
Abstract
Background:
Few biological or clinical predictors guide medication selection and/or dosing for attention-deficit/hyperactivity disorder (ADHD). Accumulating data suggest that genetic factors may contribute to clinically relevant pharmacodynamic (e.g., dopamine transporter—SLC6A3 also commonly known as DAT1) or pharmacokinetic (e.g., the drug metabolizing enzyme Cytochrome P450 2D6 CYP2D6) effects of methylphenidate (stimulant) and atomoxetine (non-stimulant), which are commonly prescribed medications. This is the first study of youth with ADHD exposed to both medications examining the clinical relevance of genetic variation on treatment response.
Methods:
Genetic variations in DAT1 and CYP2D6 were examined to determine how they modified time relationships with changes in ADHD symptoms over a 4-week period in 199 youth participating in a double-blind crossover study following a stepped titration dose optimization protocol.
Results:
Our results identified trends in the modification effect from CYP2D6 phenotype and the time–response relationship between ADHD total symptoms for both medications (atomoxetine [ATX]: p = 0.058, Methylphenidate [MPH]: p = 0.044). There was also a trend for the DAT1 3′ untranslated region (UTR) variable number of tandem repeat (VNTR) genotype to modify dose relationships with ADHD-RS total scores for atomoxetine (p = 0.029). Participants with DAT1 9/10 repeat genotypes had a more rapid dose–response to ATX compared to 10/10, while those with 9/9 genotypes did not respond as doses were increased. Regardless of genotype, ADHD symptoms and doses were similar across CYP2D6 metabolizer groups after 4 weeks of treatment.
Conclusions:
Most children with ADHD who were CYP2D6 normal metabolizers or had DAT1 10/10 or 9/10 genotypes responded well to both medications. While we observed some statistically significant effects of CYP2D6 and DAT1 with treatment response over time, our data indicate that genotyping for clinical purposes may have limited utility to guide treatment decisions for ATX or MPH because both medications were generally effective in the studied cohort after 3 weeks of titration to higher doses. The potential DAT1 association with ATX treatment is a novel finding, consistent with prior reports suggesting an association of the DAT1 in 9/9 genotypes with lower responsive rates to treatment at low and moderate doses.
Introduction
Attention-deficit/hyperactivity disorder (ADHD) is a common neurodevelopmental disorder that is frequently treated with stimulant and non-stimulant medications. ADHD is highly familial, with twin and family studies demonstrating high heritability for ADHD symptoms (Faraone and Larsson, 2019). Despite decades of use and a widening proliferation of ADHD treatment formulations, there are few reliable biomarkers or clinical predictors to guide the initial selection of medications or inform treatment changes for cases presenting with poor, partial, or non-response. Stimulant medications are often chosen as first-line medications due to their efficacy in reducing ADHD symptoms, while non-stimulants are used less frequently. Although the vast majority of children display a positive initial response to stimulants, 20%–25% display poor tolerability or non-response and are subsequently treated with another ADHD medication, either a different stimulant or non-stimulant (Charach et al., 2004; Owens et al., 2003; Thiruchelvam et al., 2001). Given that the majority of ADHD youth respond to stimulants, either the ability to predict stimulant non-response or to predict response to non-stimulants would inform a personalized medicine approach to treatment selection.
There has been considerable interest in utilizing pharmacogenetic information to personalize ADHD treatment (Froehlich et al., 2010). Pharmacogenetic studies to date have largely focused on pharmacodynamic genes related to catecholamine metabolism and signaling, or pharmacokinetic genes related to drug metabolism (Volkow et al., 2002a; Volkow et al., 2002b). Candidate gene studies have been informative in identifying potential sources of variability in dosing and response in separate studies of stimulants and non-stimulants. There are no studies, however, examining the impact of pharmacogenetic variation on treatment outcomes in participants sequentially exposed to both stimulants and nonstimulants which are needed to assess their clinical relevance.
Methylphenidate (MPH) is the most prescribed and studied stimulant medication in children (Faraone et al., 2019). MPH binds to norepinephrine (NE) and dopamine (DA) transporters and blocks reuptake into presynaptic neurons, increasing these monoamine’s availability in the extraneuronal space (Faraone, 2018). The DA transporter is encoded by the SLC6A3 solute carrier family 6 member 3 gene located at chromosome 5p15.3. SLC6A3 is also known as DAT1, which will subsequently be used herein for conceptual alignment with the DAT1 protein product. DAT1 function and expression have been examined in relation to a number of genetic variants in this gene, with notable influence by a variable number of tandem repeats (VNTRs, rs28363170) in the gene’s 3′ untranslated region (3′UTR) (Salatino-Oliveira et al., 2018). The number of repeats ranges from 3 to 11, with the 10/10 and 10/9 repeat genotypes being more common than the 9/9 repeat. Carriers of the 9 repeat allele have been reported to have decreased DAT1 expression compared to 10/10 homozygotes (Fuke et al., 2001; Mill et al., 2002). Knockout studies of DAT1 in rodents identified that this gene is important for controlling synaptic DA concentrations and that changes in DAT1 protein expression result in broader changes across DA signaling pathways (Leo and Gainetdinov, 2013). In a meta-analysis, the 9-repeat (9R) allele, which regulates DA activity in striatal brain regions, was found to be associated with increased DAT1 activity (Faraone et al., 2014).
DAT1 pharmacogenetic studies to date have revealed important findings, with some heterogeneity across studies employing different methods and populations, that underscores the need to define and better understand variable drug response (Bruxel et al., 2014). For example, in the first ADHD pharmacogenetic study focusing on a pharmacodynamic gene, participants homozygous for the DAT1 10 repeat allele (10R) were found to have a diminished response to MPH relative to those with a DAT1 10/9 genotype in a cohort of 30 stimulant-naive African-American children (Winsberg and Comings, 1999). However, subsequent studies and a meta-analysis concluded that DAT1 genotype is not consistently associated with stimulant response (Froehlich et al., 2011; Kambeitz et al., 2014; McGough et al., 2006). Since DAT1 genotype frequencies differ markedly across populations with different ethnic ancestries (Shumay et al., 2011), spurious or null findings can result from not controlling for ancestry/ethnicity. Secondly, the majority of studies compared the response of participants with DAT1 10/10 and 9/10 genotypes but did not examine the less common DAT1 9/9 genotype which is a major limitation. Employing fixed stimulant doses and placebo controls, Stein and colleagues reported that children with the less common DAT1 9/9 genotype displayed a dose–response curve distinct from those with the more common 10/10 and 10/9 genotypes (Stein et al., 2005). Similar findings were also reported by two other investigators (Joober et al., 2007; McGough et al., 2006) while a third dose–response study reported that ADHD youth with the DAT1 9/9 genotype were less likely to respond to low doses of stimulants compared to youth with at least one copy of the DAT1 10R allele (Stein et al., 2011). In contrast, Froehlich et al. found that youth with the DAT1 9/9 genotype had a more robust response to MPH than the other genotype groups; this effect was, however, most pronounced at higher MPH doses (Froehlich et al., 2011).
The first and most studied non-stimulant medication approved for ADHD monotherapy is atomoxetine (ATX), which is a selective NE reuptake inhibitor that is primarily metabolized by CYP2D6 (Brown and Bishop, 2015). The FDA product label suggests limiting dose increases in patients who are known to be CYP2D6-poor metabolizers (i.e., having two nonfunctional alleles) (Anonymous, n.d.). CYP2D6 is a highly polymorphic gene with common genetic variation across patients that results in a broad range of metabolic activity among individuals. To date, the Pharmacogene Variation Consortium (PharmVar) (Gaedigk et al., 2021) has defined over 160 unique haplotypes (so-called star alleles) not counting the array of structural variants including gene duplications, and hybrid genes that are part CYP2D6 and part CYP2D7 (Gaedigk et al., 2021; Nofziger et al., 2020). Allele frequencies vary considerably with some being found predominately in certain populations or ethic groups. Each person has two CYP2D6 haplotypes (or star alleles) that define their diplotype (often referred to as a genotype). An activity scoring (AS) system is commonly used to translate genotype to phenotype (Gaedigk et al., 2008). Briefly, a value is assigned to each allele and the sum of both alleles yields the diplotype activity score. Poor metabolizers (PMs) have AS = 0, intermediate metabolizers (IMs) have AS = 0.25–1, normal metabolizers (NM) have AS 1.25–2.25, and ultrarapid metabolizers have AS >2.25 (Caudle et al., 2020).
In the largest study to date investigating CYP2D6 variation and ATX, Michelson and colleagues reported that PMs displayed greater reductions in mean ADHD symptom severity during ATX treatment compared with NMs, increases in cardiovascular tone, and some differences in tolerability compared with CYP2D6 non-poor metabolizers taking similar doses (Michelson et al., 2007). CYP2D6 modeling studies in children with ADHD identified that dose-corrected ATX systemic exposure varied nearly 30-fold (Brown et al., 2016) and oral clearance varied nearly sevenfold (Cheng et al., 2023) across genotype-predicted metabolizer groups. Steady-state simulations of these data as well as therapeutic drug monitoring studies (Guo et al., 2024) suggest that many CYP2D6 non-PMs may not achieve adequate exposure at commonly used doses. A broader review of the literature highlights how plasma half-life increases nearly fourfold (from 4.5 to 20 hours) in CYP2D6 PMs compared to NMs, and that exposure [area under the concentration curve (AUC)] was ∼10 times greater in PMs as compared to NMs (Brown and Bishop, 2015). Recently, the Clinical Pharmacogenetics Implementation Consortium (CPIC) and Dutch Pharmacogenetic Working Group (DPWG) both evaluated an extensive body of in vitro, in vivo, pre-clinical, and clinical trial data and published guidelines for ATX dosing based on CYP2D6 genotype (Brown et al., 2019; Nijenhuis et al., 2022).
As previously noted, several, but not all studies reported associations between variation in the DAT1 and CYP2D6 genes with dose–response characteristics for MPH and ATX, respectively, (Brown and Bishop, 2015; Myer et al., 2018). However, the clinical relevance of these associations remains unclear. The present study examined the moderating effect of DAT1 and CYP2D6 genetic variation on medication response within a large, two-site, comparative effectiveness study that employed a randomized, double-blind crossover design. Each participant of this ethnically diverse study cohort was titrated to optimal response of ATX and MPH. The goal of the study was to investigate time-response profiles of CYP2D6 and DAT1 on ATX and MPH, respectively, in the context of a flexible titration dose design, to gain valuable insights on how to best select and titrate these medications based on genotype information. Based on prior studies, we hypothesized that individuals with a DAT1 9/9 genotype would be associated with a decreased response to MPH and that CYP2D6 NM and/or UMs would exhibit decreased response to ATX as compared to IMs or PMs.
Methods
Participants and assessments
Participants were recruited from clinical sites in Chicago and New York City for an NIH-funded randomized, double-blind, double-dummy, counterbalanced, crossover study comparing ATX, and MPH for the treatment of ADHD. The clinical trial serving as the source of materials for this study was registered at ClinicalTrials.gov (NCT00183391). A detailed description of the clinical trial, procedures, and initial findings have previously been published by Bedard et al. (Bédard et al., 2015) Parents gave written consent and participants gave verbal assent after a detailed description of the study was provided and an adequate understanding of study procedures was verified. Individuals included in the present analysis represent those who agreed to a blood draw for subsequent DNA isolation and genetic studies. The study was approved by the Institutional Review Boards at the University of Illinois at Chicago and the Icahn School of Medicine at Mount Sinai.
Eligible patients had a DSM-IV diagnosis of ADHD, any subtype, according to a Kiddie Schedule for Affective Disorders and Schizophrenia Present and Lifetime Version (K-SADS-PL) interview (Kaufman et al., 1997). Participants were excluded if no parent or legal guardian could provide consent in English. Additional exclusion criteria included significant medical illness or neurological disease that precluded treatment with either medication, any established history of psychosis or bipolar disorder, or established previous non-response or intolerability to ATX or MPH. Participants were 6–17 years of age, had a current IQ (WISC-III, IV, or V or equivalent measure) full-scale score >75, and an ADHD Rating Scale-IV-Parent Version (ADHD-RS-IV) severity score of 1.5 SD above sex and age means for either the hyperactivity/impulsivity or inattention scales, or total score (Reid et al., 1998). Sixty-five percent of participants were naïve to pharmacologic treatments for ADHD. A total of 232 children ages 7–17 (mean age = 10.5 ± 2.7 years) were randomized and 199 had completed at least one follow-up treatment visit with both medications and had genomic DNA available for pharmacogenetic studies. Efficacy assessments included weekly symptom improvement and response measures including the ADHD-RS-IV and the Clinical Global Impressions Severity (CGI-S) and Clinical Global Impressions-Improvement (CGI-I) scales completed by blinded raters.
Medication titration
Each study participant sequentially received MPH and ATX, with the order of treatment randomized. A schematic of the study design is summarized in Figure 1. Two capsules of OROS MPH or matching placebo and either two or three capsules of ATX (determined by the child’s weight) with placebo were used to standardize the number of assigned capsules administered each morning. Titration followed an escalating, dose optimization approach, using weekly stepped dose increases consisting of up to four dose levels for each medication (MPH: 18, 36, 54, and 72 mg; ATX: 0.5, 1.0, 1.4, and 1.8 mg/kg). There was a 2-week unblinded placebo washout between the treatments prior to beginning the second treatment block. The active medication blocks were intended to last 3–5 weeks, with each participant having two weeks of exposure to the best-tolerated dose of medication. However, shortening or lengthening the exposure in either block was permissible, to address ethical constraints in the case of poor response or tolerability, and to maximize sample retention. To examine time–response relationships, response across four weeks of treatment with either ATX or MPH was evaluated. This was done to standardize time on each dose for purposes of analyses described herein, and because the majority of participants had at least four weeks of treatment for each medication.

MACRO pharmacogenetic study design.
Genetic analyses
Genomic DNA was extracted from 10 mL of whole blood using a PureGene® kit and concentrations were adjusted to 50 ng/μL for subsequent analyses. Two candidate genes known to have common and clinically relevant genetic polymorphisms were selected for association studies based on prior functional relevance to the mechanism of action or metabolism of ATX or MPH. Variants selected for the study included the VNTR in 3′UTR of DAT1, and 12 CYP2D6 variants; CYP2D6 genotyping also included testing for gene deletions and duplications. Genotyping for DAT1 was completed as previously described using an ABI 3730XL capillary instrument to identify the number of repeats (O’Brien et al., 2013). CYP2D6 genotyping was accomplished using previously described approaches (Gaedigk et al., 2012; Gaedigk et al., 2010a; Gaedigk et al., 2010b; Gaedigk et al., 2008). Briefly, commercially available TaqMan assays (Thermo Fisher Scientific, Waltham, MA) were utilized to genotype a 7 kb-long CYP2D6-specific XL-PCR fragment as a template. The following variants were interrogated: rs16947, rs1080985, rs35742686, rs3892097, rs5030655, rs5030867, s72549350, rs1065852, rs28371706, rs59421388, and rs28371725. Testing these variants allowed the identification of CYP2D6*2, *3, *4, *6, *7, *9, *10, *17, *29, and *41. These represent tier 1 alleles recommended by the Association for Molecular Pathology for clinical assessments of CYP2D6 (Pratt et al., 2021). The CYP2D6*5 gene deletion, and presence of gene duplications or multiplications, and CYP2D6:CYP2D7 hybrid genes were determined by long-range PCR (XL-PCR). In addition, quantitative gene copy number variation (CNV) analyses were performed with TaqMan copy number assays (Thermo Fisher Scientific) targeting intron 6 and exon 9 (CNV C__32407229_60 and Hs00010001_cn) as recommended by the manufacturer. All TaqMan assays were run on an ABI StepOnePlus instrument using StepOnePlus and TaqMan Genotyper software. Approximately 10% of samples were selected for duplicate genotyping or resequencing to verify calls, all were 100% concordant. Genotype frequencies did not significantly deviate from the Hardy–Weinberg equilibrium.
Although self-reported ethnicity is commonly used in clinical research, objective measures such as ancestry informative markers (AIMs) afford more precise categorization. In addition to candidate gene assessments, 105 AIMs were genotyped using the Sequenom MassARRAY platform (Giri et al., 2009). Genetic ancestry was quantified for each participant for European, West African, and Native American components. Individual ancestry estimates were obtained from the genotype results using the Bayesian Markov Chain Monte Carlo method implemented in the program STRUCTURE 2.1 and were scored from 0% to 100% for each ancestry group (Falush et al., 2003).
CYP2D6 star alleles were called based on PharmVar definitions and collated with CNV results. Subsequently, each diplotype was translated into phenotype (i.e., assigned as PM, IM, NM, or UM) using the activity score (AS) method (Gaedigk et al., 2008). AS were then used to assign CYP2D6 metabolizer status, or phenotype, using an expert consensus genotype-phenotype translation approach such that AS 0 = poor metabolizers (PMs; n = 12), 0.25–1 = intermediate metabolizers IMs; n = 69), 1.25–2.25 = normal metabolizers (NMs; n = 101), and >2.25 = ultrarapid metabolizers (UMs; n = 12) (see Supplementary Table S1) (Caudle et al., 2020).
Statistical analysis
Responses to MPH and ATX were examined separately. Change in ADHD-RS total score over time was the primary outcome measure. The ADHD-RS Inattention and Hyperactivity/Impulsivity subscales along with the Clinical Global Impression (CGI-I) improvement scores were also used to assess response as secondary outcome measures. All participants with at least one assessment were included in the analysis. Over the course of the 4-week evaluation, 91% of participants finished at least three assessments, and 79% finished all four assessments. We performed data analysis through mixed effects regression models. Missing values were not imputed.
Impacts of CYP2D6 genotype-predicted phenotype groups and DAT1 genotype on the time–response trajectories were examined for MPH and ATX using multivariable linear mixed effects regression models (Twisk, 2013). Due to the design of the study, dose and time were tightly coupled, leading to almost identical modeling estimates. For ease of interpretation, genotype (DAT1) or genotype-predicted phenotype groups (CYP2D6) were utilized to assess symptom changes over time. Specifically, time was modeled as a predictor, and a time-by-genotype (DAT1) and time-by-phenotype (CYP2D6) interaction term was included in all models to assess the genotype effect on response over time. Age, sex at birth, percent European and West African ancestries, prior stimulant exposure, and baseline ADHD symptoms were included as covariates in all models. The within-subject correlation was accounted for by subject-specific random intercept. Fixed effects were estimated using the restricted maximum likelihood method and tested with Kenward-Roger’s approximation of degree of freedom (Kenward and Roger, 1997). All analyses were conducted using Stata 14.2 with a two-tailed p < 0.05 threshold for significance.
Results
The clinical and demographic characteristics of the study cohort are summarized in Table 1. The cohort was diverse with regard to self-reported ethnicity, with 37% self-identified as White, non-Hispanic, 32% Black/African American, 22% Hispanic/Latinx, and 10% other (i.e., Native American or Alaska Native, Asian, Native Hawaiian or Pacific Islander, or mixed). Genotyping success rates were 196/199 (98%) for DAT1 and 194/199 (97%) for CYP2D6. Observed genotype frequencies for DAT1 were 9/9 (n = 14), 9/10 (n = 58), 10/10 (n = 111), and other (a combination of rare shorter or longer repeats n = 13 that were excluded from further analyses). Consistent with prior population studies, differences in CYP2D6 phenotype group frequencies were observed across self-reported ethnic groups, as well as the proportion of European and African genetic ancestry (Gaedigk et al., 2017). There were no other significant demographic differences or differences in ADHD symptom severity for CYP2D6 phenotype and DAT1 genotype groups (Table 1).
Baseline Patient Characteristics, Overall, and by Study Arms
ADHD, attention-deficit/hyperactivity disorder; CGI, Clinical Global Impressions.
Initial efficacy results of treatment are presented elsewhere (Bédard et al., 2015), and are summarized here with a focus on the pharmacogenetic analyses (Table 2). Significant main effects of time indicative of therapeutic improvement were observed for both medications, beginning at week three for ATX and week two for MPH. Younger age (p < 0.05 for both drugs) and those with higher ADHD-RS scores at baseline (p < 0.001 for both drugs) displayed greater reduction in ADHD symptoms over time. Neither sex (p > 0.5 for both drugs) nor genetically defined ancestry was significantly associated with response for either drug (p > 0.2).
Modification Effects of CYP2D6-Defined Metabolizer Groups on ADHD Symptom Trajectories Over Time
All were based on linear mixed effects regression models which adjusted for baseline outcome values, prior stimulant naïve status, age, sex, European and West African ancestry as fixed covariates, and a subject-specific random effect for clustering. Bold data highlight statistically significant p-values <0.05.
ADHD, attention-deficit/hyperactivity disorder; CGI, Clinical Global Impressions; IM, Intermediate Metabolizer; NM, Normal Metabolizer; PM, Poor Metabolizer; UM, Ultrarapid Metabolizer.
ATX and MPH relationships with CYP2D6
A trend was observed suggestive of an overall moderation effect from the CYP2D6 metabolizer group on the time–response relationship between ADHD total symptoms and both medications (ATX: p = 0.058, MPH: p = 0.044). During ATX treatment, this overall trend was driven by significant effect modification for CYP2D6 IMs on ADHD-RS total and scores. IMs displayed significantly less symptom improvement at week three as compared to NMs (Table 2) but had similar overall outcomes after week 4 (Fig. 2). During MPH treatment, the overall trend was driven by significant effect modification for UMs; these individuals showed less symptom improvement at week 3 as compared to NMs (Table 2), but had similar outcomes overall after week 4. Dose comparisons across genotype groups for each medication are presented in Supplementary Tables S2, S3, S4, and S5.
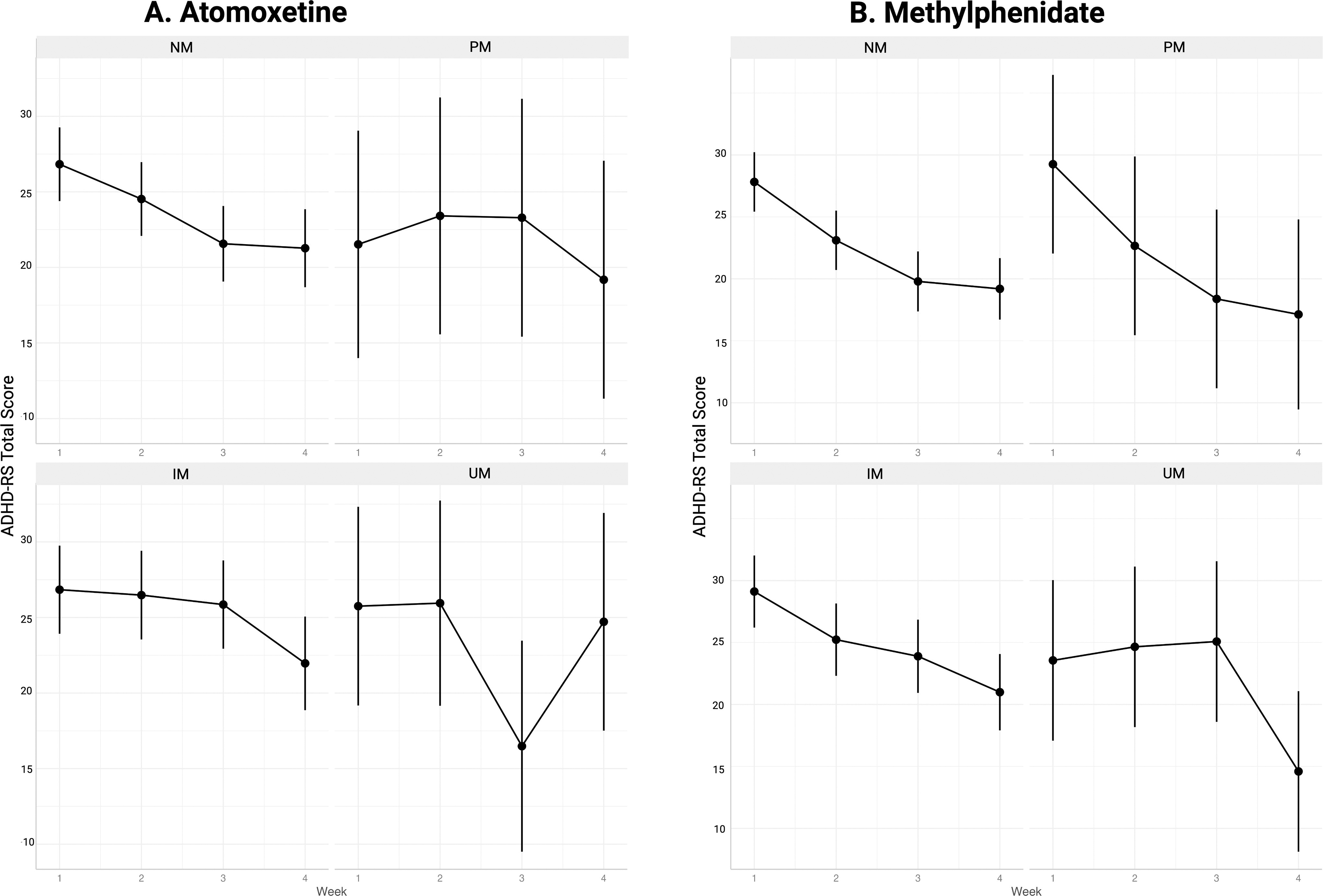
Symptom change over time by CYP2D6 phenotype group during treatment with atomoxetine
ATX and MPH relationships with DAT1
DAT1 genotype significantly modified the overall time response relationship between ATX and ADHD total symptoms (p = 0.029). Changes in inattention and hyperactivity subscale scores of the ADHD-RS were similar to total scores with respect to the direction of effect and overall patterns of response over time by genotype groups. As shown in Figure 3, better response was observed in participants with the DAT1 9/10 genotype as compared to those with the 10/10 genotype (Table 3). In the participants with prior stimulant exposure, increased ATX doses had little impact on ADHD symptoms for participants genotyped as DAT1 9/9, while trends of more pronounced dose relationships with symptom improvement were observed in those with the more common DAT1 9/10 or 10/10 genotypes, and for those who were stimulant-naïve (p < 0.1).
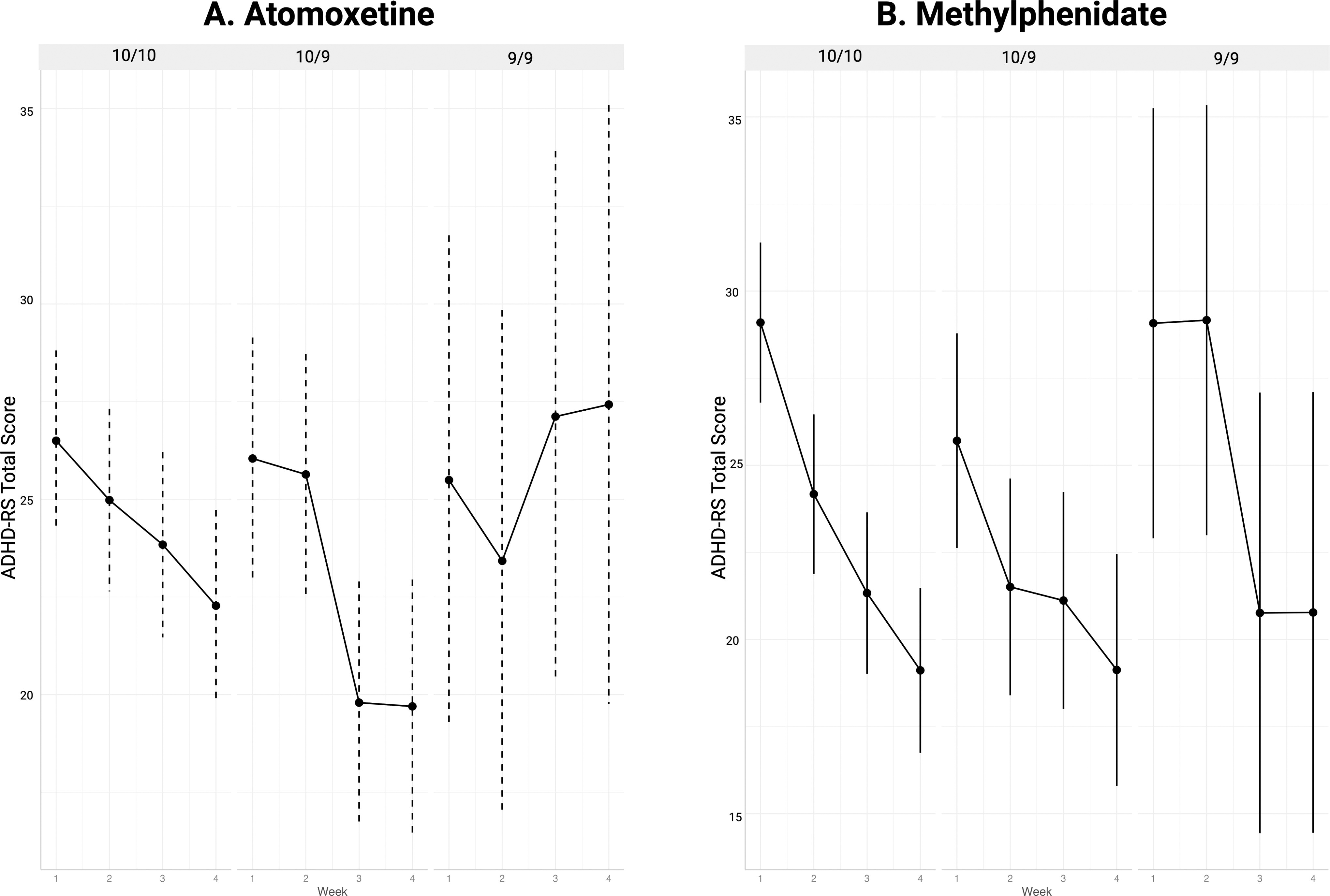
Symptom change over time by DAT1 genotype group during treatment with atomoxetine
Modification Effects of DAT1 Genotype (10/10, 9/10, and 9/9) on ADHD Symptom Trajectories Over Time
All results were based on linear mixed effects regression models, which adjusted for baseline outcome values, prior stimulant naïve status, age, sex, European and West African ancestry as fixed covariates, and subject-specific random effect for clustering.
Bold data highlight statistically significant p-values <0.05.
ADHD, attention-deficit/hyperactivity disorder; CGI, Clinical Global Impressions.
DAT1 genotype did not modify the overall time response relationship for MPH (p = 0.14). Notably, the DAT1 9/9 group did not display typically observed patterns of improved ADHD symptom reductions during treatment unless exposed to higher doses (54 and 72 mg, respectively) at week 3 of the study (Fig. 3).
Discussion
In this large, prospective, crossover study employing flexible dose titration and a robust range of doses, our data show initial variation in response moderated by DAT1 or CYP2D6 at lower doses that resolved and did not robustly impact the extent of symptom response after 4 weeks of treatment. Both MPH and ATX were associated with significant reductions in ADHD symptoms over time and with increasing doses. This report focused primarily on acute ADHD symptom response, although common adverse events and tolerability are often the rate-limiting factors in ADHD titration and adherence rather than short-term efficacy. The majority of children with ADHD, and those with the common DAT1 10/10 and 10/9 genotypes and CYP2D6 NMs responded well to both medications. This is not surprising and consistent with prior observations that the majority of youth with ADHD respond positively to initial pharmacological treatment. No evidence was found supporting that variation in DAT1 or CYP2D6 robustly impacts the extent of symptom response after 4 weeks of treatment.
For CYP2D6, the results were surprising and contrasting with previous work (i.e., PMs did not respond differently than NMs over time when treated with ATX). While some trends for altered patterns of symptom improvement trajectories were observed in IM and UM individuals during the third week of treatment, no differences in ADHD response were recorded by week 4. Previously, concerns have been raised regarding the potential for CYP2D6 NMs or UMs to unintentionally receive inadequate ATX exposure when treated with standard weight-based approaches per the product labeling (Brown et al., 2019; Brown et al., 2016). Findings from our study, however, did not reveal clinically relevant associations with response trajectories over time and dose for either ATX or MPH.
Although not statistically significant, some patterns of attenuated response were observed in subgroups including the DAT1 9/9 genotype and MPH dosing. A significant main effect of DAT1 genotype or interaction with time was not detected during MPH treatment in the present study, in contrast to prior dose–response studies finding a significant effect of genotype (Froehlich et al., 2011; Stein et al., 2014; Stein et al., 2005). However, as shown in Figure 3, those with the DAT1 9/9 genotype tended to have less improvements in ADHD symptoms until after week 3 (target dose 54 mg) compared to those with DAT1 9/10 or 10/10 genotypes. Also, to our knowledge, this is the first study interrogating genetic variation in DAT1 and ATX treatment, and the trend seen for DAT1 9/9 to moderate the effects of ATX on ADHD was unexpected. This finding may suggest that patients with the less common DAT1 9/9 genotype may be at risk of having a worse prognosis and outcome when exposed to lower doses in general, as opposed to DAT1 being of exclusive relevance to stimulants. Supporting DAT relevance to ATX, one small SPECT imaging study also suggested that striatal DAT availability decreased over the course of 8 weeks of ATX treatment (Akay et al., 2015). It is notable that the DAT1 9/9 genotype has previously been reported to be associated with a differential response to behavior interventions which were not examined in the present study. For example, the Incredible Years behavioral parent program proved to be most effective in decreasing parent-reported (but not observed) externalizing behavior in boys (but not girls) carrying more, rather than fewer, dopaminergic plasticity alleles (DAT1 9/9 was considered a lower plasticity genotype in that study) (Chhangur et al., 2017). Moreover, in a long-term follow-up study of ADHD youth, the DAT1 9/9 genotype was associated with 5-year reductions in estimated life expectancy (Barkley et al., 2019). Perhaps variation in this gene may be related to a general lack of response phenotype as compared specifically to stimulant medications as previously hypothesized.
Our findings need to be interpreted in the context of the limitations of the study design, length of treatment, and number of participants. Although the cohort was large relative to other ADHD pharmacogenetic studies, there were still relatively small numbers of DAT1 9/9, CYP2D6 UMs, and PMs, whose responses were hypothesized to be most influenced by genotype. We also note that some rare allelic variants in CYP2D6 and DAT1 were not included in our testing or analyses. Furthermore, the MACRO study design did not allow for the disentangling of time and dose, as these were highly correlated. By examining the first 4 weeks of treatment, however, we were able to examine effects where the majority had at least three dose-titration visits. Although a longer follow-up period may have captured more patients with beneficial MPH or ATX treatment, most positive responses were evident by the 4-week time point. Additionally, there are other candidate genes, for example, carboxylesterase 1 (CES1) (Brown et al., 2022), and the NE transporter (SLC6A2) (Ramoz et al., 2009) which may contribute to variable response and were not studied in this investigation.
Since the majority of youth with ADHD initially respond well to stimulants, studies of pharmacogenetic and clinical predictors of non-response may be more impactful than studies of response per se. Future studies aimed at identifying poor or non-responders and employing enriched samples with genotypes of interest, polygenic risk scoring, and/or whole genome sequencing may ultimately be more relevant for predicting treatment response for individual patients than limited candidate gene approaches. Additionally, measuring broader response phenotypes, such as adverse events or factors contributing to tolerability and/or premature discontinuation may be more likely to improve treatment personalization. With advances in informatics and more uniform use of electronic medical records, there is the potential of obtaining data for sufficiently large sample numbers to examine predictors of poor tolerability over time and also rare, but potentially serious adverse events.
Conclusions
The majority of children with ADHD, and those with the more common DAT1 10/10 and 10/9 genotypes and CYP2D6 NMs responded well to both medications. While we observed some statistically significant effects of CYP2D6 and DAT1 with treatment response over time, our data indicate that genotyping for clinical purposes may have limited utility to guide treatment decisions for ATX or MPH because both medications were generally effective in the studied cohort after three weeks of titration to higher doses. A potential association between DAT1 genetic variation with ATX treatment is a novel finding. These findings along with observations that patients with with the DAT1 9/9 genotype may not respond as well to low or moderate dose levels of medications and requires further study as a high-risk ADHD subgroup.
Footnotes
Acknowledgments
The authors thank Shitalben Patel and Katarzyna Drozda for their assistance with the laboratory analyses and to Heather Phillips, Jennifer Davidow, Jodi Uderman Lipsky, Joanne Philips, Erika Zeranski, Juan Pedraza, and Iliyan Ivanov for their assistance with data collection.
Disclosures
J.R.B. has served as a consultant to OptumRx. M.A.S. has served as a consultant to Genomind, Supernis, Periapt, and Medici. J.H.N. is/has been an advisor and/or consultant for Hippo T&C, Ironshore, Lumos, Medice, MindTension, OnDosis, Otsuka, Supernus. He has received research support from the National Institute on Drug Abuse (NIDA), the Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD) and Otsuka. He also has received honoraria from for disease state presentations from Otsuka, and served as a consultant for the US National Football League. The other authors report no disclosures or potential conflicts of interest relevant to this study.
Funding Information
This project was funded in part by grants from the NIMH (R01
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.