
Abstract
Background:
Vascular endothelial growth factor (VEGF) may be relevant to bipolar disorder (BD) and brain structure. We evaluated VEGF rs699947 single-nucleotide polymorphism in relation to structural neuroimaging phenotypes in youth BD.
Methods:
We collected 3 T anatomical magnetic resonance images from 154 youth (79 BD and 75 healthy control [HC]) genotyped for VEGF rs699947. The participants were age (BD = 17.28 ± 1.40 and HC = 17.01 ± 1.83, t = −1.02, p = 0.31) and sex (BD = 63.3% females and HC = 52.0% females, χ 2 = 2.01, p = 0.16) matched. Cortical thickness, surface area (SA), and volume were examined by region-of-interest (ROI) and vertex-wise analyses using general linear models (GLMs). ROI investigations selected for the prefrontal cortex (PFC), amygdala, and hippocampus. Vertex-wise analyses controlled for age, sex, and intracranial volume.
Results:
ROI results found lower PFC SA (p = 0.003, ηp 2 = 0.06) and volume (p = 0.04, ηp 2 = 0.03) in BD and a main effect of rs699947 on hippocampal volume (p = 0.03, ηp 2 = 0.05). The latter two findings did not survive multiple comparisons. Vertex-wise analyses found rs699947 main effects on left postcentral gyrus volume (p < 0.001), right rostral anterior cingulate SA (p = 0.004), and right superior temporal gyrus thickness (p = 0.004). There were significant diagnosis-by-genotype interactions in the left superior temporal, left caudal middle frontal, left superior frontal, right fusiform, and right lingual gyri, and the left insular cortex. Posthoc analyses revealed the AA allele was associated with larger brain structures among HC, but smaller brain structures in BD for most clusters.
Conclusions:
Overall, we found preliminary evidence of divergent associations between BD and HC youth in terms of neurostructural correlates of VEGF rs699947 encompassing highly relevant frontotemporal regions.
Introduction
Bipolar disorder (BD) is a severe mood disorder affecting 2%–5% of adolescents (Goldstein et al., 2017; Van Meter et al., 2019). In addition to the mood symptoms of depression and mania that define the condition, individuals with BD are at increased risk for multiple psychiatric and medical comorbidities (Goldstein et al., 2017). Strong associations between BD and vascular disease have been reported, independent of other cardiovascular risk factors such as smoking, hypertension, obesity, and drug use; international studies reported earlier onset, greater risk of mortality, and increased risk factors for cardiovascular disease (CVD) (Goldstein et al., 2020; Goldstein et al., 2015).
While links to vascular disease have been reported in several other psychiatric disorders, the association is among the strongest in BD (Goldstein et al., 2020; Nielsen et al., 2021). There are multiple factors that contribute to the association of BD with increased risk and premature onset of CVD. BD is associated with adverse health-related behaviors such as smoking, sedentary lifestyle, suboptimal nutrition, and substance use disorder (SUD) (Goldstein et al., 2020). In addition, BD is associated with increased rates of traditional vascular risk factors such as obesity and hypertension.
However, there is evidence, including largely untreated epidemiologic samples, that the vascular-bipolar link exceeds what can be explained by factors such as lifestyle, medications, and even by traditional vascular risk factors (Goldstein et al., 2020; Goldstein et al., 2015). Given that BD typically onsets in adolescence, whereas CVD typically onsets in mid- and late life, intermediate vascular phenotypes and biomarkers could help to inform our understanding of the vascular-bipolar link before CVD and other age-related disorders fully manifest.
The progression of BD involves a complex interplay between polygenic factors and environmental stressors. BD pathophysiology tends to draw about inflammation, oxidative stress, and impaired neurotrophic support as potential mechanisms (Berk et al., 2011; Goldstein et al., 2017; Scola and Andreazza, 2015). These processes are known to influence CVD development and may serve to bridge the relationship between vascular disease and BD (Goldstein et al., 2020). Vascular endothelial growth factor (VEGF) acts on blood vessels and neurons; it has been studied extensively in angiogenesis and implicated in atherosclerosis and chronic inflammatory states (Braile et al., 2020; Howell et al., 2005; Zhao et al., 2018). In the brain, VEGF acts as a neurotrophic factor influencing neuronal formation, plasticity, migration, and survival in physiological and pathological settings, including psychiatric disorders (Mackenzie and Ruhrberg, 2012; Scola and Andreazza, 2015).
Differential VEGF protein or mRNA levels have been reported in disease groups from multiple psychiatric conditions/states, including schizophrenia, major depressive disorder (MDD), suicidality, anxiety, and attention-deficit/hyperactivity disorder (Becerril-Villanueva et al., 2019; Isung et al., 2012; Pu et al., 2020; Sundberg et al., 2020; Yurteri et al., 2019). In terms of BD, previous studies have shown elevated peripheral VEGF protein concentrations in adults with BD during depressive and manic episodes compared to controls, and elevated VEGF in adults with BD during symptomatic mood states compared to euthymic states (Pu et al., 2020).
Similarly, there is evidence of increased VEGF mRNA during unipolar and bipolar depression (Shibata et al., 2013), and decreased VEGF mRNA following lithium treatment in BD patients (Kikuchi et al., 2011). However, findings across studies have been inconsistent, with several reports showing no significant correlation between peripheral VEGF levels and BD (Pu et al., 2020; Tuncel et al., 2020; van den Ameele et al., 2017).
BD is highly heritable, although the genetic determinants are not fully understood (Berk et al., 2011). Given the roles of VEGF in the brain and its differential expression in brain regions involved in BD, genetic variants in VEGF may be involved in neurostructural differences in BD. A functional VEGF variant, rs699947 (−2578 C/A), located in the promoter region of VEGF, has been associated with VEGF serum levels (Shahbazi et al., 2002; Steffensen et al., 2010) and conditions including treatment-resistant MDD (Viikki et al., 2010) and vascular disease (Howell et al., 2005; Zhao et al., 2018). Furthermore, rs699947 was identified as a variant of interest by the Psychiatric Genome Consortium (PGC) BD and Schizophrenia groups.
While the PGC BD group did not find a significant risk association with rs699947, the PGC Schizophrenia group reported a significant difference in rs699947 between patients with schizophrenia versus controls (p < 0.003, OR = 0.97–1.04), which did not reach genome-wide significance (p < 5 × 10−8) (Consortium, 2014; Group, 2011; Lizano et al., 2018).
Notably, these genome-wide association studies were conducted with mainly adults of European descent. Given that early-onset BD is associated with increased familial load, genetic variation may exert greater effects on youth BD (Faraone et al., 2003). Moreover, VEGF rs699947 has been implicated in neurostructural differences, such as an increase in total gray and white matter volume (Takeuchi et al., 2017) and larger hippocampal volume increase following electroconvulsive therapy in C-allele carriers in a dose-dependent manner (Van Den Bossche et al., 2019). Therefore, rs699947 may be relevant to brain structure in BD youth.
The objective of this study was to investigate the association of VEGF rs699947 on neurostructural phenotypes in a relatively large, controlled sample of youth with BD. We selected the prefrontal cortex (PFC), amygdala, and hippocampus as regions of interest (ROIs) based on their relevance to BD. In addition, we performed whole-brain vertex-wise analyses to complement ROI analyses. While there is not sufficient literature to inform directionally specific hypotheses in BD and/or in youth, we expected that the rs699947 genotype would be associated with brain structure, particularly in frontal and temporal regions.
Experimental Procedures
Participants
A total of 154 participants 13 to 20 years of age were recruited for this study. Seventy-nine youth with BD (BD-I, BD-II, or BD-not otherwise specified [NOS]) were recruited from the Centre for Youth with Bipolar Disorder, a subspecialty clinic at the Centre for Addiction and Mental Health (CAMH) in Toronto, Canada. Seventy-five healthy controls (HCs) who had no lifetime mood or psychotic disorder, no first- or second-degree family history of BD or psychotic disorders, and no anxiety disorders within the past 3 months were recruited from the community.
BD and HC groups were age (BD = 17.28 ± 1.40 and HC = 17.01 ± 1.83, t = −1.02, p = 0.31) and sex (BD = 63.3% females and HC = 52.0% females, χ 2 = 2.01, p = 0.16) matched. Exclusion criteria for the study were as follows: (1) unable to provide informed consent; (2) infectious illness within the past 14 days; (3) existing cardiac condition, autoimmune illness, or inflammatory illness; (4) currently taking anti-inflammatory, antiplatelet, antilipidemic, antihypertensive, or hypoglycemic agents; (5) magnetic resonance imaging (MRI) contraindications; (6) history of severe neurological or cognitive impairment; and (7) substance dependence in the past 3 months (nondependent substance abuse was permitted). Approval was granted by the institutional research ethics board and written informed consent was provided by participants and at least one parent/guardian before beginning the study.
Assessments
Assessments were conducted by research staff trained under the senior author (B.I.G., a licensed child-adolescent psychiatrist) and had Bachelor's and/or Master's degrees in health sciences. Participant interviews, parent/guardian interviews, medical records, and all other available information were used to diagnose participants. Current and lifetime diagnoses of BD and other psychiatric conditions were determined using the Kiddie Schedule for Affective Disorders and Schizophrenia for School Age Children, Present and Lifetime (K-SADS-PL) (Kaufman et al., 1997).
Given that participant recruitment and interviews were conducted from 2012 to 2019, Diagnostic and Statistical Manual of Mental Disorders IV (DSM-IV) criteria were used to evaluate participant and parent/guardian interviews (K-SADS-PL version DSM-5 was not available until December 2016). The Course and Outcome of Bipolar Illness in Youth study was used to define criteria for BD-NOS (Birmaher et al., 2009). Age of BD onset was defined as the age when the first episode of mania or hypomania (according to DSM-IV) was experienced, or when criteria for BD-NOS were met. Following the K-SADS-PL interviews, diagnoses were confirmed in a consensus meeting with a licensed child-adolescent psychiatrist.
Socioeconomic status was assessed using the four-factor Hollingshead Scale (Hollingshead, 1975). Body mass index (BMI) was calculated as weight (kg)/height (m)2. Weight was adjusted for clothing (−0.9 kg for short pants/short shirt, −1.1 kg for short pants or short shirt, and −1.3 kg for long pants/long shirt). Physical and/or sexual abuse and suicidality or nonsuicidal self-injury were recorded using a safety form. Extended K-SADS Depression Rating Scale (Chambers et al., 1985) and K-SADS Mania Rating Scale (Axelson et al., 2003) were used in lieu of the K-SADS-PL mood sections. The Children's Global Assessment Scale was used to evaluate global functioning throughout the most severe past episode, highest level in the past year, and current period (past month), with higher scores reflecting higher functioning (Shaffer et al., 1983).
K-SADS-PL was used to evaluate comorbid diagnoses and clinical characteristics (e.g., psychosis, psychotropic and psychosocial treatment history). The “Any Anxiety” variable included any anxiety disorder (i.e., generalized anxiety disorder, social phobia, separation anxiety disorder, agoraphobia, panic disorder, and anxiety disorder-NOS). SUD included alcohol or drug abuse or dependence. A Family History Screen (Weissman et al., 2000) from the participant parent/guardian(s) was used to evaluate the psychiatric history of first- and second-degree relatives.
Genotyping
Saliva samples were collected in Oragene DNA saliva kits from participants and genotyped for the VEGFA (henceforth referred to as VEGF) rs699947 marker at nucleotide −2578 in the VEGF gene promoter region. Briefly, total genomic DNA was extracted from the saliva using the standard protocol on a Magnetic Separation Module I automated nucleic acid extractor (Perkin-Elmer, Waltham, MA). The DNA was genotyped for the rs699947 marker using TaqMan SNP Genotyping Assay C___8311602_10 (Life Technologies, Applied Biosystems, Inc., Burlington, ON, Canada) per the manufacturer's instructions on the QuantStudio 12K Flex PCR System (Life Technologies, Applied Biosystems, Inc., Carlsbad, CA, USA). Genotyping of 10% of samples from each run was replicated for quality control purposes. All genetic sample processing (DNA extraction and genotyping) was performed by the Centre for Addiction and Mental Health (CAMH) Biobank and Molecular Core Facility.
MRI and processing
A 3 T Philips Achieva scanner (using an eight-channel head coil) was used to acquire T1-weighted high-resolution fast-echo images (repetition time/echo time/inversion time = 9.5/2.3/1400 milliseconds; spatial resolution = 0.94 × 1.17 × 1.2 mm, 256 × 164 × 140 matrix, flip angle = 8°, field of view = 240 × 191 mm, scan duration = 8 minutes 56 seconds, 140 slices). Voxel size was 1 mm isotropic.
T1-weighted images were processed using FreeSurfer v6.0 (Fischl, 2012) to calculate cortical volume, thickness, and surface area (SA). Preprocessing steps included automated skull stripping (Ségonne et al., 2004), intensity normalization (Sled et al., 1998), Talairach transformation, reconstructing the cortical surface, and white and gray matter segmentation (Fischl et al., 2004). A white matter surface mesh was generated from binary white matter masks, which was then expanded to a gray matter mesh corrected for voxel-wise and topological effects (Fischl et al., 2001).
The MRI data were further processed, surfaces were inflated, and then registered to a spherical atlas (Fischl et al., 1999). A parcellation algorithm was applied based on gyral and sulcal features, and then mapped onto the Desikan-Killiany-Tourville atlas. Then, three ROIs were generated: (1) PFC consisting of the bilateral pars opercularis, pars orbitalis, pars triangularis, rostral middle frontal, caudal middle frontal, and superior frontal gyri, and the lateral orbitofrontal cortex, medial orbitofrontal cortex, rostral anterior cingulate, caudal anterior cingulate, and frontal pole; (2) bilateral amygdala; and (3) bilateral hippocampus. Volume and SA were defined as the sum of all annotations in each ROI, and thickness was determined proportional to SA. A 10 mm isotropic kernel was used to smooth brain surfaces of participants' brains to minimize voxel-wise effects and increase signal-to-noise ratio.
Parcellation quality control was conducted by three independent raters. Three trained raters examined the images for motion artifacts, gross structural abnormalities, and other indices of image quality, and then assigned a score between 0 and 3 on the basis of overall quality, including motion artifacts during scan, and contrast between gray and white matter. Images with a score of 3 (poor quality) were excluded from preprocessing and analyses. Images with poor parcellation accuracy were manually edited or excluded from the analyses. Six participants failed quality control (two BD-CC, one BD-AC, one HC-AA, and two unknown).
Statistical analyses
All statistical analyses were conducted with Statistical Package for the Social Sciences, version 27. The demographics and clinical variables for sample groups were analyzed using independent sample t-test or one-way ANOVA for continuous variables, χ 2 test for categorical variables, and Mann-Whitney or Kruskal-Wallis tests for ordinal variables. Tukey's honest significant difference (HSD) was used for posthoc analysis of group differences. Welch test followed by Games-Howell posthoc test was used if homogeneity of variance was violated. Effect sizes were Cramer's V, Cohen's d, or partial eta squared (ηp 2). For effect sizes reported as Cramer's V, magnitude of effect size is estimated with the following guidelines: Cramer's V < 0.02 indicating a small effect, Cramer's V = 0.02–0.06 indicating a medium effect, and Cramer's V > 0.06 indicating a large effect.
The degree of freedom is 2 [χ 2 (2, N = SAMPLE SIZE) = chi-squared value, p = p value]. For effect sizes reported as partial eta squared (ηp 2), magnitude of effect size is estimated with the following guidelines: ηp 2 = 0.01 indicating a small effect, ηp 2 = 0.06 indicating a medium effect, and ηp 2 = 0.14 indicating a large effect (Cohen, 1988). A sensitivity power analysis was conducted based on the sample size. Setting alpha to 0.05 and assuming 80% power, this study was powered to detect small effect sizes (f = 0.23).
Analyses for both ROI and vertex-wise analyses were performed using a GLM design covariance analysis matrix with discrete (diagnosis and VEGF rs699947) and nuisance (age, sex, race, and intracranial volume [ICV]) covariates inputted. Race was coded as Caucasian versus non-Caucasian. ICV was measured as a continuous variable. ICV was excluded as a covariate for measures of cortical thickness. Shapiro-Wilk tests were performed to test the assumption of normality for covariates (age, ICV) before including them into the statistical model. Bonferroni correction was used to correct for multiple comparisons in ROI analyses (p < 0.017).
Vertex-wise analyses were performed using a shell script, which mapped participant cortical measurements to an average control surface. Then in each hemisphere, volume, SA, and thickness measurements were contrasted from vertex to vertex across the cortex. Finally, multiple comparisons were corrected by permutation testing using cluster-forming and cluster-wise thresholds of p < 0.05. Significant clusters from the vertex-wise analyses were then extracted, covariate adjusted using the aforementioned GLMs, and then a one-way ANOVA with posthoc testing corrected for multiple comparisons using Tukey's HSD examined for between-group differences.
Parallel sensitivity analyses were conducted to assess race, current lithium use, and current second-generation antipsychotic (SGA) use. Additional sensitivity analyses assessing for BMI, lifetime SUD, current substance abuse, BD subtypes, and time since diagnosis were conducted for ROI analyses.
Results
Demographic and clinical characteristics
Participant demographic and clinical characteristics are outlined in Tables 1 and 2. The genotypic distribution was 20.8% AA (n = 32), 53.2% AC (n = 82), and 26.0% CC (n = 40). The polymorphism was in Hardy-Weinberg equilibrium in the overall sample, as well as in BD and HC groups. Six genotype subgroups were considered in this study (i.e., AA, AC, and CC genotypes in BD and HC groups). The BD group had significantly more Caucasian participants (p = 0.04, V = 0.17) and higher BMI (p = 0.003) relative to the HC group. Within BD, significant genotypic differences were found for lifetime SGA use (p = 0.02, V = 0.31). After pairwise posthoc multiple comparisons tests, the BD-AC group had significantly greater lifetime SGA use compared to the BD-CC group (p = 0.007, V = 0.35).
Demographics of Study Participants
Values for all continuous variables are written as mean ± SD and categorical variables are written as n (% within group). Independent sample t-test or one-way ANOVA was used for continuous variables, χ 2 tests were used for categorical variables, and Mann-Whitney or Kruskal-Wallis tests were used for ordinal variables. Welch test followed by Games-Howell posthoc test was used if homogeneity of variance was violated.
Significant main effect of BD diagnosis (p < 0.05).
BD, bipolar disorder; BMI, body mass index; BP, blood pressure; HC, healthy control; n, number of participants; SD, standard deviation; VEGF, vascular endothelial growth factor.
Clinical Characteristics of Study Participants
Values for all continuous variables are written as mean ± SD and categorical variables are written as n (% within group). Independent sample t-test or one-way ANOVA was used for continuous variables and χ 2 tests were used for categorical variables. Welch test followed by Games-Howell posthoc test was used if homogeneity of variance was violated.
Significant BD diagnosis-by-VEGF rs699947 interaction (p < 0.05).
ADHD, attention-deficit/hyperactivity disorder; BD, bipolar disorder; CGAS, Children's Global Assessment Scale; HC, healthy control; n, number of participants; NOS, not otherwise specified; ODD, oppositional defiant disorder; SD, standard deviation; SGA, second-generation antipsychotic; SSRI, selective serotonin reuptake inhibitor; VEGF, vascular endothelial growth factor.
ROI analyses
There was a diagnosis main effect on PFC, where the BD group had smaller volume (p = 0.04, ηp 2 = 0.03) and SA (p = 0.003, ηp 2 = 0.06) compared to the HC group. There was also a main effect of VEGF rs699947 on hippocampal volume (p = 0.03, ηp 2 = 0.05; Supplementary Table S1). The diagnosis effect on PFC volume and the genetic effect on hippocampal volume were not significant after corrections for multiple comparisons. No significant genotypic difference was found for PFC or amygdala ROIs. Diagnosis-by-genotype interactions were not significant for any ROI. To further explore the VEGF rs699947 main effect finding, exploratory posthoc pairwise contrasts between each group were undertaken for the interaction effect on hippocampal volume. Hippocampal volumes in the BD-AA group were significantly smaller than the HC-AA (p = 0.007), HC-AC (p = 0.02), and BD-AC (p = 0.004) groups (Supplementary Fig. S1).
Vertex-wise analyses
Vertex-wise analyses revealed significant clusters with main effects of VEGF rs699947 and diagnosis-by-VEGF rs699947 interactions. Significant peak clusters and their associated additional regions are summarized in Table 3 and Supplementary Table S2. Mean values, mean differences, estimates of uncertainty, and regression coefficients for significant clusters in VEGF rs699947 and diagnosis-by-VEGF rs699947 are outlined in Supplementary Tables S3–S7. A full list of significant posthoc analyses for VEGF rs699947 main effects and diagnosis-by-VEGF rs699947 interaction effects are presented in Table 4.
Significant Cluster Results from Vertex-Wise Analyses
MNI, Montreal Neurological Institute; VEGF, vascular endothelial growth factor.
Posthoc Multiple Comparisons of Significant Clusters from Vertex-Wise Analyses
BD, bipolar disorder; HC, healthy control; HSD, honest significant difference; SA, surface area; VEGF, vascular endothelial growth factor.
VEGF rs699947 main effects
The AC group had significantly greater left postcentral gyrus volume compared to the AA (p < 0.001) and CC (p < 0.001) groups (Supplementary Fig. S2A, B). In addition, the right rostral anterior cingulate SA (extending into the medial orbitofrontal cortex) was significantly greater in the AC group compared to both the AA (p = 0.03) and CC (p = 0.01) groups (Supplementary Fig. S2A, B). Finally, the AC group had significantly smaller right superior temporal gyrus thickness (extending into the insular cortex and lateral orbitofrontal cortex) compared to the AA group (p = 0.003) (Supplementary Fig. S2C).
Diagnosis-by-VEGF rs699947 interactions
Significant clusters were identified in the (1) Dorsolateral PFC: Left superior frontal gyrus thickness extending into the caudal middle frontal gyrus, and left caudal middle frontal gyrus volume extending into the rostral middle frontal gyrus; (2) Insular cortex: Left insular cortex thickness and volume extending into the temporal gyrus, and precentral and postcentral gyri, respectively; (3) Temporo-occipital regions: Left superior temporal gyrus volume, right fusiform and lingual gyri volume, and SA extending into the lateral occipital lobe (Fig. 1 and Supplementary Figs. S3 and S4).
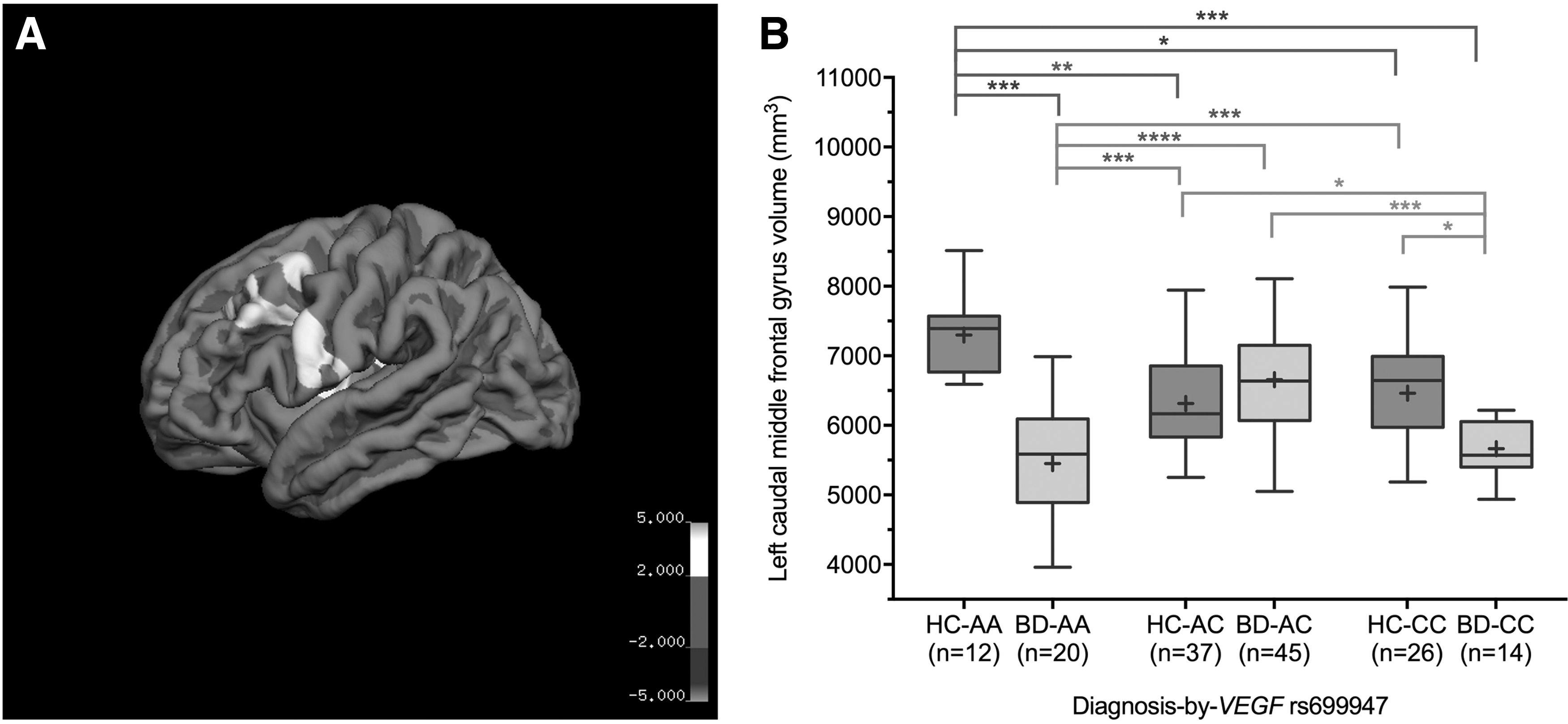
Significant diagnosis-by-VEGF rs699947 interactions from vertex-wise analyses. The rendered brain map shows a significant cluster in left caudal middle frontal gyrus
In posthoc analyses, the BD-AA group consistently had significantly smaller cortical measures in the aforementioned clusters compared to HC groups (HC-AA, HC-AC, and HC-CC) and the BD-AC group (Fig. 1 and Supplementary Figs. S3 and S4). In contrast, the HC-AA group consistently had significantly larger cortical measures in the identified clusters compared to all other groups, except for left superior frontal gyrus thickness relative to HC-AC, HC-CC, and BD-AC groups (Fig. 1 and Supplementary Figs. S3 and S4). Left caudal middle frontal gyrus volume was significantly lower in the BD-CC group compared to all HC groups and the BD-AC group (Fig. 1).
Sensitivity analyses
All ROI and vertex-wise results remained significant after performing sensitivity analyses controlling for race, current lithium use, and current SGA use. ROI results remained significant after controlling for BMI, lifetime SUD, current substance abuse, BD subtypes, and time since diagnosis.
Discussion
The objective of this study was to investigate the neurostructural implications of VEGF rs699947 in youth with BD. In ROI analyses, we found a main effect of VEGF rs699947 on hippocampal volume, which did not survive correction for multiple comparisons. We observed significant VEGF rs699947 main effects in vertex-wise analyses, whereby left postcentral gyrus volume and right rostral anterior cingulate SA were significantly greater in AC genotype group compared to other genotype groups, and right superior temporal gyrus thickness was significantly smaller in the AC group compared to the AA group.
Vertex-wise analyses also identified significant diagnosis-by-VEGF rs699947 interactions in areas within the dorsolateral PFC, insular cortex, and temporo-occipital regions known to be relevant to BD. Posthoc comparisons revealed divergent associations for the AA homozygotes: In HC, the AA genotype was associated with larger structures, whereas in BD, the AA genotype was associated with smaller structures. Results remained significant after sensitivity analyses controlling for current lithium and current SGA use. Of primary interest, these findings provide preliminary evidence of differential associations of VEGF rs699947 with neurostructure in youth BD across the brain, particularly frontal and temporal regions.
The impact of VEGF rs699947 on brain structure and mood disorders has not been fully elucidated. This study presents evidence of a divergent relationship between the HC-AA group (largest) and BD-AA group (smallest) regarding brain structure in several regions associated with BD. Given that genetic and environmental factors likely jointly contribute to brain structure in psychiatric disorders, these findings may be associated with discrepant gene-environment interactions (Boyce, 2016). Several models have been proposed to explain gene-environment interactions, including the diathesis stress, vantage sensitivity, and differential susceptibility models (Boyce, 2016; Jolicoeur-Martineau et al., 2019). One possible explanation for our results is the differential susceptibility perspective, whereby a given genotype confers differential susceptibility to negative versus positive stressors in a bidirectional manner (Jolicoeur-Martineau et al., 2019).
Individuals with BD are more likely to experience multiple negative stressors compared to healthy individuals (e.g., psychiatric symptoms, familial psychopathology, abuse, negative life events) (Kapczinski et al., 2008), which may relate to disproportionate detrimental effects on brain structure within the BD-AA group. Contrastingly, a positive environment may be associated with beneficial brain effects for AA genotype-carrying healthy youth.
VEGF signaling is complex, varies depending on cell type and upstream stimulators, and affects multiple processes, including neurotrophy, neurogenesis, and synaptic plasticity (Mackenzie and Ruhrberg, 2012; Scola and Andreazza, 2015). Further, VEGF stimulates expression of brain-derived neurotrophic factor (BDNF), which is known to influence brain structure and the mechanism of affective disorders (Kennedy et al., 2021; Legge et al., 2015; Mackenzie and Ruhrberg, 2012; Matsuo et al., 2009; Scola and Andreazza, 2015).
Beyond neurotrophic functions, VEGF also affects vascular processes such as angiogenesis and vascular permeability leading to inflammation (Braile et al., 2020). Indeed, enhanced levels of VEGF have been reported in inflammatory conditions (Braile et al., 2020; Pu et al., 2020), and several conditions rely on anti-VEGF treatments to reduce the severity of vascular, proliferative, and inflammatory responses (Hegde et al., 2018). Thus, we speculate that differential effects of VEGF on inflammatory and neurotrophic factors, and/or differential function of VEGF when exposed to these factors, may contribute to the interaction findings.
The main effect of diagnosis in the AA genotype in vertex-wise analyses was evident in regions relevant to BD: (1) Dorsolateral PFC (left superior frontal and left caudal middle frontal gyri), which plays an important role in executive functions (e.g., emotional regulation, impulse control, working memory), is heavily implicated in mood disorders (Hibar et al., 2018; Roybal et al., 2012), and has previously been associated with decreased VEGF mRNA in a postmortem brain study of schizophrenia (Fulzele and Pillai, 2009); (2) Insular cortex, which is involved in the processing of internal cognitive, affective, and somatic perceptions, and integration of those perceptions into decision making (Wise et al., 2017); and (3) Temporo-occipital regions (left superior temporal, right fusiform, and right lingual gyri), which regulate functions, including communication, facial recognition, and emotional processing (Hibar et al., 2018; Roybal et al., 2012; Wiggins et al., 2017; Wise et al., 2017).
Vertex-wise analyses also showed significantly lower left caudal middle frontal gyrus volume in the BD-CC group compared to the HC-AA, HC-AC, HC-CC, and BD-AC groups. Taken together, these results support our hypothesis that VEGF rs699947 is relevant to neurostructural phenotypes in BD, and may be associated with VEGF differential expression and multifunctionality in vascular and neurological processes (Mackenzie and Ruhrberg, 2012).
Our ROI analyses revealed a significant main effect of VEGF rs699947 for hippocampal volume, which did not survive posthoc corrections. The hippocampus is heavily involved in learning and memory, with volumetric and cognitive deficits found in BD (Otten and Meeter, 2015). A previous study of older patients with MDD showed that rs699947 is associated with hippocampal volume changes, possibly through impaired neuroplasticity and neurogenesis (Van Den Bossche et al., 2019). Furthermore, previous preclinical studies using rat models of depression or chronic stress have reported differences in VEGF protein and/or mRNA in the hippocampus, PFC, amygdala, and/or hypothalamus (Elfving et al., 2010; Nowacka-Chmielewska et al., 2017). In our study, exploratory posthoc analyses of group differences in the diagnosis-by-VEGF rs699947 interaction for hippocampal volume showed that BD-AA group had significantly smaller volume than the HC-AA, HC-AC, and BD-AC groups, corroborating the pattern we observed for the BD-AA group in our vertex-wise analyses.
There were also VEGF genotype and diagnosis main effects. We observed a VEGF rs699947 main effect where the AC group had greater cortical measures compared to the AA and CC groups in left postcentral gyrus volume and right rostral anterior cingulate SA, while right superior temporal gyrus thickness was significantly smaller in the AC group compared to the AA group. The superior temporal gyrus and anterior cingulate cortex are associated with BD, and the anterior cingulate cortex in particular has been strongly associated with affective regulation (Hibar et al., 2018; Roybal et al., 2012; Wise et al., 2017). Furthermore, our vertex-wise cluster regions for the right superior temporal gyrus and rostral anterior cingulate extended into the orbitofrontal cortex, a region heavily implicated in impulse and mood control, as well as reward behavior, with differential volumes found in youth BD compared to HCs (Najt et al., 2007). Given that these regions have been implicated in BD, main effects of VEGF in these areas may be related to youth with BD.
Prior studies have found regional differences in youth brain structure depending on age and gene effects (Hoeft et al., 2010; Parker et al., 2020). Furthermore, the angiogenic and neurotrophic properties of VEGF can both affect neurodevelopmental processes (Mackenzie and Ruhrberg, 2012). Therefore, the VEGF rs699947 genotype may be associated with differences in brain structure relating to spatiotemporal influences. Finally, consistent with prior studies in adults, ROI analyses showed that BD youth had smaller PFC volume and SA compared to controls (Abé et al., 2016).
Limitations
There are several limitations to this study. First, although a comparatively large sample within the youth BD neuroimaging literature, this study was not powered to detect small effect sizes beyond f = 0.23. This relates both to the neuroimaging findings and the clinical correlates of VEGF rs699947 within BD. Similarly, we included a heterogeneous sample reflective of the epidemiology of BD, yielding variability in BD subtypes, family history, comorbidity, and treatment. Much larger samples, likely requiring multisite initiatives, will be required to examine for small effect sizes and differences related to clinical and familial variability. In addition, we acknowledge that there may be between-group differences that are not captured by our sensitivity analyses. While this study is focused on the neurostructural correlates of VEGF rs699947, an investigation of peripheral VEGF could bolster our findings and strengthen the association between C-allele carriers and VEGF protein expression.
Previous reports have correlated rs699947 C-allele carriers with increased VEGF serum expression (Shahbazi et al., 2002; Steffensen et al., 2010), but this does not necessarily translate to higher VEGF expression in the brain. Investigations of potential effects of VEGF rs699947 on alternative splicing at the VEGFA gene (i.e., the expression of specific VEGF isoforms) may be of interest. We investigated one polymorphism in VEGF, although there are likely other genetic markers that affect VEGF gene expression and/or protein function. Finally, in this cross-sectional study, we cannot make conclusions about whether our neurostructural findings preceded the onset of BD or vice versa.
Conclusion
In summary, this study provides preliminary evidence correlating VEGF rs699947 with neurostructural phenotypes in youth with BD. We have identified diagnosis-related differences in neurostructural correlates of VEGF rs699947 in regions that are relevant to BD. We speculate that context-dependent VEGF signaling in the brain may be implicated, where the neuroprotective and proinflammatory actions of VEGF intersect and possibly act in opposition. Furthermore, given that VEGF is a regulator of other neurotrophins such as BDNF, follow-up investigations may explore whether polymorphisms in these factors interact synergistically to influence brain structure and function.
Future studies should also determine whether these neurostructural phenotypes are associated with peripheral VEGF levels, and whether such associations differ by diagnosis. Overall, this preliminary study reports a relationship between VEGF and brain structure in the early stages of BD, provides further evidence of the vascular-bipolar connection, and highlights the importance of the neurovascular nexus in the understanding of brain structure beginning in early life.
Footnotes
Clinical Significance
The vascular-bipolar connection has substantial relevance from clinical and scientific perspectives. This connection is often attributed to environmental factors, whereas little is known about genetic factors. Vascular endothelial growth factor (VEGF) has a combination of vascular and neurotrophic effects, leading to our focus on VEGF rs699947. Findings from this study link a vascular-related genetic variant, VEGF rs699947, with brain structure in youth with BD who are medically healthy and early in their course of illness, thus our study lends further support of the vascular-bipolar link. Present findings add to a sparse literature surrounding the interplay between genetics, vascular metrics, and neurostructural phenotypes in mood disorders.
Acknowledgments
The authors would like to thank all our participants, their families, and the staff at the Centre for Youth Bipolar Disorder. We thank Diana Khoubaeva for her assistance with the article.
Author's Contributions
N.E.Y.: Data analysis and interpretation, visualization, and writing-original draft. M.K.D.: Data analysis and interpretation and writing-review and editing. K.G.K.: Software, data analysis, and writing-review and editing. C.C.Z.: Technical guidance and writing-review and editing. J.L.K.: Technical guidance and writing-review and editing. B.J.M.: Technical guidance and writing-review and editing. B.I.G.: Supervision, conceptualization, methodology, writing-review and editing, and funding acquisition.
Disclaimer
The Ontario Mental Health Foundation and Canadian Institutes of Health Research had no further role in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.
Disclosures
The authors report no conflict of interest. J.L.K. is a member of the Scientific Advisory Board of Myriad Neuroscience (unpaid); J.L.K. and C.C.Z. hold several patents relating to genetic markers of suicide and pharmacogenetic tests for psychiatric medications.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.