
Abstract
Introduction
Cytokine-induced killer cells (CIKs) are heterogeneous cells derived from human peripheral mononuclear cells (PBMCs), which are cultured in the presence of multiple cytokines, including recombinant human interleukin 1α, (rhIL-1α), interleukin 2 (rhIL-2), interleukin-γ (rhIFN-γ), and mouse anti-hCD3 monoclonal antibody (McAb). 1 –3 CIKs are known as potent antitumor effectors with several advantages over the lymphokine activated killer cells (LAKs), and tumor infiltrating lymphocytes (TILs), which are expandable both from healthy donors and tumor patients with similar cell expansion efficiency, and well tolerated both in animal models and human patients. As known, CD3+CD56+ cells that coexpress CD8 rather than CD4 are the main cytotoxic population in CIKs. But in our previous study, we found CD4+ T cells occupied a considerably high proportion in harvested CIKs, 4 and a typical Fas-mediated apoptosis was identified in sensitive lymphoma cell line Raji cells after coculture with CD4+ T cells in CIKs (CD4+ CIKs), accompanied with dramatic increased expression of Fas on membrane and sustainable high levels of interferon-γ (IFN-γ) in supernatant. 5 All these data prompted us to propose a hypothesis that the antitumor cytotoxicity of CD3+CD56+ cells is boosted up by CD4+ CIKs, which induce apoptosis in tumor cells through the Fas ligand (Fas/FasL) pathway. Although the concrete mechanism is still unclear, 2 candidate molecules might be involved in this Fas-dependent apoptosis progress. One is CD40L, which combines with CD40 protein and upregulates Fas expression on many types of tumor cells. 6,7 The other is IFN-γ, which directly increases Fas expression. 8,9 Both are highly expressed in activated CD4+ T cells. 10 However, the possibility that the proapoptotic effects of these 2 molecules might be shadowed by that of FasL highly expressed on CIKs should not be excluded in Fas-sensitive Raji cells. So we selected a Fas-resistant breast cancer cell line MDA-MB-231, which shows innate but reversible resistance to Fas-mediated apoptosis under certain conditions. We cocultured MDA-MB-231 cells with enriched CD4+ CIKs, evaluated the correlation between Fas expression and apoptotic rates, and compared the proapoptotic effects of CD40/CD40L ligation and IFN-γ stimulation in order to identify which is the predominant factor of Fas-dependent apoptosis induced by CD4+ CIKs.
Patients, Materials, and Methods
Patients and Cell Lines
Fifteen (15) patients with malignant solid tumors (12 men, 3 women, mean ± standard deviation age 62.6 ± 7.3 years, range 53–72 years) were enrolled with written consent for this experiment. Two (2) human breast carcinoma cell lines, T47D and MDA-MB-231, were purchased from American Type Culture Collection (ATCC) and preserved in our laboratory. The expression of CD40 and Fas on 2 cell lines was examined by flow cytometry (BDAria, BD, Biosciences, San Diego, CA), which was 90.4% and 21.4% in T47D and 95.7% and 3.9% in MDA-MB-231, respectively.
Auto-CIK Amplification and CD4+ CIK Purification
Leukopheresis was administered to patients using CS3000plus blood cell separator (Baxter, Deerfield, IL). PBMCs were enriched and CIKs were generated as described elsewhere. 4 Fresh CD4+ CIKs and CD4+ PBMCs were purified by a magnetic beads separation system (CD4+ T cell isolation kit II, Miltenyi Biotec, Borgisa, Gladbach, Germany) following the operation manual. Purity and viability were determined by flow cytometry and trypan blue staining, respectively.
Expression of CD40L and Secretion of Cytokines in CD4+ CIKs
The expression of 2 active types of CD40L (membrane-attached CD40L and soluble CD40L) in CD4+ CIKs and CD4+ PBMCs were evaluated at mRNA and protein levels by semiquantitative reverse-transcriptase polymerase chain reaction (RT-PCR), flow cytometry, and enzyme-linked immunosorbent assay (ELISA) method. The secretion of interleukin-2 (IL-2), IFN-γ, and tumor necrosis factor-α (TNF-α) in supernatants from CD4+ CIKs and CD4+ PBMCs after 4- and 24-hour culture in serum-free medium X-VIVO 20 (Cambrex, East Rutherford, NJ) at 2 × 106 cells/mL were simultaneously measured by ELISA.
Identification of Sensitivity to Fas Activation
MDA-MB-231 cells were cocultured with or without supernatants of CD4+ CIKs before adding agonist anti-Fas monoclonal antibody (CH11, Upstate Biotech, Billerica, MA). Following incubation, the apoptosis of MDA-MB-231 cells was evaluated using flow cytometry.
Cell Coculture and Antibody Block
Two (2) breast cancer cell lines, Fas-sensitive T47D cells and Fas-resistant MDA-MB-231 cells, were cocultured with CD4+ CIKs at ratios of 1:1, 1:5, and 1:10 for 24 hours. Fas expression and early stages of apoptosis were detected using flow cytometry. To investigate the key regulator of the apoptosis induced by CD4+ CIKs, 3 neutralizing McAbs, including anti-human FasL McAb (ZB4, eBioscience, San Diego, CA), anti-human CD40L McAb (B-B29, Diaclone Research, USA), and antihuman IFN-γ McAb (B-B1, Diaclone Research, Besancon, France) were added into the coculture system composed of MDA-MB-231 cells and CD4+ CIKs at ratio of 1:5 for either 6 or 24 hours. The apoptotic rates and expression of Fas were detected 24 hours after incubation and compared with the controls. In order to determine changes in apoptotic signal pathways, the expression of 4 representative apoptosis-related genes (including Bcl-2, Bax, Fas-associating protein with death domain [FADD], and FLICE inhibitory protein [c-FLIP]) were measured 6 and 24 hours after incubation at RNA level using real-time quantitative RT-PCR assay.
Semiquantitative and Real-Time Quantitative RT-PCR
The mRNA expression of CD40L gene was measured using semiquantitative RT-PCR assay, in which a roughly relative quantitation was obtained by densitometric analysis and normalized with the density of β-actin. The amplification conditions were as follows: 94°C 30 seconds, 56°C 30 seconds, 72°C 45 seconds, 25 cycles. Two (2) proapoptotic genes, FADD and Bax, and 2 antiapoptotic genes, c-FLIP and Bcl-2, were quantified using real-time quantitative RT-PCR assay with LightCycler RNA Amplification kit SYBR Green I (Roche Molecular Biochemicals, Roche, Basel, Switzerland) following the manufacturer's instructions. The relative amount of target was normalized to β-actin and calculated with the formula 2−ΔCt (ΔCt= Ctsample—Ct β−actin). All tests were repeated 4 times to obtain the mean and the standard deviation. The primers of CD40L, Bcl-2, Bax, FADD, c-FLIP, and β-actin are listed in Table 1.
The Primer Sequences for Reverse-Transcriptase Polymerase Chain Reaction
Flow Cytometry
The expression of CD40L and Fas were detected using fluorescein-5-isothiocyanate (FITC)-labeled anti-CD40L, phycoerythrin (PE)-labeled anti-Fas, and PE-Cy5-labeled anti-CD4 monoclonal antibodies (BD PharMingen, San Diego, CA), respectively. The 5 × 105 cells were incubated with 20 μL antibodies each in the dark for 30 minutes. Then the cells were washed and resuspended in the assay buffer (phosphate-buffered saline, 2% newborn calf serum, and 1% sodium azide) for analysis. The isotype-matched IgG1 was used as negative control. The early stage of apoptosis were detected using Annexin-V staining as described elsewhere. 5 The 1 × 105 cells were stained with 5 μL Annexin V-FITC (BD Pharmingen) and 2.5μL propidium iodide (PI) in the dark for 15 minutes. After incubation, 400 μL of 1 × binding buffer were added and apoptosis was analyzed. Untreated cells were used as negative control.
Elisa
The levels of different cytokines in supernatants after 4- and 24-hour culture, including IL-2, IFN-γ, TNF-α and soluble CD40 ligand (sCD40L), were measured by the respective ELISA kits (Bender, Medsystems Vienna, Austria) according to the manufacturer's instructions. Briefly, supernatant samples were added to plates and incubated for 2 hours. After washes, horseradish peroxidase-linked secondary antibodies specific to the cytokines listed above were added. The substrate solution was added after washing, and color was measured at 450 nm using a multiwell plate reader. The concentration of each cytokine was calculated based on the calibration curves.
Statistical Analysis
Data were presented as the mean ± standard divergence. Statistical analysis was performed using a SPSS 13.0 software package. The quantitative data were compared using one-way analysis of variance and least significant difference (LSD) method. The correlation among Fas expression, mRNA expression of 4 apoptosis-related genes, and apoptotic rates were measured using linear regression analysis. A p-value < 0.05 was considered statistically significant.
Results
Amplification of CIK and Purification of the CD4+ CIKs
In these 15 patients, the average of CIKs after 14–16 days of amplification reached 40–78 × 108 (median 69.1 × 108). The purity of CD4+ CIKs or the CD4+ PBMCs isolated by the magnetic separation system was 93.2% ± 4.7% with vitality higher than 95% based on trypan blue staining.
Expression of CD40L and Secretion of IFN-γ Increased in CD4+CIK
In order to examine different cytokines secreted by CD4+ CIKs, the supernatants of purified CD4+ CIKs and CD4+ PBMCs were collected after 4- and 24-hour culture in blank serum-free medium X-VIVO 20 without any extrinsic cytokine supplement. The concentrations of IL-2, IFN-γ, TNF-α, and sCD40L were measured. The levels of 4 cytokines dramatically increased in the supernatants of CD4+ CIKs following either 4 or 24 hours of culture. However, the concentrations of IFN-γ and sCD40L were remarkably increased in the supernatants of CD4+ CIKs after 24 hours of culturing compared with CD4+ PBMCs, which increased from 192.82 ± 123.03 and 230.28 ± 127.58, to 2203.79 ± 237.56 and 1934.57 ± 436.99, with p value < 0.01, respectively (Table 2).
Comparison of Cytokines Secreted by CIKs and PBMCs (n = 7, 5 × 105 cells, pg/mL)
CIK, cytokine-induced killer cell; PBMC, peripheral mononuclear cell; IL-2, interleukin-2; IFN-γ, interferon-γ; TNF-α, tumor necrosis factor-α.
The sCD40L is the sheared extramembrane domain of human CD40L that is capable of performing active functions similar to the membrane-attached CD40L. Therefore, the expression of full-length CD40L at both mRNA and protein levels were detected by semiquantitative RT-PCR and flow cytometry. Compared with the CD4+ PBMCs isolated from the same patients, a significant increase was identified not only at the level of mRNA synthesis but also in the proportion of membrane-conjuncted CD40L positive cells, which elevated from 1.58% ± 0.53% to 25.51% ± 4.23% (Fig. 1A–D).
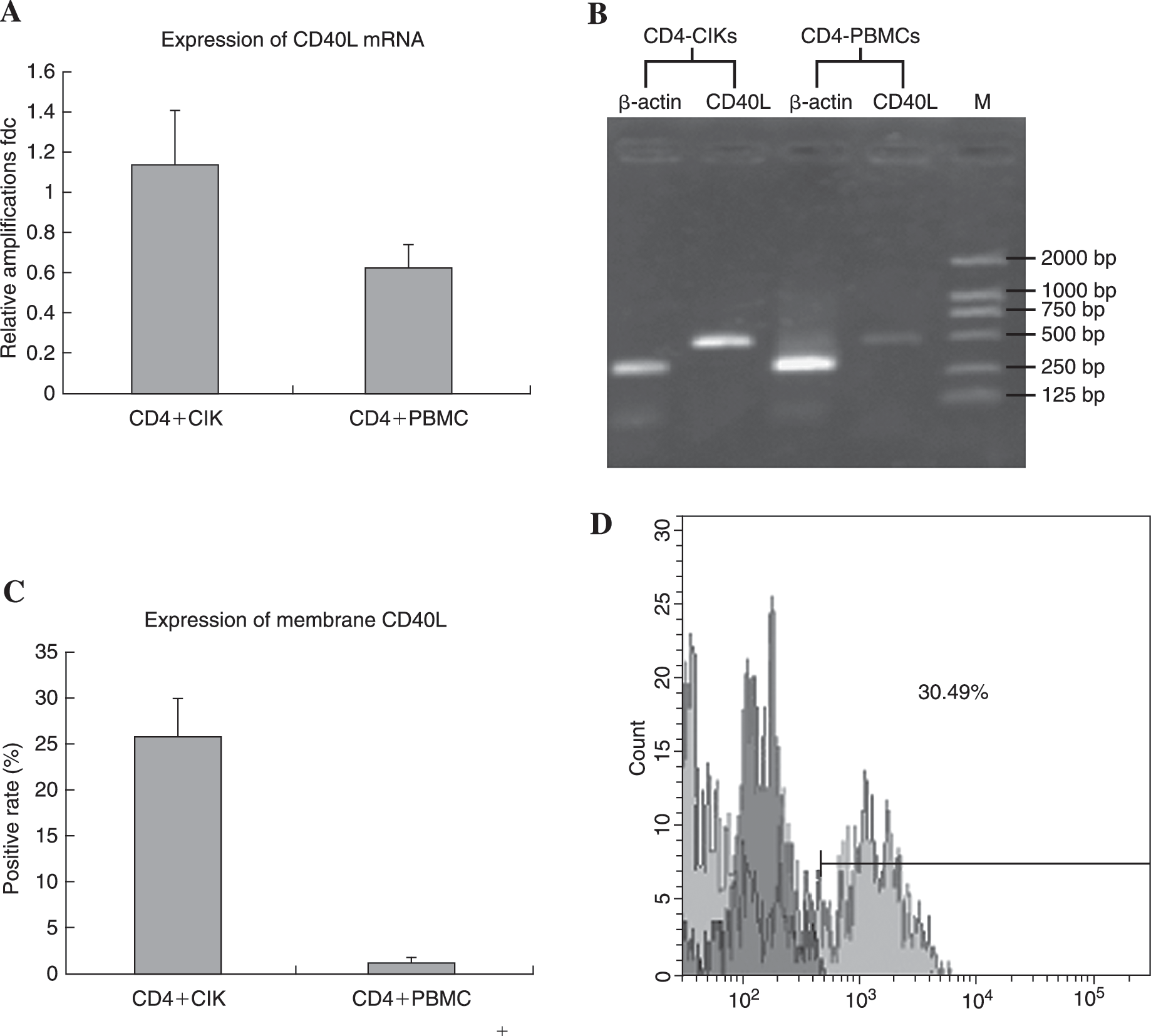
The increased expression of CD40L in CD4+ cytokine-induced killer cells (CIKs) was identified at mRNA level and protein level using semiquantitative reverse-transcriptase polymerase chain reaction (RT-PCR), flow cytometry, and enzyme-linked immunosorbent assay (ELISA) analysis. Compared with CD4+ peripheral mononuclear cells (PBMCs), the mRNA synthesis increased twofold after calibration against the housekeeping gene β-actin. Because the CD40L protein included 2 types, we detected membrane type and soluble type separately. The membrane-attached CD40L on CD4+ CIKs gated by phycoerythrin (PE)-Cy5-labeled anti-CD4 monoclonal antibody (McAb) was detected using fluorescein-isothiocyan (FITC)-labeled anti-CD40L McAb. After 12–14 days of culturing in the presence of multiple cytokines, the rates of CD40L-positive cells increased significantly from 1.58% ± 0.53% in CD4+ PBMCs to 25.5% ± 4.23% in CD4+ CIKs, with p < 0.01. The soluble CD40L (sCD40L) remarkably increased in supernatants of CD4+ CIKs as detected by specific human sCD40L ELISA kit (see Table 2).(
Susceptibility to Fas-Mediated Apoptosis of Insensitive MDA-MB-231 Cells Increased after Pretreatment with Supernatants from CD4+ CIKs
The expression of Fas on T47D cells and MDA-MB-231 cells are 21.4% and 3.9%, respectively. Therefore, the sensitivity of these 2 breast cancer cell lines to Fas-mediated apoptosis is variable. Contrary to highly sensitive T47D cells, the MDA-MB-231 cells are resistant to Fas and none of the Annexin-V-positive apoptotic cells were observed when MDA-MB-231 cells were cocultured with activated anti-Fas McAb CH11 (Fig. 2B). But when these resistant breast cancer cells were pretreated with the supernatant of purified CD4+ CIKs for 24 hours, an obvious apoptosis was identified in almost a quarter of total MDA-MB-231 cells after CH11 was added. These results implied that something in supernatants of CD4+ CIKs altered sensitivity of MDA-MB-231 cells to Fas-mediated apoptosis (Fig. 2C).
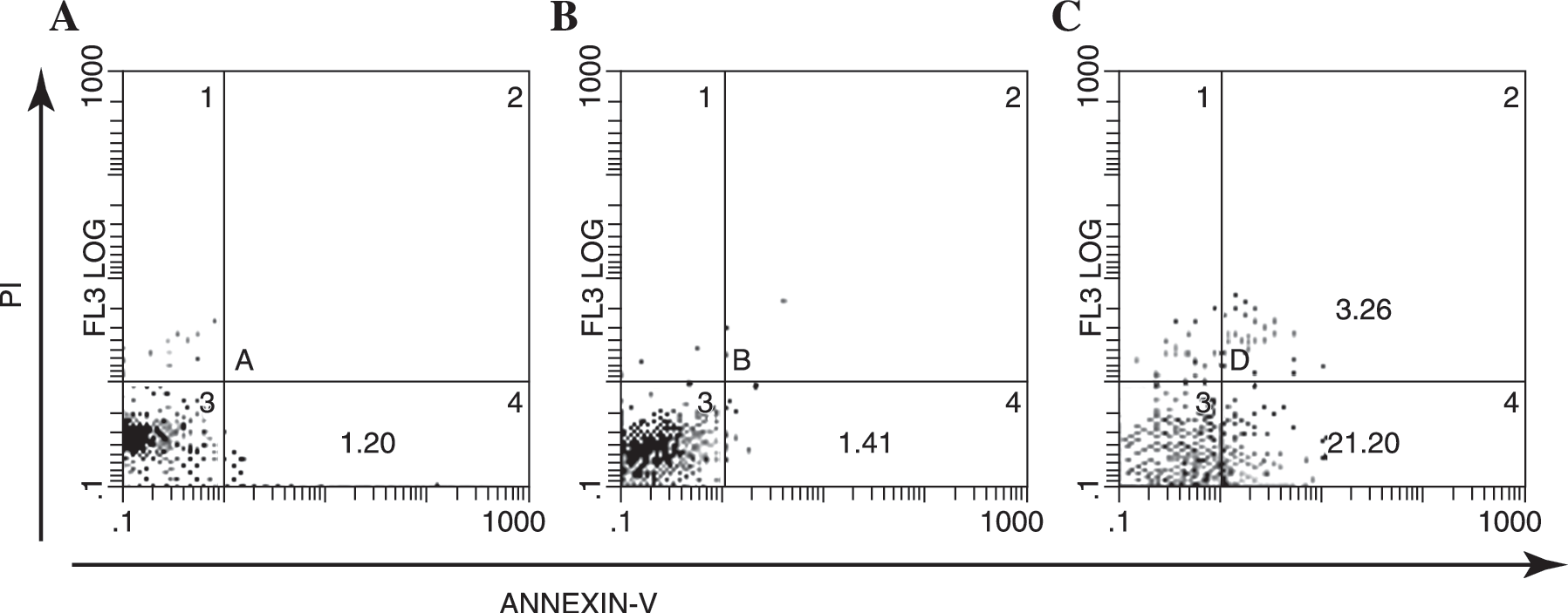
The resistance to Fas-mediated apoptosis of MDA-MB-231 cells was reverted after pretreated with the supernatants of purified CD4+ cytokine-induced killer cells (CIKs) for 24 hours. The apoptotic cells were detected by Annexin-V staining using flow cytometry. Less than 4% of MDA-MB-231 cells expressed detectable levels of Fas on the surface; therefore, none of the Annexin-V-positive apoptotic cells were observed when MDA-MB-231 cells were cocultured with activated anti-Fas monoclonal antibody (McAb) CH11. But when these resistant breast cancer cells were pretreated with the supernatant of CD4+ CIKs for 24 hours, apoptosis was clearly identified in more than 20% of MDA-MB-231 cells after addition of CH11. (
Apoptosis of MDA-MB-231 Cells Induced by CD4+ CIK Correlated with Upregulated Expression of Fas
In order to verify the effects of CD4+ CIKs on reversion of Fas-resistance of MDA-MB-231 cells, T47D and MDA-MB-231 cells were cocultured with CD4+ CIKs, respectively, for 24 hours at variable ratios. A dose-dependent increase of apoptotic rates was detected both in T47D and MDA-MB-231 cells though the constitutive expression of Fas on surface varied dramatically. Furthermore, increased apoptotic rates observed in the MDA-MB-231 cells positively correlated with the elevated expression of Fas, the same as that observed in the T47D cells (Fig. 3A). After making a statistical analysis between apoptosis rates and Fas expression, distinctly positive correlation between theses 2 variables was verified with statistical significance, which implied that the apoptosis observed in MDA-MB-231 cells induced by CD4+ CIKs was regulated through Fas pathway (Fig. 3B).
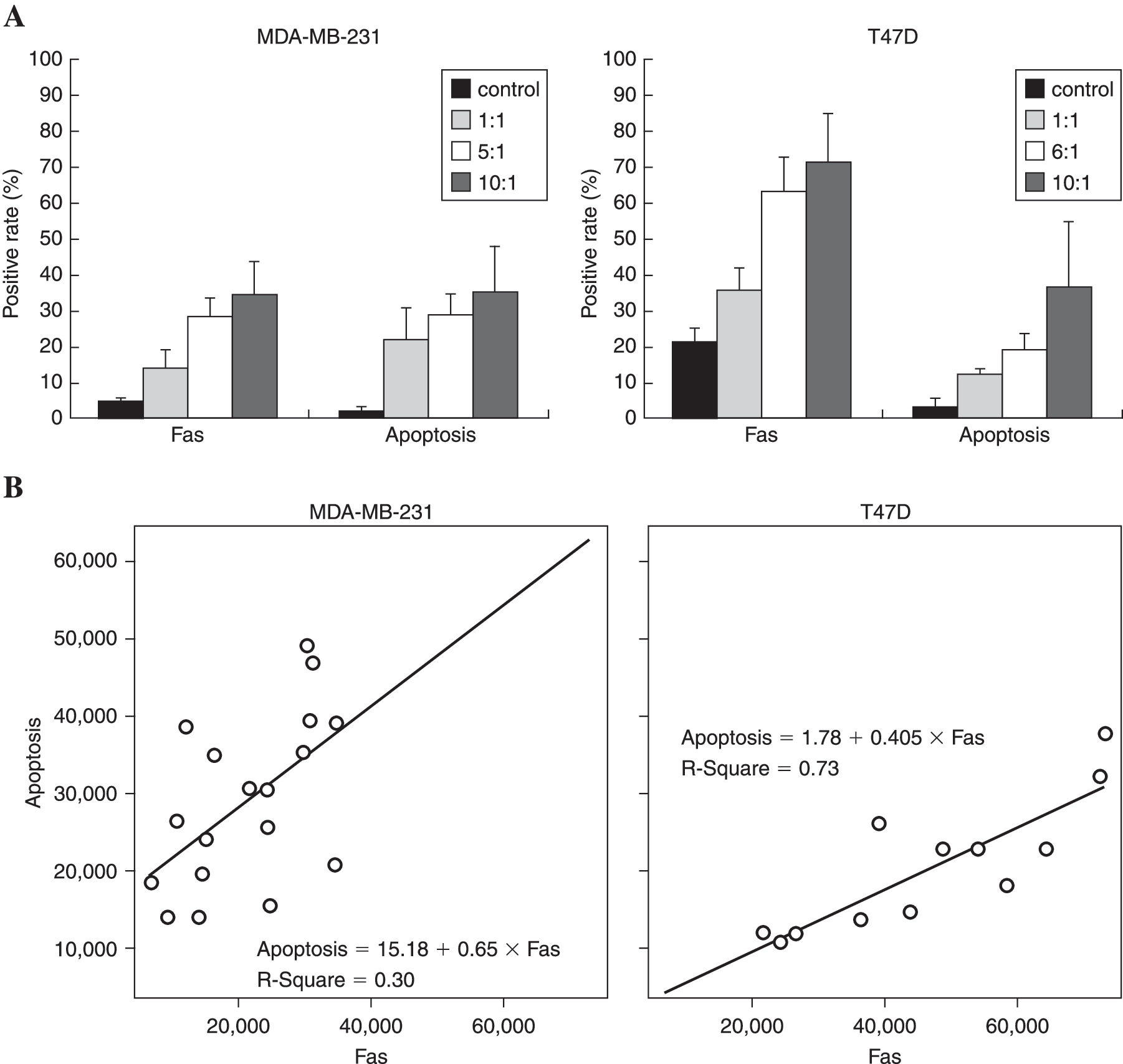
The apoptotic rates and Fas expression of T47D and MDA-MB-231 cells cocultured with CD4+ cytokine-induced killer cells (CIKs) at variable ratios for 24 hours were analyzed using flow cytometry. Higher levels of Fas (21.4%) were expressed on T47D cells than MDA-MB-231 cells. After coculture with CD4+ CIKs, a similar sharp increase of Fas expression in a dose-dependent manner was detected in T47D cells and MDA-MB-231 cells, though the level of FAS expressed on T47D cells was significantly higher than that of MDA-MB-231 cells, 72.97% ± 13.96% and 33.67% ± 9.46% at ratio of 10:1, respectively. Furthermore, the dramatic increase of apoptotic rates were observed in MDA-MB-231 cells as well as in T47D cells that positively correlated with elevated expression of Fas as verified by a linear regression analysis between these 2 variables (p < 0.05). (
Apoptosis of MDA-MB-231 Cells Induced by CD4+ CIK Was Fas-Dependent and Both CD40/CD40L Ligation and IFN-γ Stimulation Participated the Process
In order to determine whether the apoptosis induced by CD4+ CIKs is Fas-dependent, abundantly specific, blocking monoclonal antibody targeted to FasL was added into the coculture system. The anti-FasL McAb ZB4 almost completely eliminated apoptotic MDA-MB-231 cells from the coculture system, which dropped from 33.70% ± 2.77% to 7.57% ± 1.15% (Fig. 4A). This suggested that the apoptosis was Fas-dependent.
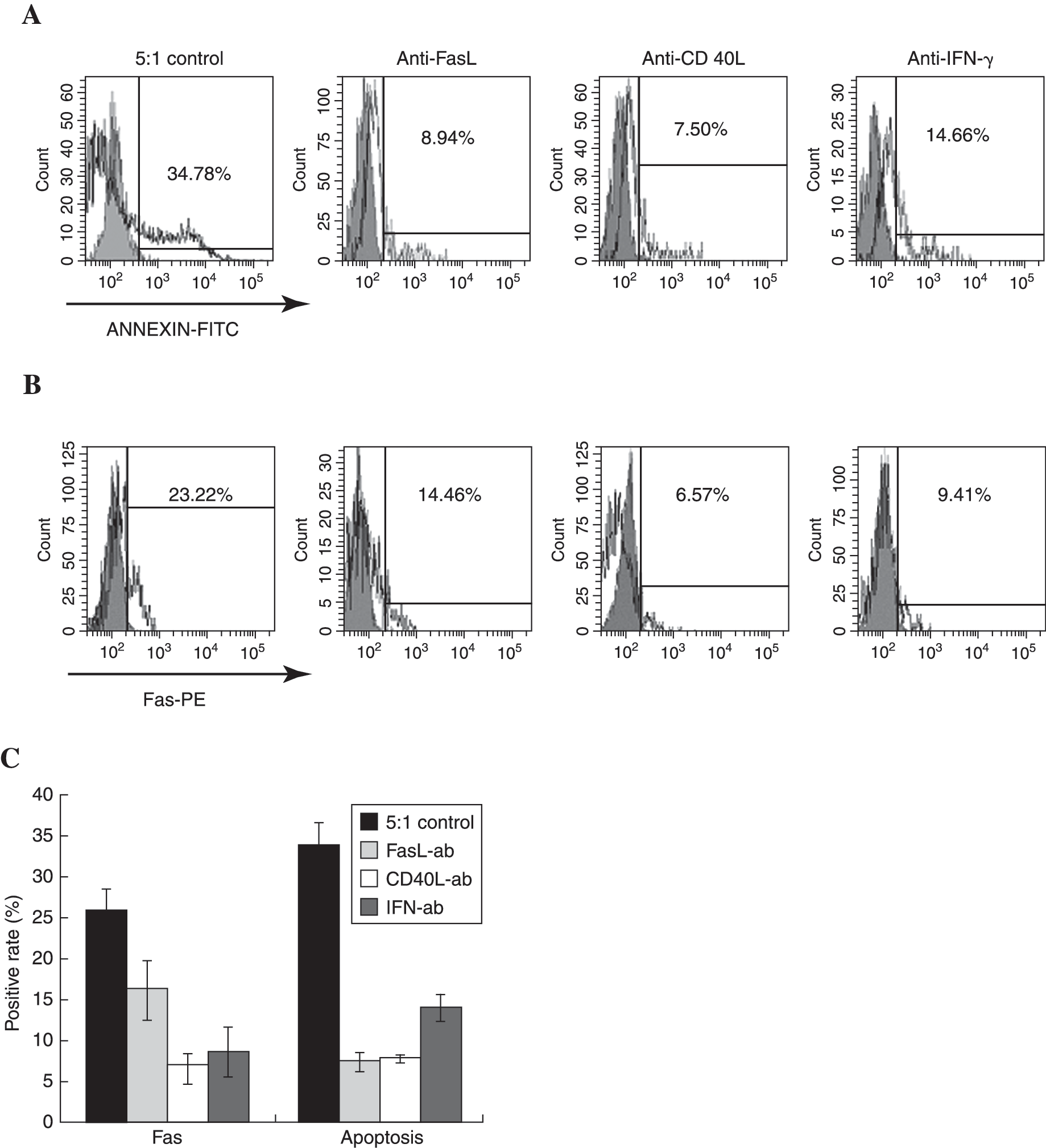
The apoptotic rates and Fas expression of MDA-MB-231 cells cocultured with CD4+ cytokine-induced killer cells (CIKs) at ratio of 1:5 with or without specific blocking antibodies were analyzed. At the beginning of the experiment, abundant blocking antibodies were added into the coculture system and flow cytometry analysis was performed 24 hours later. Compared with considerable apoptosis and enhanced Fas expression detected in the control cells, the anti-FasL McAb ZB4 almost completely eliminated apoptotic MDA-MB-231 cells in the coculture system, from 33.70% ± 2.77% to 7.57% ± 1.15%, though the expression of Fas was inhibited slightly, which dropped from 25.9% ± 2.45% to 16.20% ± 3.38%, both with p < 0.05. Both anti-CD40L monoclonal antibody (McAb) B-B29 and anti-IFN-γ McAb B-B1 effectively blocked the expression of Fas on the surface. However, anti-CD40L McAb B-B29 was more effective in preventing MDA-MB-231 cells from apoptosis induced by CD4+ CIKs than anti-IFN-γ McAb B-B1, with decreased apoptotic rates of 7.80% ± 0.26% and 14.20% ± 1.70%, respectively (p < 0.05). (
As mentioned above, both activation of CD40 and high level of IFN-γ stimulation had the ability to increase expression of Fas on tumor cells. Because the reversible resistance of MDA-MB-231 cells to Fas-induced apoptosis depended on variable expression of competent Fas protein on membrane, to discriminate which is the major promoter of Fas expression will help explain the inducible reversion. In this study, abundantly blocking monoclonal antibodies specific to CD40L and IFN-γ were added separately. Both anti-CD40L McAb and anti-IFN-γ McAb effectively decreased the expression of Fas on the surface in a similar manner, which dropped from 25.90% ± 2.45% to 6.93% ± 1.56%, and 8.73% ± 2.93%, respectively (Fig. 4B). However, anti-CD40L McAb and anti-IFN-γ McAb were distinguished by notably different potentials in inhibition of apoptosis. The anti-CD40L McAb protected more than 75% of MDA-MB-231 cells from apoptosis, much higher than the anti-IFN-γ McAb, and almost as powerful as the anti-FasL McAb (Fig. 4C). These results demonstrated that both CD40/CD40L ligation and IFN-γ stimulation participated in the process of apoptosis by upregulating the expression of Fas on the surface. However, it was interesting that only interruption of CD40/CD40L ligation rather than extermination of IFN-γ almost eliminated Fas-dependent apoptosis of MDA-MB-231 cells induced by CD4+ CIK, though both CD40/CD40L and IFN-γ blocked the interaction between Fas and FasL. It is acceptable that other factors in MDA-MB-231 cells, which amplified the apoptotic signals downstream of Fas engagement, emerged or activated after CD40 activation rather than IFN-γ stimulation.
Downregulated Expression of c-FLIP Rather Than Bcl-2 Coincided with the Early Stage of Apoptosis in MDA-MB-231 Cells
In order to determine the disparity between CD40/CD40L ligation and IFN-γ stimulation on MBA-231 cells, the expression of 4 proapoptotic and antiapoptotic genes (FADD, c-FLIP, Bax, and Bcl-2), which represent both extrinsic and intrinsic pathways involved in Fas-mediated apoptosis, were analyzed at the mRNA level. The total RNA was extracted from MDA-MB-231 cells alone or cocultured with CD4+ CIK for 4, 6, and 24 hours, respectively. Though no significant difference was found in Bax and FADD, the transient downregulated expression of Bcl-2 and c-FLIP was identified 6 hours after coculture, followed by a rapid elevation exceeding the baseline levels at 24 hours after coculture (Fig. 5A). Therefore, the ratio of Bax/Bcl-2 increased from 27.02 ± 6.39 to 54.17 ± 10.82, followed by a decrease to 14.57 ± 1.74). The ratio of FADD/c-FLIP also increased from 1.21 ± 0.38 to 3.84 ± 0.55, and then decreased to 0.97 ± 0.15 (Fig. 5B). This result implied that the mitochondrial and extrinsic Fas/FasL pathways were both involved in inducible Fas-dependent apoptosis of MDA-MB-231 cells by CD4+ CIK. However, the regression analysis showed that downregulation of c-FLIP rather than Bcl-2 significantly correlated with the early stages of apoptosis observed in MDA-MB-231 cells with the standardized correlation coefficient R 2 of c-FLIP equal to 0.806.
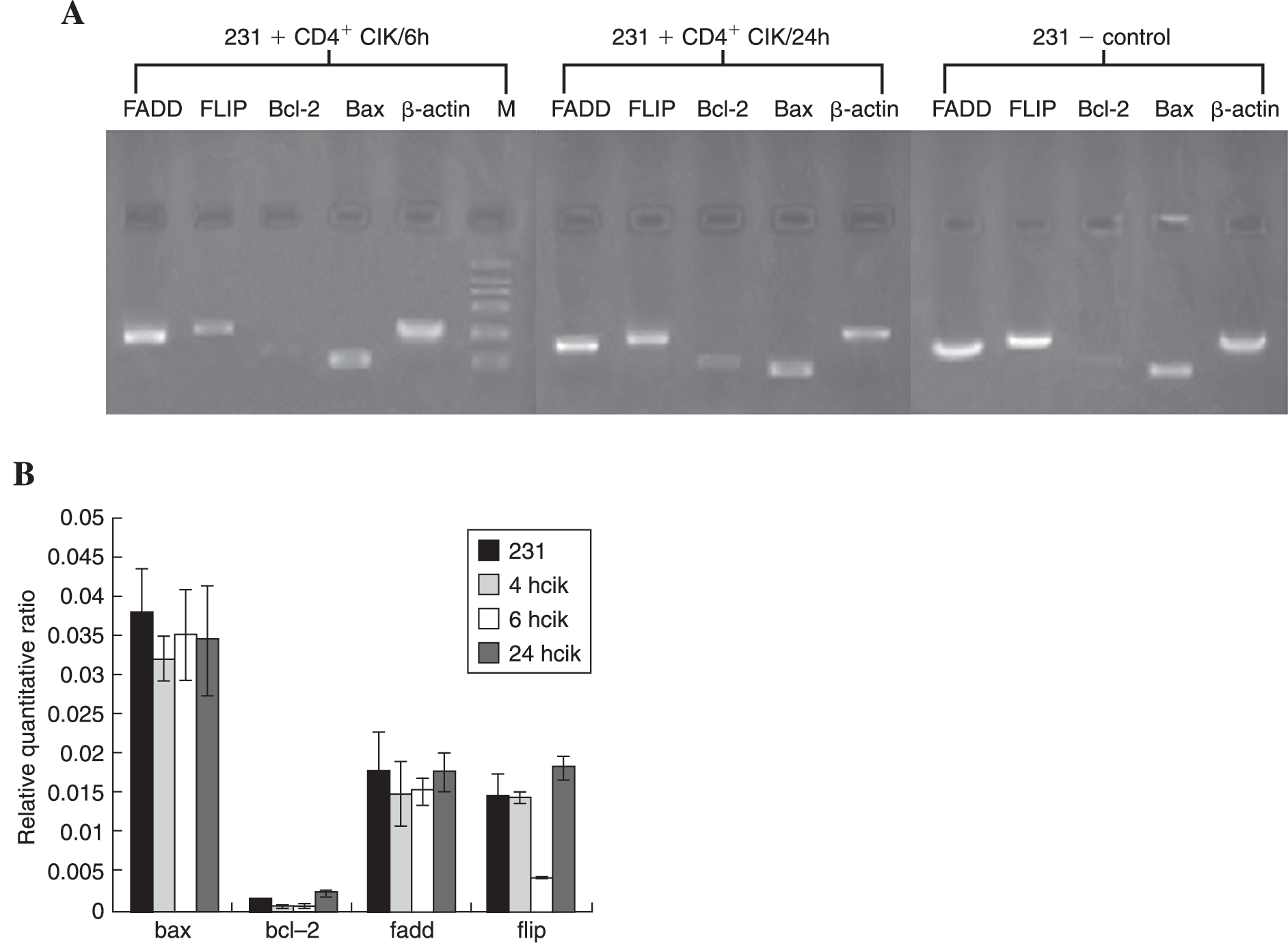
The expression of 4 apoptosis-related genes, including FADD, c-FLIP, Bax, and Bcl-2, in MDA-MB-231 cells alone or cocultured with CD4+ cytokine-induced killer cells (CIKs) for different periods of time were analyzed at the mRNA level using real-time quantitative reverse-transcriptase polymerase chain reaction (RT-PCR). Compared with the normal MDA-MB-231 cells, transient downregulation of mRNA synthesis of Bcl-2 and c-FLIP were identified at 6 hours after coculture, which decreased 2.18 times and 3.65 times, respectively (both p < 0.01). Thereafter, rapid elevation of 2 genes exceeding the baseline levels were followed until 24 hours after coculture, which increased 3.65 times and 4.59 times, respectively (both p < 0.001). No significant difference was found in Bax and FADD (p > 0.05). Therefore, the ratio of Bax/Bcl-2 increased from 27.02 ± 6.39 to 54.17 ± 10.82, then decreased to 14.57 ± 1.74, while the ratio of FADD/c-FLIP also increased from 1.21 ± 0.38 to 3.84 ± 0.55, then decreased to 0.97 ± 0.15. However, the regression analysis showed that downregulation of c-FLIP rather than Bcl-2 correlated with apoptosis of MDA-MB-231 cells induced by CD4+ CIKs significantly, and the standardized correlation coefficients R
2 of c-FLIP and Bcl-2 were 0.806 and 0.274, with p values equaled to 0.002 and 0.228, respectively. (
Contrary Effect of CD40/CD40L Ligation and IFN-γ Stimulation on Synthesis of c-FLIP though Both of Them Significantly Upregulated Synthesis of FADD, Bax, and Bcl-2
Abundant blocking antibodies against CD40L and IFN-γ were added into coculture systems at 6 and 24 hours. The similar potent inhibition of Bax, Bcl-2, and FADD was observed at 6 hours, which implied that both CD40/CD40L ligation and IFN-γ stimulation could increase mRNA synthesis of Bax, Bcl-2, and FADD. However, contrary effects were verified on synthesis of c-FLIP, which CD40/CD40L ligation significantly inhibited while IFN-γ stimulation promoted the mRNA expression (Fig. 6A). The disparity of CD40/CD40L ligation and IFN-γ stimulation were only detected during early stages of coculture, for no difference in the synthesis of 4 genes was identified 24 hours after incubation (Fig. 6B). The phenomenon coincided with detectable early stages of apoptosis determined by Annexin-V staining, which could be detected under fluorescent microscopy early at 4 hours and culminated at 24 hours of incubation with CD4+ CIKs. These results implied that CD40/CD40L ligation significantly promoted intrinsic downstream apoptotic signal transferring after Fas engagement by simultaneously inhibiting expression of the antiapoptotic gene c-FLIP and stimulating expression of proapoptotic gene FADD, resulting in significant difference between CD40/CD40L ligation and IFN-γ stimulation in reversing Fas-resistance of MDA-MB-231.
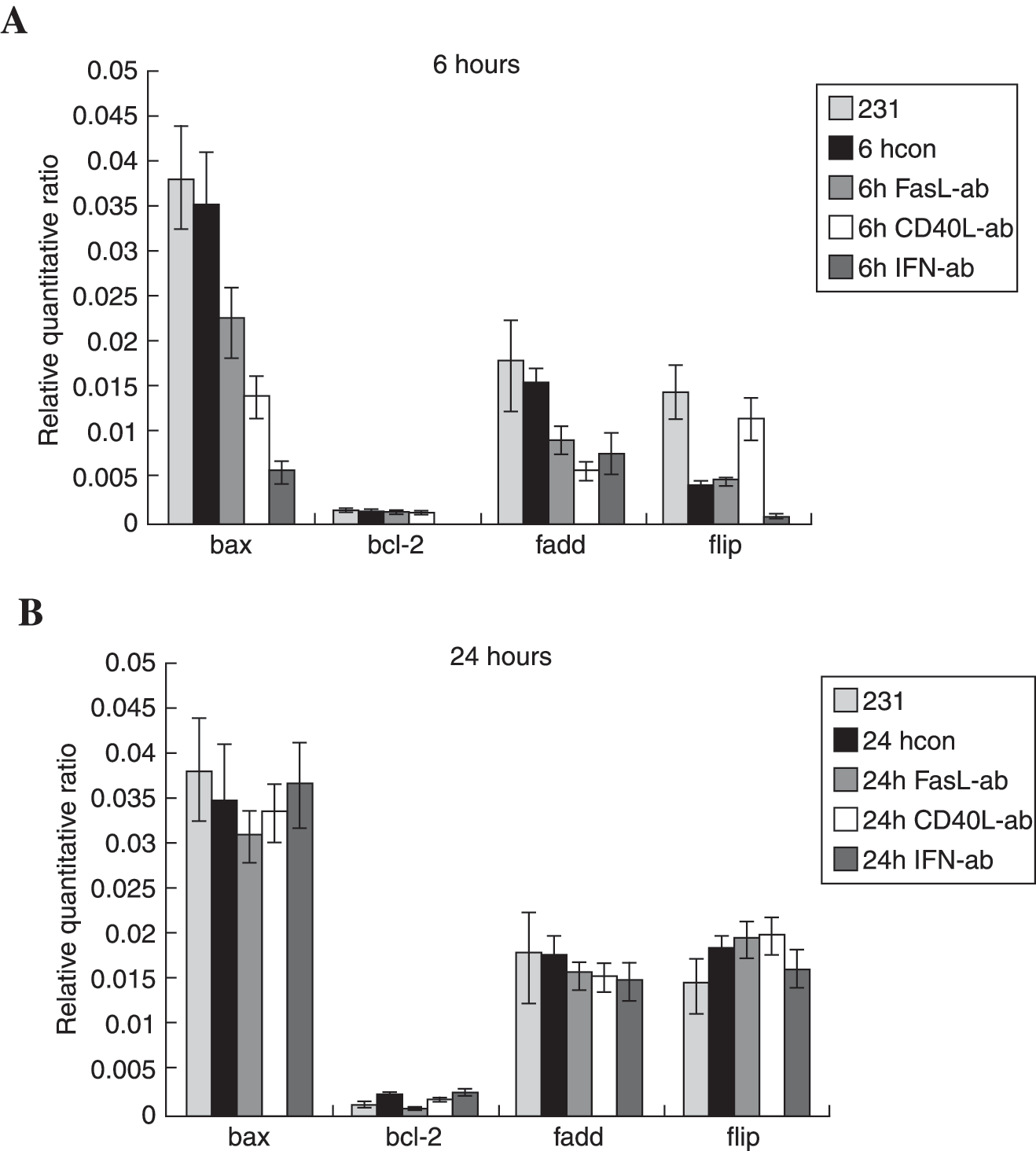
The levels of mRNA expression of 4 apoptosis-related genes in MDA-MB-231 cells were compared at either 6 or 24 hours after coculture with CD4+ cytokine-induced killer cells (CIKs) with or without specific blocking antibodies. Abundant blocking antibodies against FasL, CD40L, and interferon-γ (IFN-γ) were added into the coculture systems. The sharply fluctuating quantity of all these 4 genes were demonstrated at 6 rather than 24 hours, regardless of which antibody was used. The same potent inhibition of mRNA synthesis of Bax, Bcl-2, and FADD were observed using anti-FasL, anti-CD40L, or anti-IFN-γ monoclonal antibodies (McAbs), except c-FLIP. As displayed, anti-CD40L neutralizing antibody significantly increased the mRNA synthesis of c-FLIP about threefold, which was contrary to the inhibitory effects of anti-FasL and anti-IFN-γ McAbs. These results implied that absolutely contradictory effects on c-FLIP produced the largest difference between CD40/CD40L ligation and IFN-γ stimulation in reversing Fas-resistance of MDA-MB-231, though both of them increased expression of Bax, Bcl-2, and FADD. The CD40/CD40L ligation significantly promoted the intrinsic downstream apoptotic signal transferring after Fas engagement by inhibiting expression of antiapoptotic gene c-FLIP and stimulating expression of proapoptotic gene FADD simultaneously. (
Discussion
Compared with traditional LAK and TIL treatments, CIKs have unique advantages, including enhancement of cell proliferation and cytotoxic activity, enlargement of the antitumor spectrum, more sensitivity to drug-resistant tumor cell lines, resistance to Fas-mediated apoptosis, and lower hematopoietic toxicity. 11 –13 CIKs are heterogeneous cells, and the major population (about 90%) was CD3+ T cells, in which CD8+ T cells and CD4+ T cells each held half. In our previous studies, we have identified that CD4+ CIKs are a distinctly critical subset that is competent to produce considerable antitumor effects by inducing Fas-dependent apoptosis in tumor and producing Th1 dominance cytokines. 4,5 In addition, CD40/CD40L ligation and high levels of IFN-γ in the supernatant of CD4+ CIKs seemed to participate partly in the apoptosis progress. However, Fas-dependent apoptosis was detected in Raji cells that were sensitive to Fas activation. As reported, FasL is highly expressed on CIKs. 13 Therefore, CD4+ CIKs should easily trigger a typical apoptosis in Raji through the Fas/FasL pathway, and other proapoptotic factors such as CD40L and IFN-γ may be ignored or even overlapped.
IFN-γ is produced mainly by immune competent cells and exhibits an important antitumor effect. IFN-γ activates cells of the innate immune system to proliferate, produce cytokines, and lyse tumor cells. IFN-γ promotes the development of a Th1 antitumor response and furthermore exerts direct antiproliferative and proapoptotic effects on a variety of tumor cells. 14,15 Furthermore, it was reported that high levels of IFN-γ stimulation increased the levels of CD40 and Fas on the cell surface. 16 CD40 is a transmembrane glycoprotein receptor belonging to the tumor necrosis factor (TNF) receptor superfamily, which is considered a survival factor, triggering normal B-cell growth, survival, and differentiation. 17 CD40L is mainly expressed on activated T cells, especially activated CD4+ T cells, which represent the majority of CD4+ CIKs. Contradictory cellular fates in response to CD40 activation have been reported in different cell types. 18 For example, CD40/CD40L ligation provides an antiapoptotic signal for normal B cells, yet results in growth inhibition of some malignant epithelial cell lines, such as lymphoma, multiple myeloma, malignant melanoma, bladder carcinoma, pancreatic carcinoma, and breast carcinoma, and further sensitizes them to apoptosis. 19 –22 So in this study, we selected a Fas-resistant breast cancer cell line, MDA-MB-231, which expresses small amounts of Fas protein but high levels of CD40 protein as demonstrated by us and other researchers. 23,24 In this cell line, the direct coreaction between Fas and FasL was interrupted, and the proapoptotic effects of other factors that were capable of inducing apoptosis through Fas/FasL pathway indirectly were distinguished.
The resistance of MDA-MB-231 cells to Fas-mediated apoptosis is due to a downregulation of membrane Fas, but when pretreated with the supernatant of CD4+ CIKs for 24 hours, increased apoptosis induced by agonist anti-Fas McAb (CH11) was observed. Moreover, the apoptosis in a dose-dependent manner correlating with up-regulation of Fas on the surface was detected in MDA-MB-231 cells incubated with CD4+ CIKs and blocked by anti-FasL McAb. These results suggested that the apoptosis observed in MDA-MB-231 cells was Fas-dependent and promoted by some factors secreted by CD4+ CIKs. The elevated expression of membrane and soluble CD40L, together with increased secretion of IFN-γ, was certified in the CD4+ CIK. Because T47D cells and MDA-MB-231 cells are both CD40 high-expressing tumor cells, it was supposed that the CD40/CD40L ligation and IFN-γ might participate in the process of Fas-dependent apoptosis induced by CD4+ CIK. Therefore, the anti-CD40L McAb and anti-IFN-γ McAb were added into the coculture system, and both effectively restrained the expression of Fas on the surface and protected MDA-MB-231 cells from apoptosis. However, the anti-CD40L McAb protected more MDA-MB-231 cells from apoptosis than the anti-IFN-γ McAb did. The anti-CD40L McAb was almost as powerful as the anti-FasL McAb in preventing apoptosis. These data suggested that apart from upregulating the expression of Fas on the surface, there were other proapoptotic molecular mechanisms involved to explain the great discrepancy between CD40/CD40L ligation and IFN-γ stimulation in reversion of Fas-resistance of MDA-MB-231 cells.
Upregulation of Fas does not necessarily sensitize cells to apoptosis. The variable changes of proapoptotic and antiapoptotic proteins during the extrinsic death signal transfer inside the cells will determine cell fate, whether apoptosis occurs or not. Clustering of Fas protein on the membrane triggered by its natural ligand FasL or cross-linked with anti-Fas agonist antibody CH11 induced recruitment of the FADD-containing protein to the trimerized intracellular death domain, which in turn recruited the FADD-like IL-1β converting enzyme (FLICE) and led to the formation of death-inducing signaling complex (DISC). 25 After the formation of the DISC, caspase-8 was activated and a cascade of caspases leading to apoptosis was initiated. 26 This process can be interfered by c-FLIP, an analog of caspase-8 without protease activity. If the amount of DISC is deficient, the second Fas-independent intrinsic apoptotic pathway will be activated, resulting in cytochrome C release from the mitochondria to the cytoplasm, and will be modulated directly by the members of the Bcl-2 family genes, including proapoptotic gene Bax and antiapoptotic gene Bcl-2. 27,28
To investigate the apoptosis regulatory molecules involved in MDA-MB-231 cells induced by CD4+ CIKs, 4 pivotal proapoptotic and antiapoptotic genes, including FADD, Bax, c-FLIP, and Bcl-2, were detected by real-time quantitative RT-PCR analysis. All genes were strongly expressed in MDA-MB-231 cells besides Bcl-2. After coculture with CD4+ CIKs, downregulation of mRNA levels of c-FLIP and Bcl-2 occurred in a time-dependent manner and recovered after 24 hours. The downregulation of c-FLIP rather than Bcl-2 significantly correlated with the early stages of apoptosis in MDA-MB-231 cells according to the linear regression analysis. Both CD40/CD40L ligation and IFN-γ stimulation had the capability to upregulate Fas levels on the membrane. Two neutralizing antibodies, anti-CD40L McAb and anti-IFN-γ McAb, were used to distinguish which was more critical for apoptosis of MDA-MB-231 cells. At 6 hours after incubation, the expression of proapoptotic genes FADD and Bax, as well as antiapoptotic gene Bcl-2 decreased significantly either by anti-CD40L McAb or by anti-IFN-γ McAb, which implied that both CD40/CD40L ligation and IFN-γ stimulation could increase the synthesis and expression of Bax, Bcl-2, and FADD. However, the contrary effects on expression of another antiapoptotic gene, c-FLIP, were verified in that CD40/CD40L ligation was significantly inhibited while IFN-γ stimulation promoted the mRNA expression. These results implied that CD40/CD40L ligation rather than IFN-γ stimulation was the essential factor that triggered the apoptosis in MDA-MB-231 cells by downregulating transcription of c-FLIP, though both significantly increased the membrane Fas protein. Our results are consistent with the findings of other researchers 14,17 that IFN-γ stimulation is not an indispensable factor for CD40/CD40L ligation-induced growth inhibition and apoptosis, although an enhancing effect cannot be excluded.
Footnotes
Acknowledgments
We thank Prof. Pengbo Zhou of pathology and laboratory medicine at the Weill Medical College of Cornell University for providing help and assistance for this article. This work was supported by grants from Tianjin Nature Science Foundation (06FZZDSF01500).