
Abstract
Many approaches targeting MUC1 for breast tumor immunotherapy have been attempted. However, preclinical trials with MUC1 showed that MUC1 is a relatively poor immunogen in human. B7 molecules that bind CD28 provide an antigen-nonspecific signal, which, along with an antigen-specific signal, is crucial for T-cell activation. In the present study, we constructed a novel Bacillus Calmette-Guérin-based breast cancer vaccine that coexpressed four VNTRs (variable-number tandem repeats) of MUC1 and CD80 (rBCG-MVNTR4-CD80). The aim of our study was to enhance anti-MUC1 tumor immunity by vaccination of hu-PBL-SCID mice with the recombinant BCG vaccine. The inhibition effect on tumors from the mice immunized with rBCG-MVNTR4-CD80 significantly increased, compared with rBCG-MVNTR4, BCG-pDE22, and phosphate-buffered saline immunized mice (p < 0.05, p < 0.05, p < 0.05). ELISpot assays showed that there was a significant increase in interferon-gamma production in the splenocytes from the mice immunized with rBCG-MVNTR4-CD80. In addition, CD4 and CD8-positive lymphocytes in tumors from rBCG-MVNTR4-CD80-immunized animals were detected. These data showed that rBCG-MVNTR4-CD80 immunization elicited tumor-specific immune response, which closely related with the B7 molecule (CD80), indicating that the vaccine may be a good candidate for MUC1-positive breast cancer immunotherapy.
Introduction
MUC1 is a type I transmembrane glycoprotein expressed by human epithelial cells; it is composed of a variable number (20–125) of 20 amino-acid variable-number tandem repeats (VNTRs) and is heavily glycosylated. 1 –3 MUC1 is expressed on a variety of epithelial-derived cells, including breast ductal cells. Many of the approaches targeting MUC1 for breast tumor immunotherapy have been attempted. However, preclinical trials with MUC1 showed that MUC1 is a relatively poor immunogen in human. A possible reason for this lack of immunogenicity is that MUC1 is a self-antigen, and the immune system has safety mechanisms for preventing self-tissue antigen recognition and autoimmune responses.
The development of an anti-MUC1 vaccine that can successfully induce antitumor cellular immune responses capable of leading to tumor rejection requires the “breaking” of immune tolerance to the molecule. 4 It is now established that members of the B7 family ligands play a central role in the positive and negative regulation of antigen-specific T-cell-mediated immune responses. B7 molecules that bind CD28 provide an antigen-nonspecific signal, which, along with an antigen-specific signal, is crucial for T-cell activation. 5 –7 Recognition of a foreign epitope in conjunction with MHCI/II by the antigen-specific T-cell receptor without additional costimulation provided by B7/CD28 association results in T-cell anergy. 5,8,9 Actually, the expression of B7 resulted in an enhancement of cellular immune responses to different tumor antigens. 10,11
Studies have demonstrated that MUC1-specific immunodominant epitopes are derived from the VNTR region. 12 –14 Consequently, immunotherapy that targets multiple tandem repeats of MUC1 may enhance immune responses against MUC1-expressing cancer cells.
In the present study, we constructed a novel Bacillus Calmette-Guérin-based breast cancer vaccine that coexpressed multiple tandem repeats of MUC1 and CD80. The effect of the vaccine on the growth of breast tumor on hu-PBL-SCID mice was evaluated. The results showed that the vaccine induced antitumor cellular immune responses.
Materials and Methods
Bacterial strains, vectors, animals, and reagents
The BCG vaccine strain was obtained from the Lanzhou Bioethical Production Institute (Lanzhou, China). The pDE22 is a shuttle plasmid capable of replicating and expressing recombinant proteins in Escherichia coli and BCG, maintained in the Laboratory Animal Research Center of the Fourth Military Medical University (Xi'an, China). 15 Female SCID mice (CB17 SCID; 4–5 weeks old) were purchased from the Shanghai Laboratory Animal Center (Shanghai, China) and maintained in pathogen-free facilities at the State Key Laboratory of Cancer Biology (Shanghai, China) and handled in accord with the Institutional Animal Care and Use Committee guidelines. Anti-MUC1 antibody was purchased from Sigma (St. Louis, MO). Anti-CD80 antibody was purchased from DAKO Corp. (Carpinteria, CA). Complete RPMI-1640 media was prepared by supplementing plain RPMI-1640 with 2% 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), 0.1 mM of 2-mercaptoethanol, 100 U/mL of penicillin, 100 μg/mL of streptomycin, 2 mM of glutamine, and 10% fetal calf serum (FCS). The HSP65-MUC1 fusion protein was stored in a refrigerator in our laboratory center 16 between 2 and 8°C.
Construction of rBCG-MVNTR4 and rBCG-MVNTR4-CD80 strains
One single VNTR sequence of human MUC1 was synthesized by AuGCT Biotechnology (Beijing, China) and cloned into pUC18 plasmid (pUC-MVNTR1). The sequence was designed as follows:
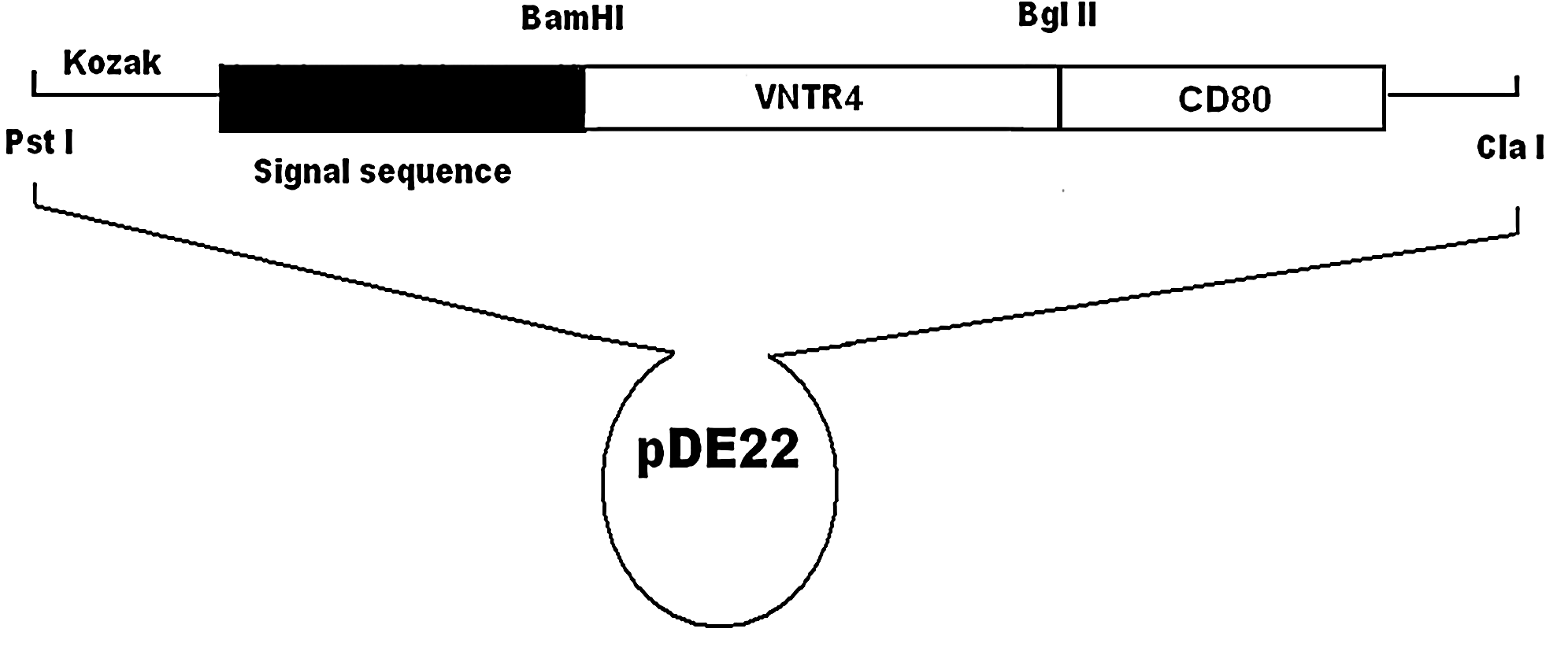
Schematic diagram of pDE-MVNTR4-CD80.
Cell preparation
Two human breast cancer cell lines, MCF-7 and MDA-MB-231 (ATCC), were maintained in our laboratory. MUC1 expression was determined by immunocytochemistry. These cells were trypsinized and incubated with anti-MUC1 antibody (Sigma) at 1:20 dilutions. Breast cancer cells were stained with anti-MUC1 antibody; thereafter, they were incubated with antimouse fluorescence-labeled antibodies.
Reconstitution of SCID mice (hu-PBL-SCID mice)
SCID mice were reconstituted, as previously described. 17 Briefly, female SCID mice were reconstituted with 50 × 106 human PBLs to create a xenograft of human lymphocytes in SCID mice (hu-PBL-SCID mice). PBLs (HLA-A2) were procured from human buffy coats obtained from the Xijing Hospital Blood Center (Xi'an, China). The buffy coats were resuspended in PBS and layered under a high-density solution of Ficoll-Paque PLUS (Amersham Pharmacia Biotech AB, Uppsala, Sweden) and centrifuged for 30 minutes at 1500 rpm. Subsequently, the interface containing PBLs was harvested and washed twice in PBS. The washed cell pellet was resuspended in PBS, and each of the SCID mice was injected intraperitoreally (i.p) with 50 × 106 PBLs.
Therapeutic vaccination studies
The vaccines used in this experiment included rBCG-MVNTR4 [106 colony-forming units (cfu) of experimental vaccine], rBCG-MVNTR4-CD80 (106 cfu of experimental vaccine), BCG-pDE22 (106 cfu; blank vector control), and PBS (sham vaccine) diluted in a total volume of 200 μL of PBS. A total of 8 mice were included in each group. To perform therapeutic studies, 3 days after lymphocyte reconstitution, the mice were subcutaneously (S.C.) injected with 4 × 106 MCF-7 breast cancer cells and subsequently received 2 i.p. vaccine injections at biweekly intervals on days 7 after MCF-7 cell inoculation. Outcomes of interest comprised time to tumor detection, size of primary tumor, and rate of tumor growth. In the case of each experimental group, the median time to tumor onset was determined. For each group, mean tumor-size curve was plotted over time after tumor engraftment. Palpable tumors were measured by calipers, and tumor volumes were calculated by using a standard formula: width (mm)2 × length (mm) × 0.52. Tumor growth was monitored for 70 days after tumor challenge. The mice were observed until impending death or 70 days after tumor inoculation; thereafter, they were sacrificed. All tumor deposits were carefully measured and fixed in a 10% formalin solution. Further, spleen samples were harvested. The harvested tissue was fixed in 10% formalin solution and embedded in paraffin wax. Subsequently, the tissue blocks were sectioned for immunohistochemistry.
IFN-γ-producing cells determined by ELISpot
An interferon-gamma (IFN-γ) ELISpot assay was performed in poly vinylidene fluoride (PVDF)-bottomed 96-well plates (Millipore, Bedford, MA, USA) by using an IFN-γ ELISpot kit (Diaclone, Besancone, France), according to the manufacturer's instructions. Briefly, the ELISpots plates were coated with anti-IFN-γ capture antibody (at 4°C) overnight. After washing three times with PBS, the plates were blocked with 10% FBS-RPMI complete medium for 2 hours at 37°C. Splenocytes from immunized mice were collected and treated with 0.73% NH4Cl for 10 minutes at 37°C to lyse the red blood cells. Next, 5 × 105 splenocytes, in 100-μL medium, were incubated with 20 μg/mL of HSP65-MUC1 fusion protein or media alone for 18 hours at 37°C in IFN-γ ELISpots. Biotinylated anti-IFN-γ antibody (Pharmingen, San Diego, CA) was added and incubated overnight at 4°C. Free antibody was washed out, and the plates were incubated with streptavidin-alkaline phosphatase for 1 hour at 37°C, followed by extensive washing with PBST (PBS + 0.1% Tween20), and with PBS. Spots were visualized by the addition of the alkaline-phosphatase substrate, BCIP/NBT. All wells were examined for positive spots. IFN-γ spot-forming cells were enumerated by using a standard dissection microscope.
Immunohistochemistry of samples
Immunohistochemistry of tissue sections was performed by using the labeled streptavidin biotin (LSAB) + peroxidase kit (DAKO Corp.). Briefly, slides containing the tissue sections were deparaffinized and rehydrated in alcohol and xylene. Endogenous peroxidase activity was suppressed with a 3% hydrogen peroxide solution; further, nonspecific antigen binding was blocked by incubating these sections with swine serum. Subsequently, the tissue was incubated with the appropriate primary antibodies at 4°C overnight. The following Abs were used: anti-MUC1 Ab, antimycobacterial Ab (DAKO Corp.), anti-CD4 Ab (DAKO Corp.), and anti-CD8 Ab. After incubation with the primary Ab, the secondary Ab, namely, streptavidin peroxidase, was added. Following incubation, color development was observed with a substrate chromogen solution (3′, 3′-diaminobenzidine chromogen solution). Subsequently, slides were counterstained with hematoxylin (DAKO Corp.). Finally, slides were dehydrated with alcohol and xylene and mounted for histologic evaluation.
Statistical analysis
Quantitative data are represented as “x ± s (mean ±standard deviation).” One-way analysis of variance (ANOVA) with Bonferroni's post-test was used to compare significant difference of tumor volume between three or more groups; qualitative data were represented as rate (%). Fisher's probabilities test with partitions of the χ2 method were used to compare the significant difference of survival rate between three or more groups. Further, a p-value threshold of ≤0.05 was taken to indicate a statistically significant difference. All statistical analyses were performed with the statistical package for social sciences (SPSS, Inc., Chicago, IL) 16.0 software.
Results
Construction, growth characterization, and expression of rBCG-MVNTR4 and rBCG-MVNTR4-CD80
All recombinant plasmids were identified by restriction enzyme digestion analysis and DNA sequencing. The correct plasmids were then used to transform BCG. After electroporation, colonies were grown on selectable agar media. Growth curves of rBCG-MVNTR4, rBCG-MVNTR4-CD80, and BCG strains (control) showed that there was no difference in the growth patterns among these strains (Fig. 2A). We identified the expression of rBCG-MVNTR4 and rBCG-MVNTR4-CD80 fusion proteins in mycobacteria samples. Further, rBCG-MVNTR4 (14.2 kDa) and rBCG-MVNTR4-CD80 (43 kDa) fusion proteins could be recognized with anti-MUC1 (Fig. 2B) and anti-CD80 Abs (Fig. 2C), respectively.
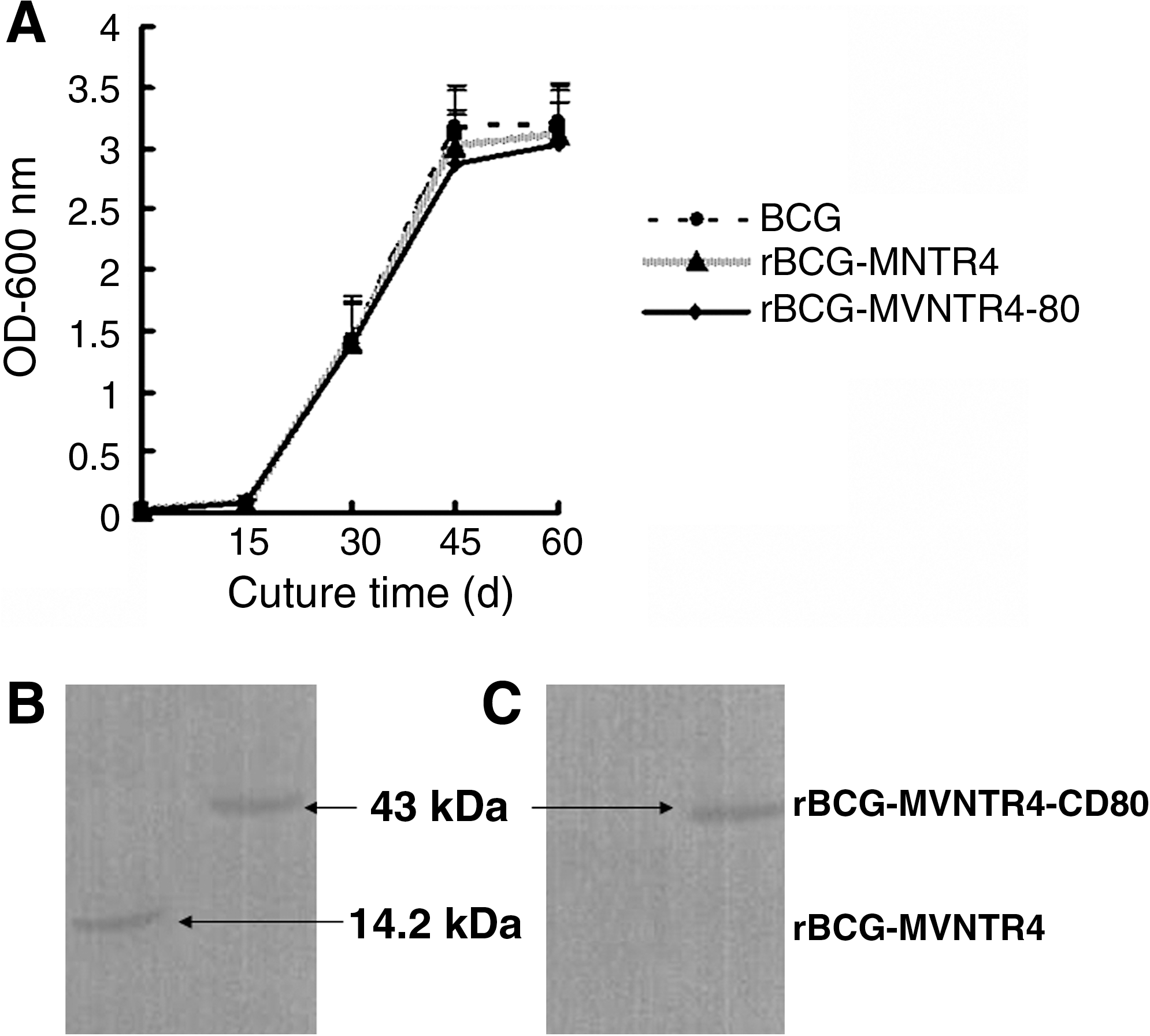
Construction and growth characterization of rBCG-MVNTR4 and rBCGMVNTR4-CD80. (
Therapeutic antitumor effect on mice after immunization of rBCG-MVNTR4 and rBCG-MVNTR4-CD80
The hu-PBL-SCID mice (8 per group) were inoculated with MUC1-positive MCF-7 breast cancer cells (4 × 106) on day 0 and then immunized with rBCG-MVNTR4 and rBCG-MVNTR4-CD80 on days 7 and 21. Control mice were treated with BCG-pDE22 and PBS. Tumor growth was monitored for 70 days after tumor inoculation. As can be seen in Figure 3, tumor growth in the animals immunized with rBCG-MVNTR4-CD80 was significantly inhibited at 42 days, compared with that in the animals immunized with BCG-PDE22 (p < 0.05) and PBS (p < 0.05), whereas tumor growth in the animals immunized with rBCG-MVNTR4 did not show obvious inhibition. Aftert 49 days, tumor growth in the animals immunized with rBCG-MVNTR4-CD80 was significantly inhibited, compared with that in the animals immunized with rBCG-MVNTR4 (p < 0.05) By day 70 post-tumor implantation, 87.5% (7/8) of mice in the PBS control group, 87.5% (7/8) of mice in the BCG-pDE22 control group, and 62.5% (5/8) of mice in the rBCG-MVNTR4 group were dead, whereas only 25% (2/8) of mice in the rBCG-MVNTR4-CD80-treated group were dead, showing a prolonged survival (p < 0.05, versus control group).

Tumor growth in the animals immunized with rBCG-MVNTR4-CD80 was significantly inhibited, compared with that in the animals immunized with rBCGMVNTR4 (p < 0.05), BCG-pDE22 (p < 0.05), and phosphate-buffered saline (p < 0.05).
Induction of MUC1-specific T-cell-mediated immune responses by rBCG-MVNTR4-CD80 immunization
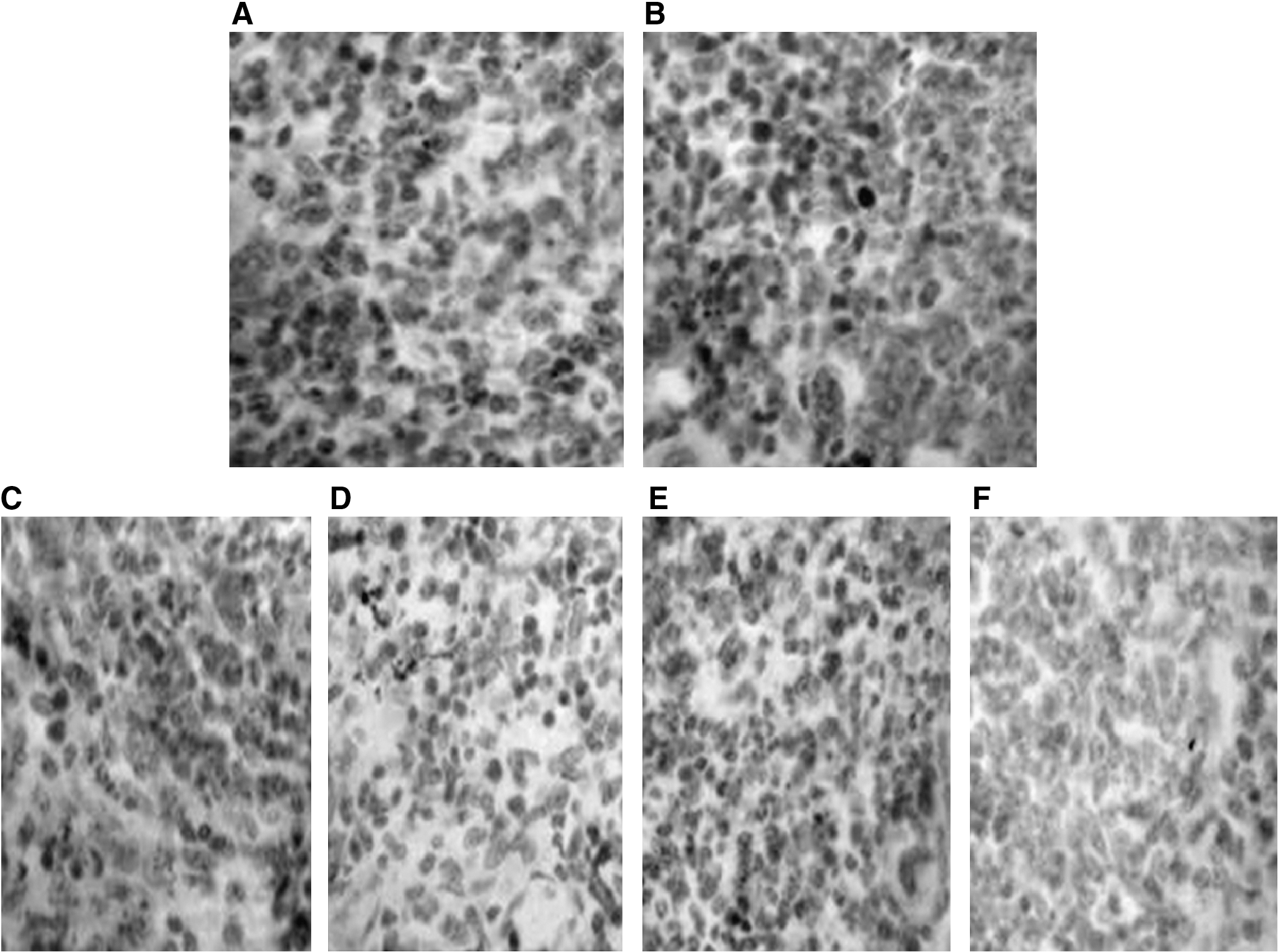
To identify whether the inhibition effect of tumor growth in the animals immunized with rBCG- MVNTR4-CD80 was due to a MUC1-specific T-cell response, IFN-γ ELISpot assays were performed. As can be seen in Figure 4, there was a significant increase in IFN-γ production in the splenocytes from the mice immunized with rBCG-MVNTR4-CD80, compared to the splenocytes from the mice immunized with rBCG-MVNTR4 (p < 0.01) or rBCG-PDE22 (p < 0.01). These results indicated that IFN-γ production was induced by immunization with rBCG-MVNTR4-CD80. To determine whether rBCG-MVNTR4-CD80 was able to induce specific T-cell responses, immunohistochemistry of tumor samples for CD8+/CD4+ human lymphocytes was performed. The results revealed that CD8+ and CD4+ cells in the tumors were detectable (Fig. 5A and 5C); however, these cells could not be detected in control tumors (Fig. 5B and 5D).
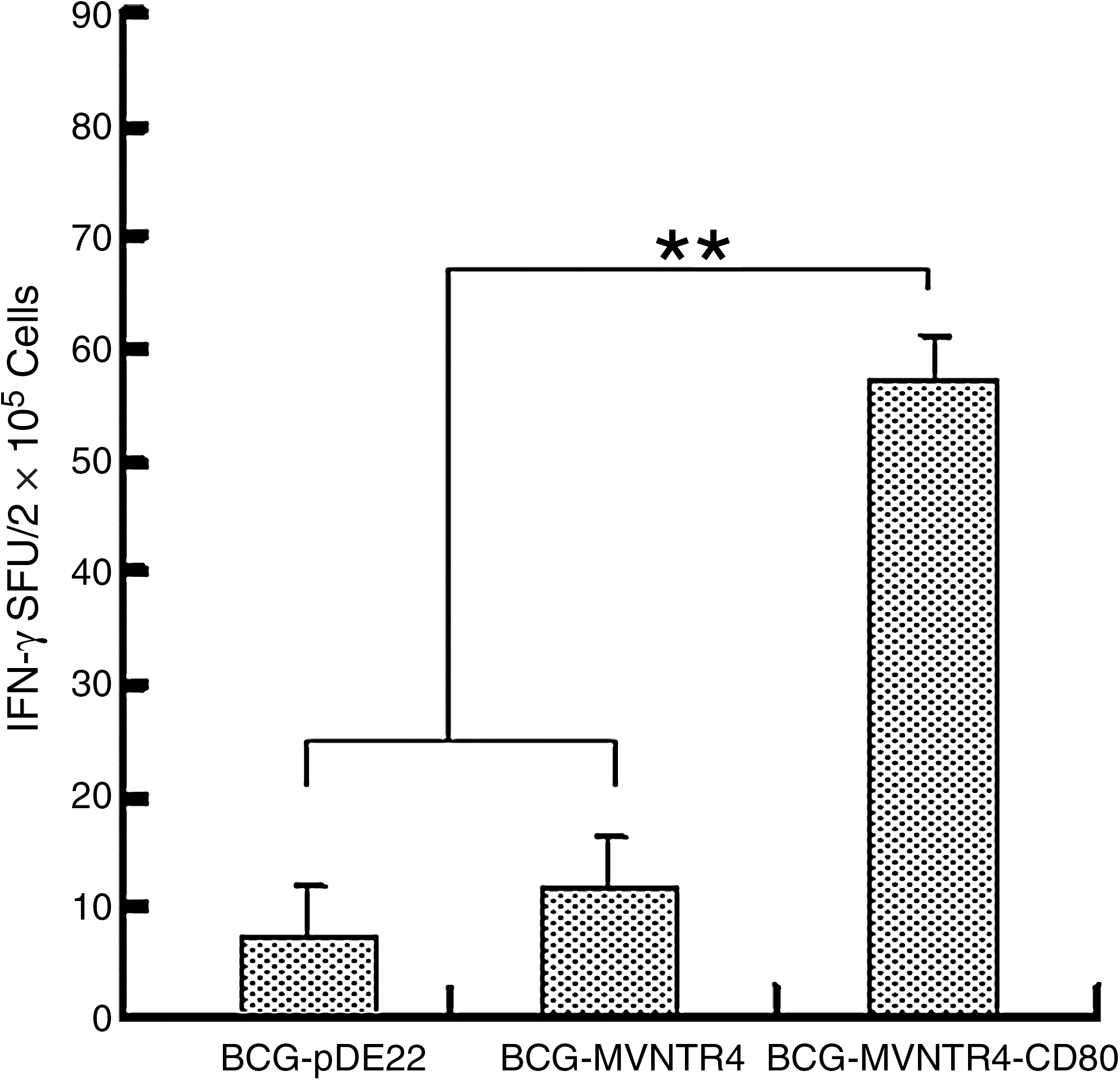
Interferon-gamma (IFN-γ) ELISpot assay of splenocytes from immunized mice. Secretion of IFN-γ by splenocytes in response to MUC1 (n = 3 mice/group) was detected. For each animal, the mean number of spots of negative control wells was subtracted from the number of spots in test wells to determine the mean number of MUC1-specific IFN-γ spot-forming units. A significant IFN-γ increase in the splenocytes from rBCG-MVNTR4-CD80-immunized mice was found, compared with those from rBCG-MVNTR4 or rBCG-pDE22-immunized mice (**p < 0.01).
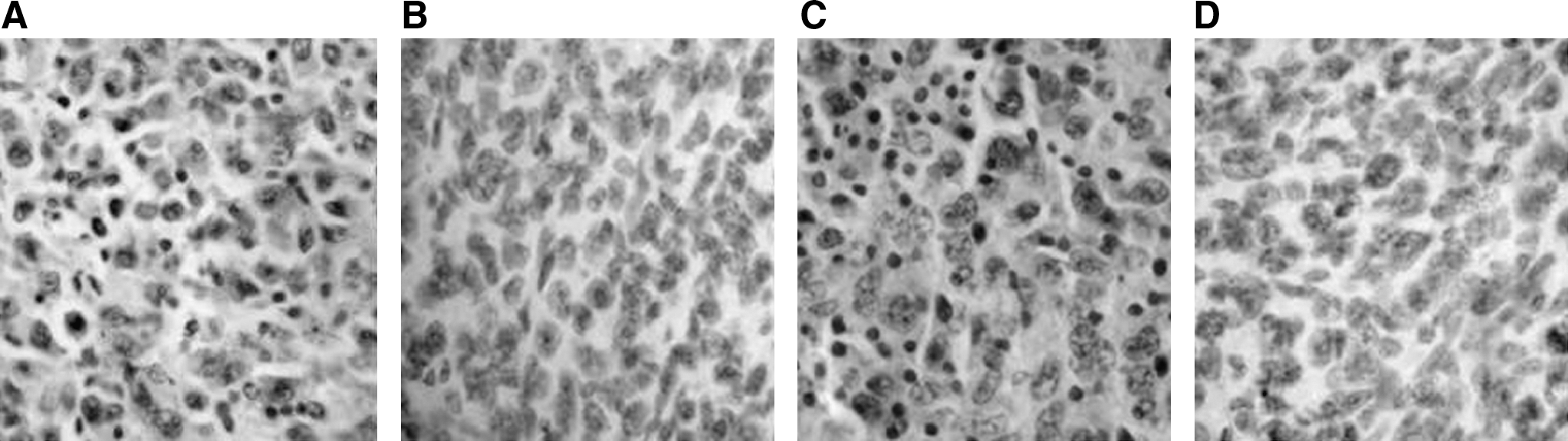
Immunohistochemistry of tumors samples for CD4/CD8-positive lymphocytes. Tumors samples obtained from mice immunized with rBCG-MVNTR4-CD80 were stained for the presence of CD4-positive lymphocytes (
Retention of pDE-MVNTR4 and pDE-MVNTR4-CD80 by BCG in vivo
In order to detect whether BCG, MUC1, and CD80 proteins still existed at 70 days after tumor inoculation, the spleens obtained from mice immunized with rBCG-MVNTR4-CD80 were detected by immunostaining. As can be seen in Figure 6, the spleens from rBCG-pDE22 or rBCG-MVNTR4-CD80-immunized animals were strongly positive for mycobacteria (Fig. 6A and 6B). In addition, the expression of MUC1 and CD80 proteins in the spleens from rBCG-MVNTR4-CD80-immunized animals could be detected (Fig. 6C and 6E). There were no detectable expressed MUC1 and CD80 protein in spleens obtained from mice immunized with BCG-pDE22 (Fig. 6D and 6F). This result suggested that BCG could retain pDE-MVNTR4-CD80 for at least 70 days in vivo.
BCG retention of pDE-MVNTR4-CD80 in vivo. Spleens from mice immunized with rBCG-pDE22 and rBCG-MVNTR4-CD80 were stained for the presence of mycobacteria, MUC1, and CD80 expression. Mycobacteria were detected in the spleens from rBCG-pDE22-vaccinated animals (
Discussion
Although MUC1 is a breast cancer-associated antigen, cellular immune responses to MUC1 are impaired in breast cancer patients and MUC1 transgenic animals. 18,19 One main reason for this weak immunogenicity of MUC1, we suppose, was the lack of costimulatory signal of B7 molecules. Therefore, the vaccine, BCG-MVNTR4-CD80, was designed with the following features: 1) multiple tandem repeats (VNTR4) of MUC1 were used as an antigen; 2) the costimulatory molecule CD80 and VNTR4 were coexpressed; and 3) BCG was used as a vaccine vehicle vector.
The ability of rBCG-VNTR4-CD80 to inhibit the growth of xenografted human breast cancer cells in hu-PBL-SCID mice model was observed. The results showed that the inhibition effect of tumor growth on rBCG-VNTR4-CD80-immunized mice was significantly stronger than that on rBCG-MVNTR4, rBCGpED22, and PBS-immunized mice. However, the tumor growth in the animals immunized with rBCG-MVNTR4 did not show obvious inhibition, suggesting that the rBCG-MVNTR4 immunization alone was insufficient to induce a cellular immune response against MUC1-positive breast tumor challenge in hu-PBL-SCID mice. Importantly, the survival of the animals immunized by rBCG- MVNTR4-CD80 was markedly increased. It indicated that animals vaccinated with rBCG-MVNTR4-CD80 were protected against breast cancer development.
Previous studies showed that cellular immune responses are required for tumor protection. 20 –22 To identify whether the inhibition effect of tumor growth in the animals immunized with rBCG-MVNTR4-CD80 was due to a MUC1-specific T-cell response, IFN-γ ELISpot assays were performed. The result showed that rBCGMVNTR4-CD80 immunization induced a significant level of IFN-γ secreting T-cells, whereas the rBCG-MVNTR4 immunization alone did not, indicating that rBCGMVNTR4-CD80 immunization could break the immune tolerance to MUC1 in mice. Because the mice immunized with rBCG-MVNTR4 did not show a significant antitumor effect and level of IFN-γ secreting T-cells, we suggested that CD80 played a crucial role in breaking the immune tolerance to MUC1 in hu-PBL-SCID mice. It has been proved that CD8+ T-cells play a dominant role in tumor protection mediated by the vaccine. 17 In this study, CD8 and CD4-positive human lymphocytes were detected only in tumors grown in rBCG-MVNTR4-CD80-immunized animals. Taken together, we suggest that rBCG-MVNTR4-CD80 may stimulate a cell-mediated immune response, resulting in the inhibition growth of MUC1-positive breast tumor cells.
BCG is currently the most widely used human vaccine in the world because it can potentially offers unique advantages for developing a safe, effective multivaccine vehicle. 23 –25 Basically, BCG is safe as a vaccine vehicle. We observed that the retention time of pDE-MVNTR4-CD80 in the spleens from mice immunized with rBCG-MVNTR4-CD80 was at least 70 days. Meanwhile, the expression of MUC1 and CD80 proteins in the spleens from rBCG-MVNTR4-CD80-immunized animals could also be detected. More important, the survival of the mice immunized by rBCG-MVNTR4-CD80 vaccine was markedly increased in this period, indicating that our rBCG vaccine was safe and stable in vivo.
Conclusions
In conclusion, we have engineered an rBCG-based vaccine that coexpressed four MVNTRs of MUC1 and CD80 and observed the therapeutic effect of the vaccine on MUC1-positive breast tumor. The results showed that the growth of MUC1-positive breast tumor in the mice immunized with the vaccine was obviously inhibited, and the survival of the mice was significantly increased.
Footnotes
Acknowledgments
We thank Professor Li Yuan (Department of Microbiology, Fourth Military Medical University, Xi'an, China) for kindly providing the E. coli-BCG shuttle vector pDE22. This work was supported by a grant from the National Natural Science Foundation of China (No. 30571802).
Disclosure Statement
No competing financial interests exist.