroups, hour, and time were fixed-effects factors, and patients was a random-effects factor. Comparisons between cases and controls were performed using the same analysis repeated in two different situations because there was only one measure for controls, which were as follows: at baseline and after treatment (in cases). The factorial model had the following three factors: (1) group with three categories (controls, PAD, and PHI); (2) subjects (controls and patients) nested in groups, and patients nested in CDI subgroups; and (3) hour, with two categories, morning and evening, that was crossed with group/subgroups. Group and hour were fixed-effects factors, and subjects random-effects factor. For both comparisons, an ANOVA table was built and higher interactions were determined. If these were significant, multiple pairwise comparisons were made using Bonferroni's correction, and if not, these corrections were applied to the principal effects in the table. The experimental quantities for these comparisons were not “t” as expected, because we have used “z,” the normal approximations for “t`s,” because of the global sample sizes. The reported analyses were crude analyses and adjusted analyses by age and sex, which were carried out using analysis of covariance methodology. In all cases, the interactions were studied for levels <0.15, and the latest comparisons were considered significant at p < 0.05 after applying the penalty provided by the correction. When analyzing the variances in different groups, homogeneous transformations were carried out on data using natural logarithms to achieve uniformity. We used the statistical package STATA 12.0 for all analyses.
Results
Anthropometrics, ADHD, CDI, and intellectual scores for ADHD and the CGs are given in Table 1. The mean heights and weights were significantly higher in the controls, partially because of the slightly higher mean age than that in the ADHD group. However, there were no body mass index (BMI) differences. After treatment, the average patient height was unchanged, whereas the weight decreased, which was expected and previously reported (Molina-Carballo et al. 2013). Although not significantly different, the prevalence of DSs was 20.7% in patients with PAD and 24.4% in PHI group; it was more common in girls (34.8%) than in boys (20.2%). More than 80% of participants experienced improvement in EDAH scores after methylphenidate treatment that were based on parent evaluation data.
Tryptamine
There were no differences in serum tryptamine values between our CG (28.57 ± 11.06 and 34.90 ± 9.27 ng/mL, morning and evening) and ADHD group (26.61 ± 8.46 and 32.65 ± 11.38 ng/mL, morning and evening) at baseline or after methylphenidate in the ADHD group (25.85 ± 10.32 and 30.99 ± 11.05 ng/mL, morning and evening) (Table 2). Tryptamine concentrations showed a daily profile with significantly higher evening values (vs. morning) in both the CG and ADHD groups (z = 5.05, p < 0.0001). This daily profile is not modified by methylphenidate, ADHD subgroups, or DS factor (Fig. 1). In urine, baseline tryptamine excretion in the ADHD group was very similar to that in the CG (0.55 ± 0.59 ng/mgCr). Methylphenidate was associated with an excretion decrease in tryptamine (in similar amount for all patients with ADHD) (Table 3) with a significant difference relative to baseline (z = 2.47, p = 0.01) and greater but nonsignificant excretion decrease in DS− (0.59 ± 0.34 and 0.44 ± 0.26 ng/mgCr, baseline and posttreatment) compared with that in DS+ (0.37 ± 0.18 and 0.33 ± 0.16 ng/mgCr, baseline and posttreatment).
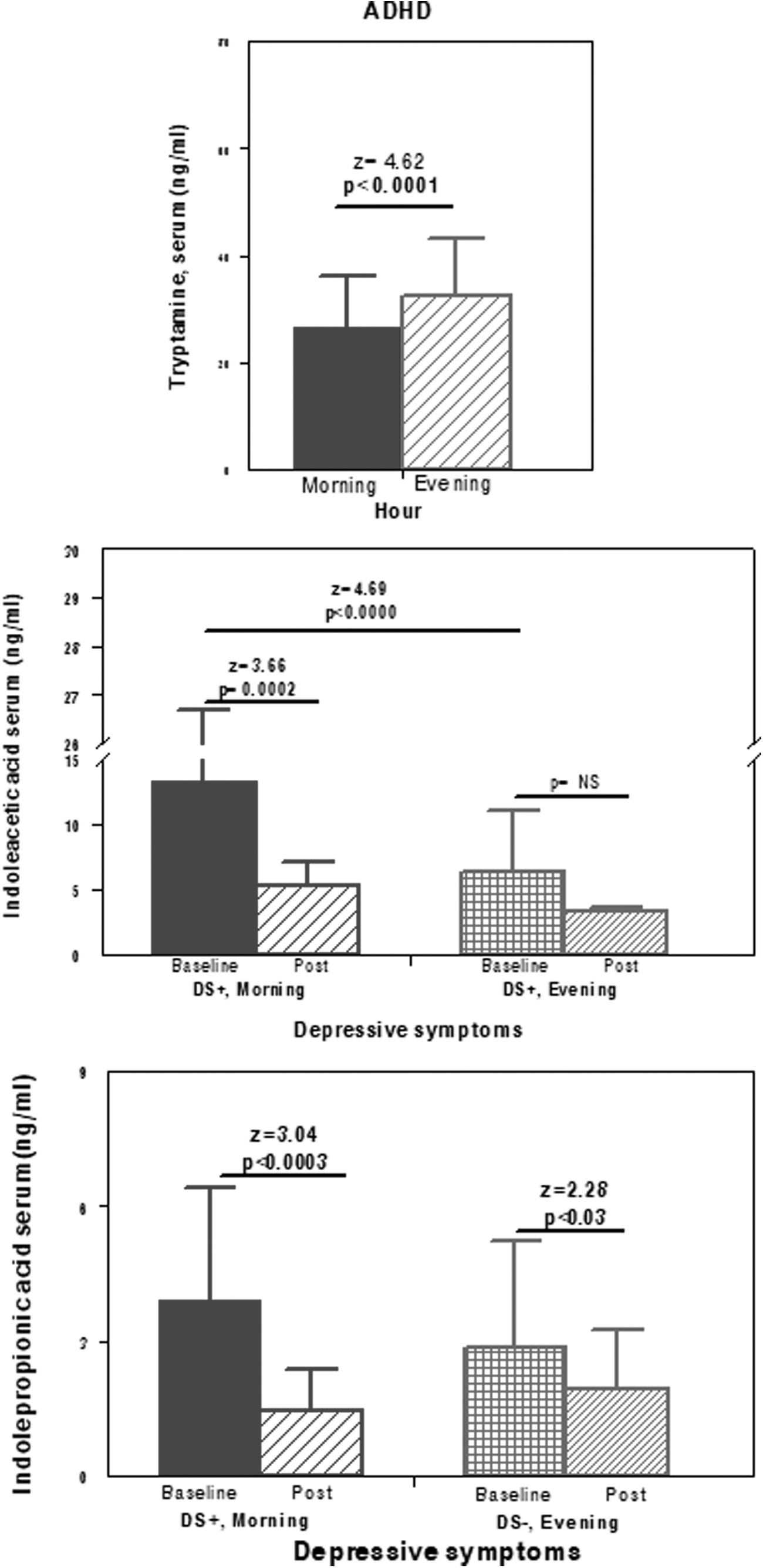
Comparisons of serum concentrations of several measured indoles in the ADHD sample. Top: comparison by hour (morning vs. evening) factor for tryptamine; Middle: comparisons for serum indoleacetic acid in the ADHD subgroup with presence of depressive symptoms between baseline and posttreatment separately during the morning and the evening. Bottom: comparisons between baseline/posttreatment for serum indolepropionic acid separately during the morning and the night. ADHD, attention-deficit/hyperactivity disorder.
Serum Levels of Indols and of Protein S100B Among ADHD Patients With or Without Depressive Symptoms Before and After Treatment with Methylphenidate, and Healthy Controls in the Morning and in the Evening
For indolepropionic acid values after methylphenidate in the whole ADHD: (z = 2.83, p = 0.0047) versus baseline indolepropionic acid.
Morning–evening profile in the whole sample only for tryptamine and S100B: for tryptamine, z = 5.05, p < 0.0001, and for S100B, z = 3.19, p = 0.001.
(z = 4.87, p < 0.0001) versus morning baseline PHI with DS−.
(z = 3.66, p = 0.002) versus morning baseline PHI with DS−.
(z = 2.72, p = 0.007) morning baseline DS− versus morning baseline DS+.
(z = 2.28, p = 0.02) versus evening baseline ADHD.
(z = 3.04, p = 0.002) versus morning baseline ADHD.
Indolepropionic acid morning–evening profile in ADHD with DS+ (z = 2.24; p < 0.03).
Nocturnal 12 Hour Urinary Excretion of Indole Tryptophan Metabolites in the ADHD Groups (Baseline and After Methylphenidate Treatment) and the Control Group
z = 2.47, p = 0.01; versus baseline urinary tryptamine excretion.
Indoleacetic acid
At the baseline measurement, indoleacetic acid (IAA) in the ADHD group had slightly higher morning values that were not significantly different from the evening values. Methylphenidate excretion resulted in a slight reduction in IAA, without significant differences in the morning (6.44 ± 7.10 and 5.17 ± 2.67 ng/mL, baseline and post) or the evening (5.09 ± 4.62 and 4.53 ± 1.49 ng/mL; baseline and post). In the PHI group, before and after treatment, morning IAA values were higher than evening values; in contrast, in the (PAD) group, the evening values were higher and remained elevated after treatment. In both groups, no changes reached significance.
In the PHI DS+ group, the baseline morning IAA was much greater than the evening IAA (z = 4.87, p < 0.0001). Methylphenidate treatment was associated with a 50% decrease in this value (z = 3.66, p = 0.002), resulting in the values nearly matching those observed in the evening, with disappearance of the high variability among individual serum concentrations.
We found no differences in IAA nocturnal urinary excretion between the ADHD subgroups (PAD vs. PHI; Fig. 2), between baseline versus posttreatment, and between DS+ and DS− subgroups (Table 3).
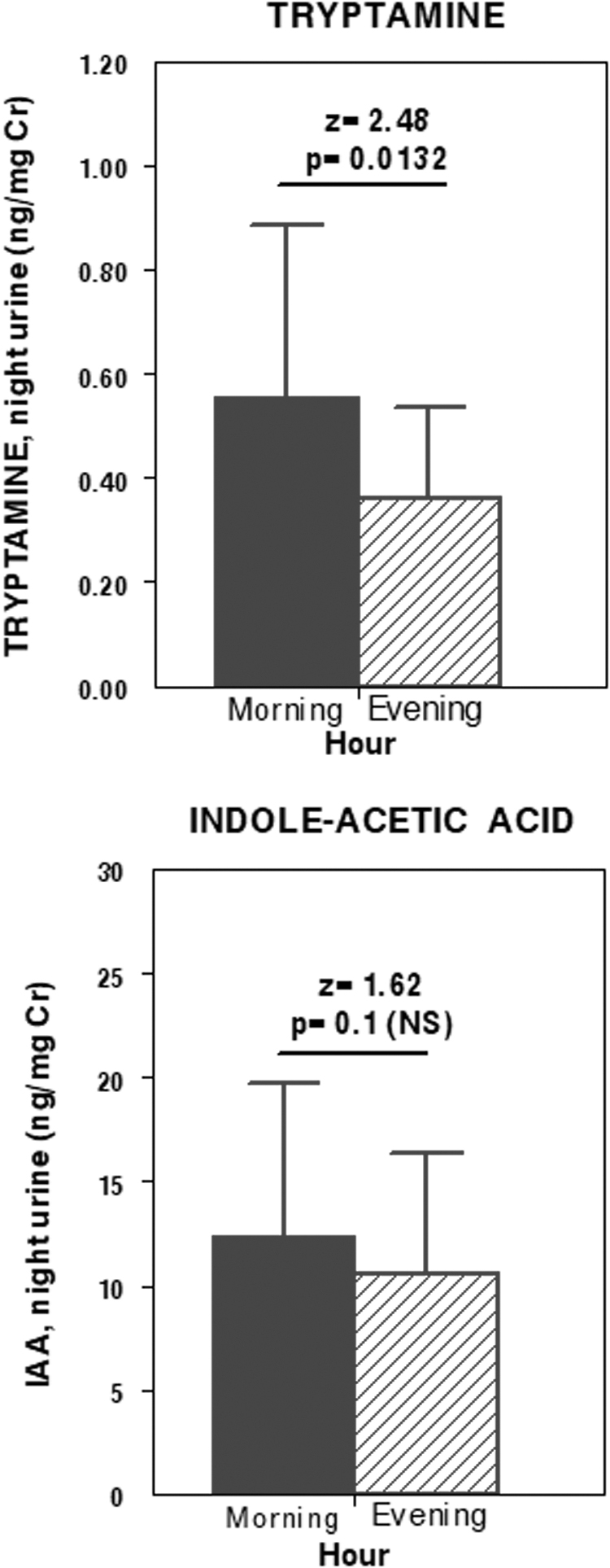
Nocturnal urine excretion of tryptamine (top) and indoleacetic acid (bottom) at baseline (black bars) and after methylphenidate treatment (dashed bars).
Indolepropionic acid
At baseline, IPA (Table 2) was not significantly different between patients and the CG (p = 0.07), and between controls in the morning (1.76 ± 1.67 ng/mL) versus evening (2.35 ± 1.08 ng/mL), and patients in the morning (2.84 ± 2.63 ng/mL) versus evening (2.39 ± 0.95 ng/mL), nonsignificant.
In the ADHD group, methylphenidate was associated with a serum decrease in IPA in all data pairs (2.81 ± 2.41 and 1.96 ± 1.74 ng/mL, baseline and posttreatment; z = 2.83, p = 0.0047). Methylphenidate caused a concentration decrease both in the morning and in the evening until the values reached those observed in the CG, restoring the daily profile (with minimal evening concentration predominance) found in the CG (1.91 ± 1.75 and 1.99 ± 1.75 ng/mL, morning and evening, post). Even posttreatment concentrations were slightly but not significantly higher than those in the CG. Considering the DS factor, methylphenidate also was associated with IPA decrease in all pairs of data, with a decrease that reached significance in the morning concentration, making it similar to the rest of the values in DS+ (z = 3.04, p = 0.002). In contrast, the evening concentration was significant in DS− (z = 2.28, p = 0.02). With respect to the morning/evening profile, the ADHD with DS+ group had a greater morning than evening IPA (z = 2.24; p < 0.03). This difference was erased by methylphenidate, with nocturnal values becoming even slightly higher. The baseline morning IPA was greater in patients with DS+ than DS− (z = 2.72, p = 0.007). Patients with ADHD (PHI and PAD) had a similar IPA concentrations, with higher morning values at baseline (z = 2.38, p < 0.02) in the PHI with SD+ group compared with those in the same subgroup with DS−. After methylphenidate, the IPA concentration tended to decrease in all pairs of data from both ADHD groups, with significant differences in the morning (baseline and post) only in the PHI with DS+ group (z = 2.31, p < 0.03) and at evening in the PAD with DS− group (z = 2.43; p < 0.02).
Protein S100B
In the whole sample, S100B was greater in the morning (49.5 ± 18.1 ng/mL) than in the evening (43.7 ± 11.9 ng/mL; z = 3.19, p = 0.001), with no differences between our CG (Table 2) and ADHD groups/subgroups. In ADHD, methylphenidate was associated with a nonsignificant increase in serum S100B [the lack of differences were probably because of the large spread of the data (large standard deviation)]. The profile of the response to methylphenidate was similar for both ADHD subgroups, with the persistent higher concentrations in the morning. With regard to the comparison of DS, we observed only a trend toward significance that showed a greater concentration in DS+ than in DS− (z = 1.8, p = 0.07).
Discussion
To achieve a better understanding of the neuroendocrine changes that may contribute to the efficacy of methylphenidate treatment in ADHD children, our study is the first to report serum and urinary levels of ITM, in addition to its daily fluctuations in children with ADHD classified into PAD and PHI groups, with subgroups according to comorbid DS, both before and after chronic treatment with methylphenidate. The variables used for sample classification aims to define homogeneous groups and subgroups that allow us the direct comparison of data pairs that differs in such classification factor, for example, the variable labeled “depressive symptomatology.”
Tryptamine
Tryptamine as a trace amine released by neurons (Jones 1982) may couple with dopaminergic and serotonergic neurotransmission, opposing serotonin actions (Mousseau 1993), and increasing monoamine release (Lindemann et al. 2005). The only route of synthesis for brain tryptamine is by direct decarboxylation of
In our study, there were no significant differences in tryptamine between the CG and the ADHD group. In both groups, the nocturnal concentration was higher (34.90 ± 9.27 and 32.65 ± 11.38 ng/mL, with significant morning/evening differences (z = 4.6, p < 0.0001) (Figure 1, top). This profile is not modified by methylphenidate or by the DS factor. Therefore, our data in children are consistent with previous data in healthy adults (Radulovacki et al. 1983) showing an increase in serum tryptamine as the day progresses. In urine, baseline nocturnal tryptamine excretion is very similar in both the CG and ADHD group, with methylphenidate associated with a decrease in the nocturnal excretion of tryptamine (z = 2.47, p = 0.01). A similar profile of decreased excretion by ADHD groups or by DS was also observed. Although methylphenidate did not induce serum changes, it caused a significant decrease in nocturnal excretion, which could be attributed to greater MAO-A/B enzyme activity induced by methylphenidate (Bartl et al. 2017). In addition, β-phenylethylamine, another trace amine, has significantly lower levels in children with ADHD than in controls, but with significantly increased excretion after methylphenidate in responders to treatment but not in nonresponders (Kusaga et al. 2002). In addition, serotonin showed a differential daily profile in the ADHD group compared with that in the CG because of the higher nocturnal concentrations in ADHD (p < 0.034). Methylphenidate corrects these subtle changes in the daily serotonin fluctuation and concentration (Molina-Carballo et al. 2013) and those in melatonin metabolism (Cubero-Millán et al. 2014).
Tryptamine, a trace amine classified as an endogenous amphetamine, increases in concentration along the day perhaps helping to maintain an appropriate alertness. Its serum concentration was not modified by methylphenidate, a stimulant drug that we administered in the morning and also increases alertness. In contrast, methylphenidate was associated with a decrease in the nocturnal urinary excretion of tryptamine that might be a result of being processed through a different metabolic pathway in the service of nightly different physiological needs.
Indoleacetic acid
IAA, a phytohormone, also called indole-3-acetic acid, is produced in mammals from tryptophan in several tissues and in the intestines by enteric bacteria. IAA is originated from oxidative deamination of tryptophan or by decarboxylation of tryptamine, as its main metabolite (Young et al. 1980).
IAA production has been related with ADHD symptoms. Lower plasma IAA (and IPA) concentrations were associated with higher ADHD-like scores in dogs (Puurunen et al. 2016). Gut microbiota production of both IAA and IPA (Maqsood and Stone 2016) modulates the kynurenine pathway and brain-derived neurotrophic factor (BDNF) functions in the CNS. In the absence of GI microbes, reduced central BDNF levels inhibit the maintenance of N-methyl-
In our ADHD sample, the baseline morning IAA was much higher in DS+ than in DS− (z = 5.03, p < 0.0001). Methylphenidate induces a 50% decrease in serum morning IAA in ADHD with DS+ (z = 3.66, p = 0.0002) until these levels virtually match the rest of the measurements and the high data variability disappears (Fig. 1). Alternatively, there were no differences at evening, which did not change after methylphenidate. With our low sample size of ADHD children of PAD subtype with DS+, these data should be applicable to PHI with DS+. The PHI group before and after methylphenidate had greater morning concentrations of IAA (in contrast to tryptamine levels), without differences compared with those at evening. Nocturnal excretion of IAA is similar for both the CG and ADHD group at baseline and was unaltered by methylphenidate or by the DS factor. To our knowledge, no other studies have quantified IAA levels in ADHD children. Urine IAA levels were correlated with anxiety and depression scores in adults with chronic kidney disease (Karu et al. 2016). Moreover, no change is detected in urinary excretion of IAA in patients with depression, with slight increase only in cerebrospinal fluid levels in patients with depression and intellectual disability (Anderson et al. 1984).
Indolepropionic acid
In contrast to tryptamine and IAA, the production of IPA [the most effective hydroxyl radical scavenger (Poeggeler et al. 1999)] is completely dependent on the gut microflora (Wikoff et al. 2009). Dietary tryptophan metabolized by the gut microbiota into IPA (and also to IAA) both as aryl hydrocarbon receptor (AhR) agonists can limit CNS inflammation by acting on astrocytes (Rothhammer et al. 2016).
When the intestinal barrier becomes compromised, the access of both dietary antigens and pathogens is facilitated, eventually leading to innate immune activation with increased cytokine production. IPA confers a neuroprotective effect (Poeggeler et al. 1999; Hwang et al. 2009). The impact of a high-fat diet on brain dopamine metabolism that triggers anxiety-like behaviors and learning/memory impairments in mice may be prevented by methylphenidate (Kaczmarczyk et al. 2013). In experimental ADHD, both IPA and IAA showed negative correlations with ADHD-like behavioral scores, suggesting differences in the intestinal microbiota in dogs (Puurunen et al. 2016). Decreased IPA may predispose individuals to oxidative stress (Zhang and Davies 2016); in addition, stress caused by hyperactive and impulsive behavior may lead to altered gut microbiota with reduced IPA. Probiotics interact with microbial tryptophan metabolism raising the concentration of IPA (Chyan et al. 1999)], and also regulating hypothalamic–pituitary–adrenal (HPA) axis and the immune system (Abildgaard et al. 2017).
Methylphenidate treatment was associated with serum IPA decrease in all comparisons of the ADHD group (z = 2.83, p = 0.0047) and made the profile very similar to that observed in the CG still showing higher but nonsignificant IPA concentrations. The greater decrease occurred in the morning in such a way that the concentrations between morning and evening were no longer different. The higher IPA concentration was measured at baseline in the morning in the ADHD with DS+ group, which showed a highly significant decrease after methylphenidate that equaled the value of the rest of the data pairs. At evening, methylphenidate also decreased IPA, with this decrease reaching statistical significance in the ADHD with DS− group (z = 2.28, p = 0.02). Baseline IPA concentrations were greater in the ADHD with DS+ group (z = 2.72, p = 0.007, and with DS−, morning values), which experienced a similar decrease in response to methylphenidate.
The modulation of tryptophan availability by the gut microbiota also has implications for kynurenine pathway metabolism for the following reasons (Cervenka et al. 2017): (1) Downstream metabolites of kynurenine pathway (KA and quinolinic acid, QA) act as antagonists on NMDA and α7 nicotinic acetylcholine receptors in the CNS and ENS; and (2) microbial metabolites can impact intestinal barrier integrity inducing mucosal inflammation with increased intestinal permeability and subsequent systemic inflammation that could also lead to alterations in kynurenine pathway metabolism (Kelly et al. 2015).
Methylphenidate slightly decreases weight and BMI, which was not related to leptin and BDNF concentrations but most likely to ghrelin and adiponectin. Although posttreatment ghrelin and adiponectin levels were significantly higher in ADHD, BDNF levels were significantly lower (Sahin et al. 2014). In animal models of ADHD, methylphenidate causes an important alteration in energetic metabolism with increased oxidative stress (Comim et al. 2014). These changes are in part owing to dopaminergic changes (Volkow et al. 1997). Energy metabolism in developing children would be facilitated through further production to NAD(+) from tryptophan (Kanchanatawan et al. 2018). Neuronal metabolic responses to stimulation increase direct neuronal glycolysis (Diaz-Garcia et al. 2017). Methylphenidate increases cerebral glucose metabolism in patients with impaired consciousness with an associated improvement in Glasgow Coma Scale scores (Kim et al. 2009). We have reported that methylphenidate modifications to kynurenine pathway metabolites may contribute to marked clinical improvements in the key symptoms of ADHD in children (unpublished results).
In laboratory animals, concentrations of both IAA and IPA show an inverse correlation with anxiety and depression scores. However, among our patients, treatment with methylphenidate was associated with a significant decrease of the morning concentrations of IAA and IPA in the subgroup of patients with DSs. One could interpret that the improvement from DSs owing to methylphenidate would render unnecessary the stimulant properties of IAA and the antioxidant properties attributed to IPA. The strong evidence of a proinflammatory state associated with DSs would support the role of methylphenidate in reducing both DSs and the proinflammatory state, which would render unnecessary buffering by IPA and other molecules.
The apparently paradoxical decrease of IAA and IPA in response to methylphenidate is similar to the paradoxical decrease of the neurotrophin BDNF reported among the same patients (Cubero-Millán et al. 2017).
S100β protein
An earlier study showed no relationship of S100B levels with the core manifestations of ADHD, although its activity shows a trend toward lower levels in patients with anxiety, disturbed mood, and poor self-esteem (Oades et al. 2010). Our data also show no differences in serum S100B in children in the ADHD group with respect to those in the CG. In both groups, there was a higher morning S100B concentration, with highly significant differences (z = 3.19, p = 0.001) with respect to the evening. S100B showed a slight increase after methylphenidate, which may be related to DS+, with a trend toward statistical significance (z = 1.8, p = 0.07). Our data are consistent with previous studies (Oades et al. 2010) in which ADHD seems to be very different from the severe disorders associated with proinflammatory states that give rise to increased S100B secretion. Other measures of CNS inflammation, such as increases in interleukins, IL-16 (along with decreased S100B) and IL-13 in ADHD, have been described in association with total ADHD scores. Most cytokine levels were modestly increased in children with ADHD (not S100B and IL-1β), and all tended to normalize with psychostimulant medication (Oades et al. 2010).
In children with ADHD more so than in controls, the balance between proinflammatory and anti-inflammatory cytokines (such as S100B protein) and potentially toxic or protective kynurenine metabolites may reflect the perinatal experience associated with stress (Oades 2011). Children with ADHD show a reduction in serum levels of kynurenine pathway metabolites [anthranilic acid (AA), KA, and xanthurenic acid (XA)]; in contrast, increased serum levels of tryptophan and kynurenine with unchanged QA levels are observed (Evangelisti et al. 2017). Healthy developing children rather than children with ADHD show higher levels of KA and 3HK (Kanchanatawan et al. 2018). Our data regarding several metabolites of kynurenine pathway are consistent with these observations, corroborating the presence of glutamatergic abnormalities in ADHD (Hammerness et al. 2012), contributing to knowledge regarding a trend toward normalization after methylphenidate (unpublished results).
Prenatal stress, early life stress, and psychological stress, all these factors that has been associated with ADHD, can change the composition of the gut microbiota, with a bidirectional microbial–neuroendocrine relationship (Bailey et al. 2011). The exaggerated HPA axis and stress responses exhibited by Germ-free mice could be normalized in the early weeks of life in a time-dependent manner by probiotics for the correct establishment of the immune system, metabolic system, gut–brain axis, and stress responses (Gareau et al. 2007). In adult ADHD, the physiological levels of peripheral proinflammatory markers, IL-6 and tumor necrosis factor α do not increase (Corominas-Roso et al. 2017). A prebiotic supplement can modulate the magnitude of the cortisol increase within the first 30 minutes after awakening (CAR) (Schmidt et al. 2015).
Indoles such as IPA and IAA, which are generated from tryptophan by the gut microbiota, can limit CNS inflammation (Rothhammer et al. 2016). According to our results, KA increases after methylphenidate (unpublished data) and is accompanied by a slight decline in BDNF levels (Cubero-Millán et al. 2017). Although Tsai proposed blood BDNF and other NTs as ADHD biomarkers by considering NTs as a link between ADHD and other comorbid mental disorders (Tsai 2017), our results for BDNF are consistent with previous studies (Girdler and Klatzkin 2007; Munoz-Hoyos et al. 2011), suggesting that low basal neuroendocrine mediator levels in response to chronic stress and inadequate responses to stimuli are the consequences of repeated biological adaptations to increased life stress; this finding may also be applicable to decreased glucocorticoids with increased proinflammatory signaling (Miller et al. 2009) and to the responses of neurosteroids to methylphenidate in ADHD (Molina-Carballo et al. 2014). The supposed proinflammatory state in patients with ADHD was not reflected in differences in the serum concentration of the proinflammatory cytokine S100B, which was not different in patients compared with healthy controls. In the morning, when the level of activity is higher, the cytokine S100B was significantly higher than in the evening. The progressive physiological exhaustion throughout the day was not associated with an increase in the concentration of the S100B protein that contrariwise decreased. Moreover, the physiological stimulation by methylphenidate was not followed in a significant increase of S100B throughout the day.
As strengths of our study, we should highlight that it is the first study measuring indole metabolites, as objective neuroendocrine modifications, in children with ADHD at baseline and after methylphenidate treatment and the use of a health CG composed mainly of siblings sharing genes, environment and diet regimes with those patients. We recognize the following limitations: (1) a small CG that is largely composed of the siblings of the patients; (2) a small number of females, adolescents and patients belonging to the PAD group; and (3) a proportion of PHI ADHD children with comorbid CD. Those possible confounders of biological measures were not controlled, beyond recommending to patients to follow-up a standard diet several days before analytical extractions. To control possible confounders of biological measures we included sibling of the patients in the CG, despite the well-known high genetic load of ADHD. Thus, the numerical differences between study groups in the biological variables measured could have been larger if the CG had not included siblings of the patients. All subjects live in the same geographical area and have a medium-low socioeconomic level.
Conclusions
Our results suggest that modifications of several indole tryptophan metabolites may participate in the highly favorable clinical response to methylphenidate was associated with modifications may be considered protective based on our knowledge about the physiological functions of the quantified metabolites. In addition, our study highlights the relevance of DSs on the variations in the measured parameters.
Clinical Significance
Prenatal stress, early life stress, and psychological stress can change the composition of the gut microbiota, with a bidirectional microbial–neuroendocrine relationship. Indoles such as indolepropionic acid and indoleacetic acid, which are generated from tryptophan by the gut microbiota, can limit CNS inflammation.
Changes in plasma levels of indole tryptophan metabolites were mainly related to depressive symptoms and achieved a similar profile to controls under methylphenidate treatment.
ADHD seems to be different from the severe psychiatric disorders associated with proinflammatory states that give rise to increased S100B secretion.
Footnotes
Authors' Contributors
A.M.-C., J.U., and A.M.-H. designed the study and wrote the protocol. L.F.-L., A.C.-R., I.C.-M., I.M.-C., E.B.-J., and A.J.-C. made the sample collection and managed the literature searches and analyses. A.M.-C. and A.M.-H. undertook the statistical analysis; I.C.-M. and A.M.-C. wrote the first draft of the article. All researchers contributed and have approved the final version of the article.
Acknowledgments
The authors thank Juan de Dios Luna del Castillo, from the Departamento de BioEstadística. Facultad de Medicina, Universidad de Granada, for his statistical support. The authors also thank Puri Ubago-Corpas for her whole supervision and support.
Disclosures
No competing financial interests exist.