
Abstract
Aim:
The aim of this study was to explore the inhibitory effects of Tanshinone IIA on the production of proinflammation cytokines in radiation-stimulated microglia.
Methods:
Microglia cells were treated with 2, 4, 8, 16, and 32 Gy of irradiation or sham-irradiated in the presence or absence of 1.0 μg/mL of Tanshinone IIA. The effects of Tanshinone IIA on radiation-induced proinflammatory cytokines were evaluated by real-time polymerase chain reaction; the expression level of nuclear factor (NF-κβ) p65 in cytoplasm and nucleus was measured by Western blot. Immunofluorescence staining and confocal microscopy analysis were applied to detect the expression of γ-H2AX and p65 postirradiation.
Results:
Radiation-induced release of proinflammatory cytokines in BV-2 cells was detectable after irradiation. Tanshinone IIA decreased the radiation-induced release of proinflammatory cytokines. Further, Western blotting showed that Tanshinone IIA could attenuate the nuclear translocation of (NF-κβ) p65 submit postirradiation. Immunofluorescence staining showed γ-H2AX foci formation with p65 translocation into the nucleus postirradiation.
Conclusions:
Our data indicated that Tanshinone IIA exerts anti-inflammatory properties by suppressing the transcription of proinflammatory cytokine genes that might be associated with the NF-κβ signaling pathway. It is postulated that irradiation causes immediate cellular reaction, and that double-strand breaks trigger the molecular response that leads to NF-κβ pathway activation.
Introduction
Although associated with the risk of adverse side-effects, cranial radiotherapy (CRT) is an invaluable therapeutic tool of primary and secondary malignant brain tumors in both pediatric and adult patients. About 50% of long-term survivors develop late-delayed effects caused by radiation-induced injury of normal brain tissue, 1 particularly in children. 2,3 Radiation-induced brain injury (RIBI) is a complex, dynamic progress. Although the exact pathogenic mechanisms of RIBI are still undefined, recent data suggest that neurogenesis inhibited by proinflammatory responses might be a role. 4 –6 Thus, inhibition of these proinflammatory mediators and cytokines with new drugs may be a new clew to the treatment of RIBI.
Microglia cells are a kind of brain mononuclear phagocytes, accounting for up to 10% of the overall number of cells for most of development. In the rest state, microglia cells are ramified and characterized by a small cell body with multiple processes, which is the basis of normal neurogenisis. 7 Activated microglia cells develop a more rounded appearance, and their processes retract into the soma. Microglia cells could be activated by a variety of stimuli and release many proinflammatory cytokines and chemokines. 8 There are studies indicating that brain irradiation leads to a marked increase in microglia activation associated with both a concomitant decrease in neurogenesis of the hippocampus and spatial memory retention deficits in vivo. 9,10 These data suggest that radiation-induced microglia activation may play a role in RIBI.
A previous study showed that microglia could be activated by many stimuli, such as: lipopolysaccharide (LPS), 6 ischemia, trauma, and so on. Recent research suggested that irradiation could induce microglia activation in vitro, which leads to a marked increase in the expression of proinflammatory cytokines. 11 Tanshinone IIA is a derivative of phenanthrene-quinone isolated from Salvia miltiorrhiza Bunge. It has antioxidant, inflammation, 12 and tumor properties. 13 It has been shown that Tanshinone IIA significantly inhibits LPS-induced Iκβ alpha degradation and nuclear factor (NF-κβ) activation in RAW 264.7 cells. 12 DNA double-strand breaks (DSBs) are the most deleterious form of DNA damage, and numerous in vitro studies have emphasized the tremendous effects of an efficient DSB repair system on cell survival after exposure to ionizing radiation. DSB could rapidly trigger the activation of the NF-κβ pathway. The aim of this study was to investigate the effects of Tanshinone IIA on the release of proinflammatory cytokines interleukin-1-beta (IL-1β), tumor necrosis factor alpha (TNF-α), IL-6, interferon-gamma (IFN-γ), and cyclooxygenase-2 (COX-2) in irradiated microglia and the relationship between DSB and microglia's NF-κβ pathway activation.
Materials and Methods
Cell culture and reagents
The immortalized BV-2 murine cells were obtained from the Central Laboratory of Wuhan Union Hospital (Wuhan, China). They were maintained in RPMI-1640 (Gibco, BRL, Grand Island, NY), supplemented with 10% heat-inactivated fetal bovine serum (FBS), 100 U/mL of penicillin, and 100 g/mL of streptomycin at 37°C, in a humidified atmosphere of 5% CO2. Tanshinone IIA was purchased from the Chinese Institute for Drug and Biological Product Control (Beijing, China). 3-(4,5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2H-tetrazolium bromide (MTT) was obtained from Sigma (St. Louis, MO). The anti-γ-H2AX mouse monoclonal antibody (mAb) was bought from Abcam, Inc. (Cambridge, MA), the AlexaFluor-488 conjugated goat-antimouse secondary antibody and AlexaFluor-568 conjugated goat-antirabbit secondary antibody were purchased from Invitrogen (Carlsbad, CA). VCTASHIELD mounting medium with 4’,6-diamidino-2-phenylindole was taken from Vector Laboratories (Burlingame, CA). The ECL Western blotting detection system was purchased from Millipore (Bedford, MA). P65 rabbit polyclonal antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA), and the antibody against β-actin was purchased from Sigma. Trizol was obtained from Invitrogen. M-MLV reverse transcriptase and SYBR Green I were purchased from Toyobo Company (Osaka, Japan). The Oligo dT and primers were synthesized by Shanghai Invitrogen (Shanghai, China). The dNTP and Taq DNA polymerase were purchased from Fermentas International, Inc. (Burlington, Canada).
Cytotoxicity assay
The cytotoxicity of Tanshinone IIA was determined by MTT assay. Cells (1 × 105/well) were planted in 100 μL of medium/well in 96-well plates. After incubation overnight, the medium in each well was discarded and replaced with a fresh medium with various concentrations of Tanshinone IIA (0.25–2.0 μg/mL). BV-2 cells without treatment with Tanshinone IIA were used for control. After treatment with Tanshinone IIA for 24, 48, 72, and 96 hours, 20 μL of 5 mg/mL of MTT (pH 4.7) were added to each well and cultivated for another 4 hours, the supernatant fluid was removed, 100 μL/well of dimethyl sulfoxide (DMSO) were added and samples were shaken for 15 minutes. The optical density (OD) at 570 nm was measured with a microplate reader (Bio-Rad, Richmond, CA), using wells without cells as blanks. All experiments were performed in triplicate.
Irradiation
Cells were irradiated by using a 137Csir radiator (Siemens, Munich, Germany) at a dose rate of 2.0 Gy/min. All irradiations were performed at room temperature, whereas control cells received sham irradiation. After irradiation, the culture dishes were returned to a standard condition (37°C, 5% CO2).
Cell morphous observation
After BV-2 cells were irradiated for 3, 6, 12, 24, and 48 hour in the presence or absence of 1.0 μg/mL of TanshinoneIIA, the cell morphology was observed under inversion microscopy and were compared with the changes of the control group.
Colocalization of γ-H2AX and p65 immunofluorescence staining and confocal microscopy
BV-2 cells were plated onto polylysine-coated cover glasses and treated with irradiation. Then, samples were fixed in methanol, permeabelized in acetone, and then blocked for 30 minutes in 1% goat serum. Anti-γ-H2AX primary antibody (1:800) and anti-p65 primary antibody (1:200) were applied after blocking overnight in 4°C, followed by three 5-minute washes in phosphate-buffered saline (PBS), then AlexaFlour-488-conjugated goat-antimouse secondary antibody (1:400) and AlexaFluor-568 conjugated secondary antibodies (1:400) was applied for 1 hour at room temperature (RT) in the dark, also followed by three 5-minute washes in PBS. After, samples were mounted in VECTASHIELD mounting medium with 4,6-diamidino-2-phenylindole. Cells were observed by using fluorescence microscope (Olympus, Tokyo, Japan) for green, red, and blue fluorescence, then observed by using confocal laser microscopy (FV500; Olympus) at X600 magnification.
RNA isolation and real-time QPCR
Total RNA was purified from cultured cells by using Trizol reagent, according to the manufacturer's protocol. Primer sequences designed from using Beacon Designer software (Bio-Rad) were as follows: mouse TNF-α 5′ primer TTC TCA TTC CTG CTT GTG G and 3′ primer CTT GGT GGT TTG CTA CGA C; mouse IL-1β5′ primer AAA TCT CGC AGC AGC ACA T and 3′ primer CAC ACA CCA GCA GGT TAT CA; mouse IFN-γ 5′ primer TAA CTC AAG TGG CAT AGA TGT G and 3′ primer GAA GAA GGT AGT AAT CAG GTG TG; mouse IL-6 5′ primer TTG CCT TCT TGG GAC TGA T and 3′ primer TTG CCA TTG CAC AAC TCT T; mouse COX-2 5′ primer GAG TGG GGT GAT GAG CAA and 3′ primer GCA ATG CGG TTC TGA TAC T; GAPDH 5′ primer TCA CCA CCA TGG AGA AGG C and 3′ primer GCT AAG CAG TTG GTG GTG CA. Real-time reverse transcriptase polymerase chain reaction (RT-PCR) was performed by Stratagene MX3000P QPCR System (Stratagene, La Jolla, CA), according to the manufacturer's recommendations. Each 20-μL real-time PCR system contained 6.4 μL 3dH2O, 0.8 μL (20 pmol/μL) of each primer, 10 μL SYBR Green Realtime PCR Master Mix, and 1 μL of the original cDNA synthesis reaction. Fifty (50) cycles of amplification were performed: after 94°C, 30 seconds, to 57°C, 30 seconds, then to 72°C, 30 seconds was carried out 50 times. The fluorescence signal was detected at the end of each cycle. Melting curve analysis was used to confirm the specificity of the products. The 2−ΔΔCt method was performed to analyze the results. The relative expression levels of each mRNA were calculated by using the ΔΔCt method normalizing to GAPDH and relative to the control samples. Briefly, the Ct of the housekeeping gene was subtracted from the Ct of IL-1β, TNF-α, IL-6, IFN-γ, and COX-2 to get ΔCt. The ΔCt value of the sham-irradiated sample was then subtracted from the ΔCt of the rest of the treatments to get the ΔΔCt value. Fold differences, compared to the sham-irradiated sample, are obtained by calculating 2−ΔΔCt for each treatment group. Data represent the mean ± standard error of the mean of three independent experiments.
Western blot analysis
Cells were seeded in six-well plates and pretreated with or without 1.0 μg/mL of Tanshinone IIA before 32-Gy irradiation for 3, 6, 12, 24, and 48 hours. The cells were then lysed for immunoblotting. To determine the levels of NF-κβ, cytoplasmic or nuclear extracts were prepared from irradiated cells 14 and then resolved on 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE).
Proteins were transferred to nitrocellutose membranes for 12–16 hours at 30 V and blocked in 5% nonfat skim milk in 1% Tris-buffered saline with 0.1% Tween-20 (TBST) for 1 hour at RT and then incubated overnight with primary antibody (1:200) diluted in 2% BSA in TBST at 4°C. The following day, proteins were visualized by using the ECL detection system after incubation with the respective horseradish peroxidase (HRP)-conjugated secondary antibody (1:1000). Complexes of the primary and secondary antibodies were visualized by using a chemiluminescnet substrate kit and exposed to medical X-ray film. The intensity of the blots was quantified with a gel-image analyzer (JS380; Peiqing Science and Technology, Shanghai, China).
Statistical analysis
Data are presented as the mean ± standard error of at least three separate experiments. The Student's t-test was used to determine statistical significance between the control and test groups. A p-value less than 0.05 was considered to be statistically significant.
Results
Irradiation-activated microglia cells demonstrated by morphologic observation
In the rest situation, microglia showed a dendritic morphology with many highly ramified processes (Fig. 1A). There was not morphologic change in BV-2 after 2, 4, and 8-Gy irradiation. However when treated with 16 and 32 Gy, the BV-2 cells changed to round or amoeboid appearance, which indicated the microglia was activated by irradiation (Fig. 1B). A majority of the irradiated microglia cells pretreated with Tanshinone IIA showed ramified cytoplasmic processes of various degrees (Fig. 1C).
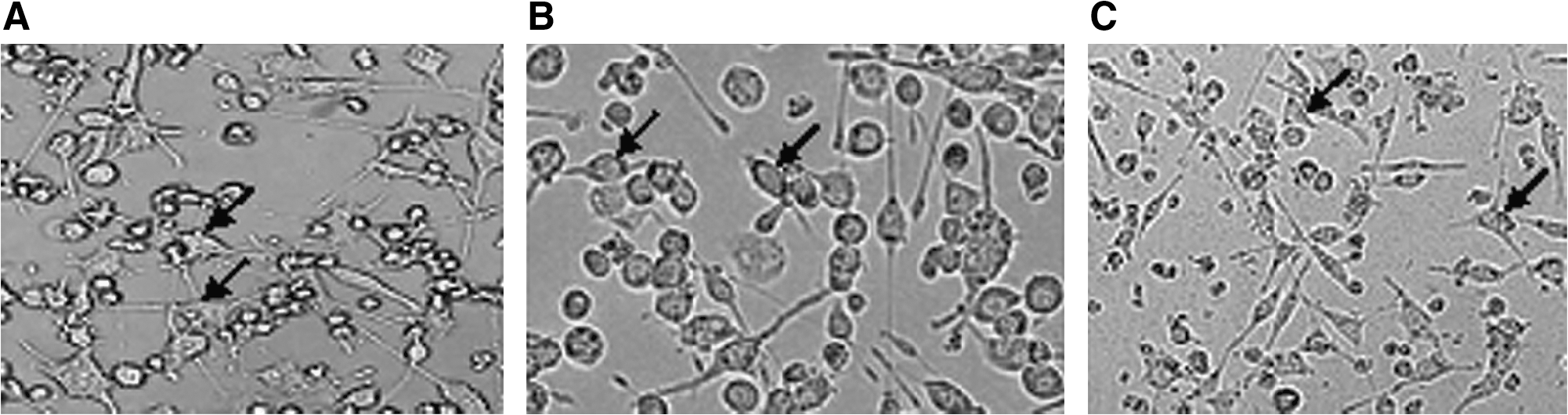
The morphologic change of the BV-2 cells postirradiation. (
Tanshinone IIA decreased the production of proinflammation cytokines in radiation-induced microglia inflammatory response
To investigate the effect of Tanshinone IIA on cell viability, the cells were exposed to Tanshinone IIA for 24 hours. As shown in Figure 2, the concentrations (0.25–2.0 μg/mL) of Tanshinone IIA used here had no effect on the viability of BV-2 cells. We used the dose below 2.0 μg/mL to avoid the Tanshinone IIA cytotoxicity effect on the BV-2 cell line.
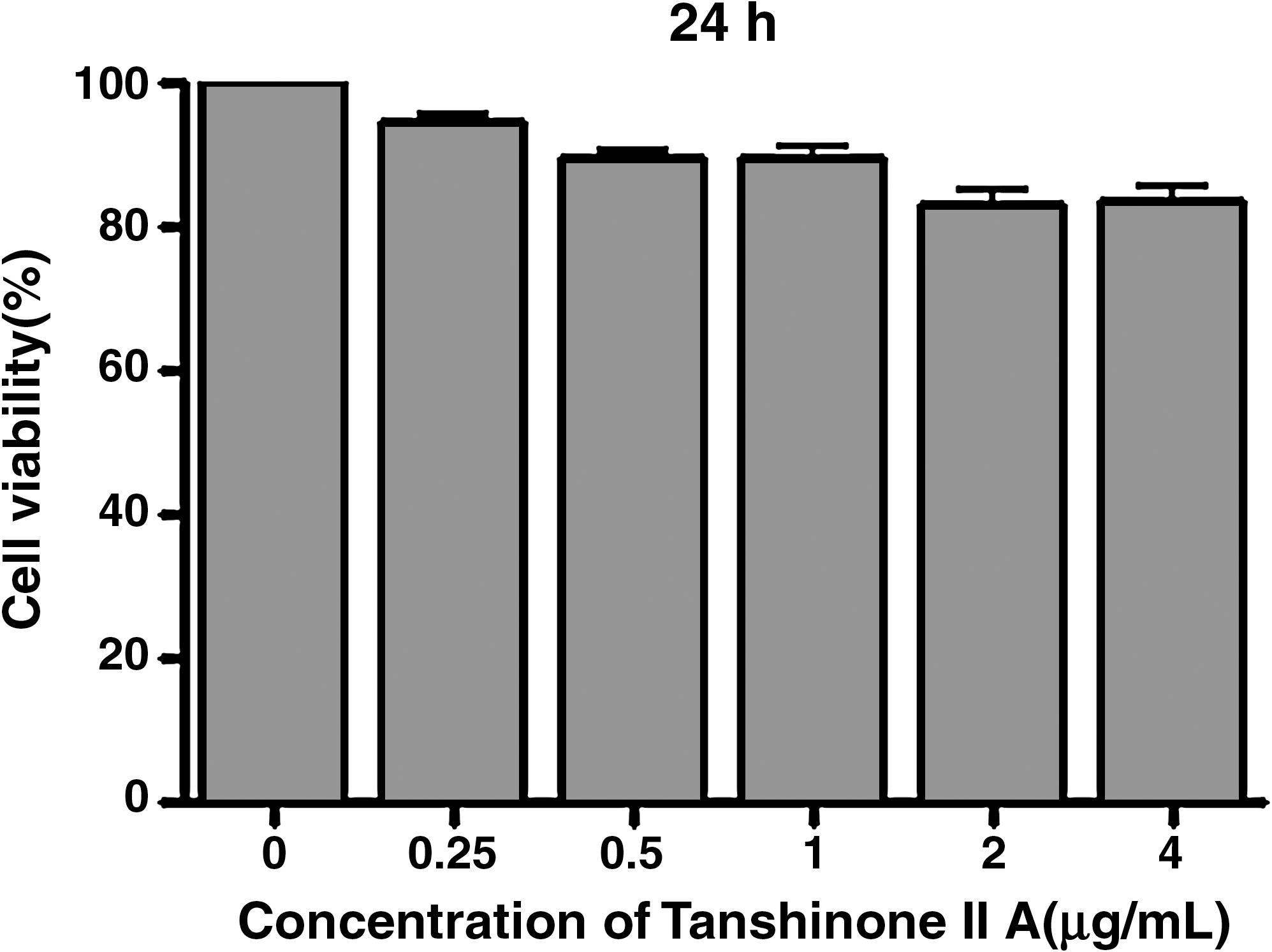
Effect of Tanshinone IIA on the cell viability of BV2 microglia. The BV-2 cells were seeded into culture plates and treated with 0.25, 0.5, 1.0, 2.0, and 4.0 μg/mL Tanshinone IIA for 24 hours, then cell viability was determined by MTT assay. Data are expressed as mean ± standard deviation of three measurements.
As shown from Figure 3 on proinflammatory cytokines, the levels from being stimulated by irradiation were significantly higher than those of the control group (p < 0.05). There was a similar change with 8-Gy postirradiation. But after 2- and 4-Gy irradiation, no increase of proinflammatory cytokine expression was found. We next investigated Tanshinone IIA's effect on the expression of proinflammatory cytokines, such as IL-1β, TNF-α, IL-6, IFN-γ, and COX-2. BV-2 microglia cells were incubated with Tanshinone IIA (1.0 μg/mL) in the presence or absence of irradiation (32 Gy). Pretreatment with Tanshinone IIA for 2 hours resulted in a decrease in cytokine production at 24 hours postirradiation (Fig. 3). IL-1β, TNF-α, IL-6, IFN-γ, and COX-2 mRNA were not detectable under normal culture conditions; however, real-time PCR assays revealed that pretreatment with Tanshinone IIA for 2 hours attenuated the upregulation of these cytokines (p < 0.05). These results suggested that Tanshinone IIA acted principally by preventing the accumulation of proinflammatory cytokines via inhibiting the transcription of these genes.
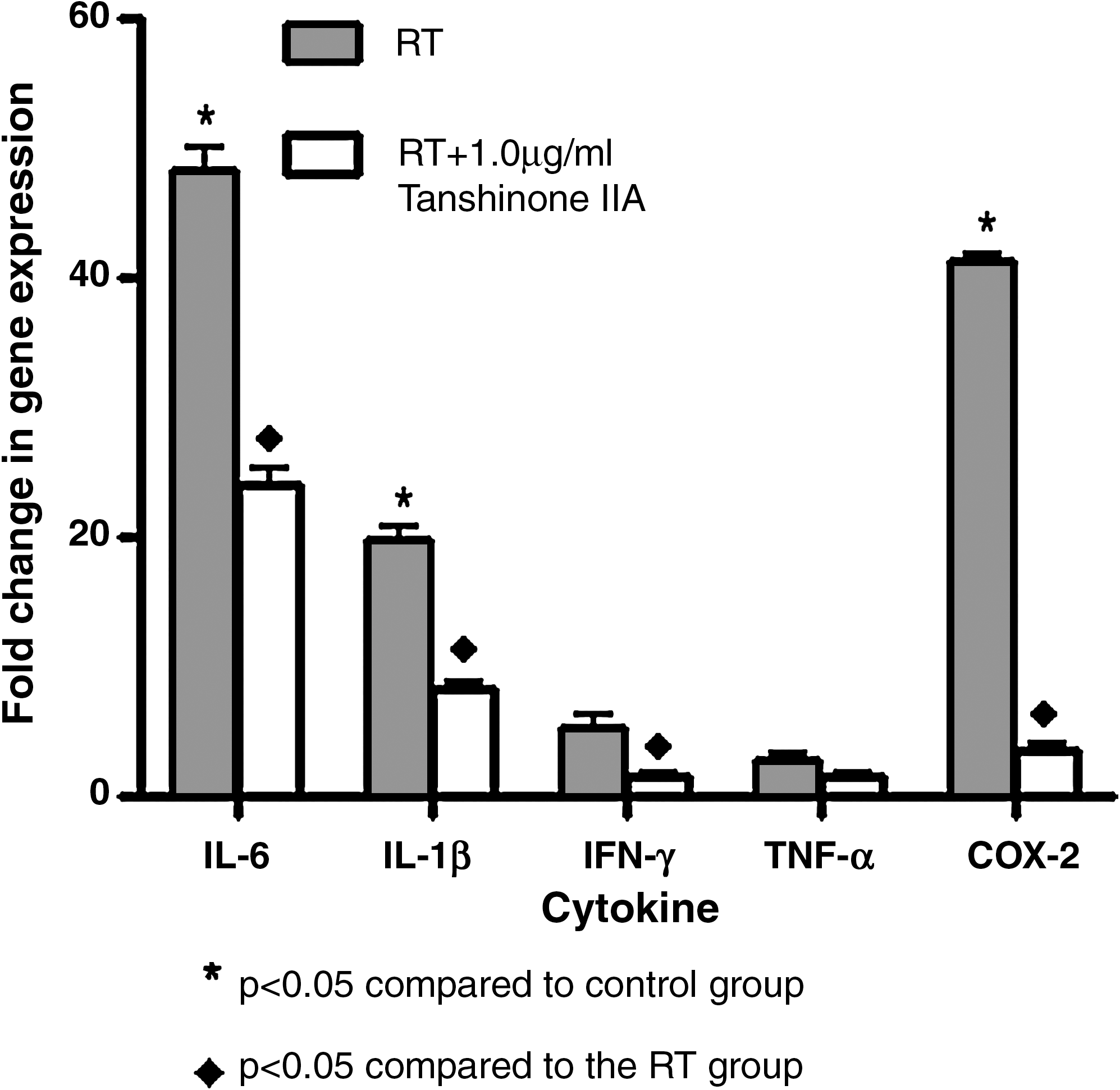
Effect of Tanshinone IIA on proinflammatory cytokines in irradiation-stimulated microglia. Cells were pretreated with 1.0 μg/mL of Tanshinone IIA for 2 hours before 16-Gy irradiation and the expression of inflammatory cytokines was analyzed at 24 hours.
Inhibitory effects of Tanshinone IIA were mediated by NF-κβ pathway suppression in radiation-induced BV-2 cell inflammatory response
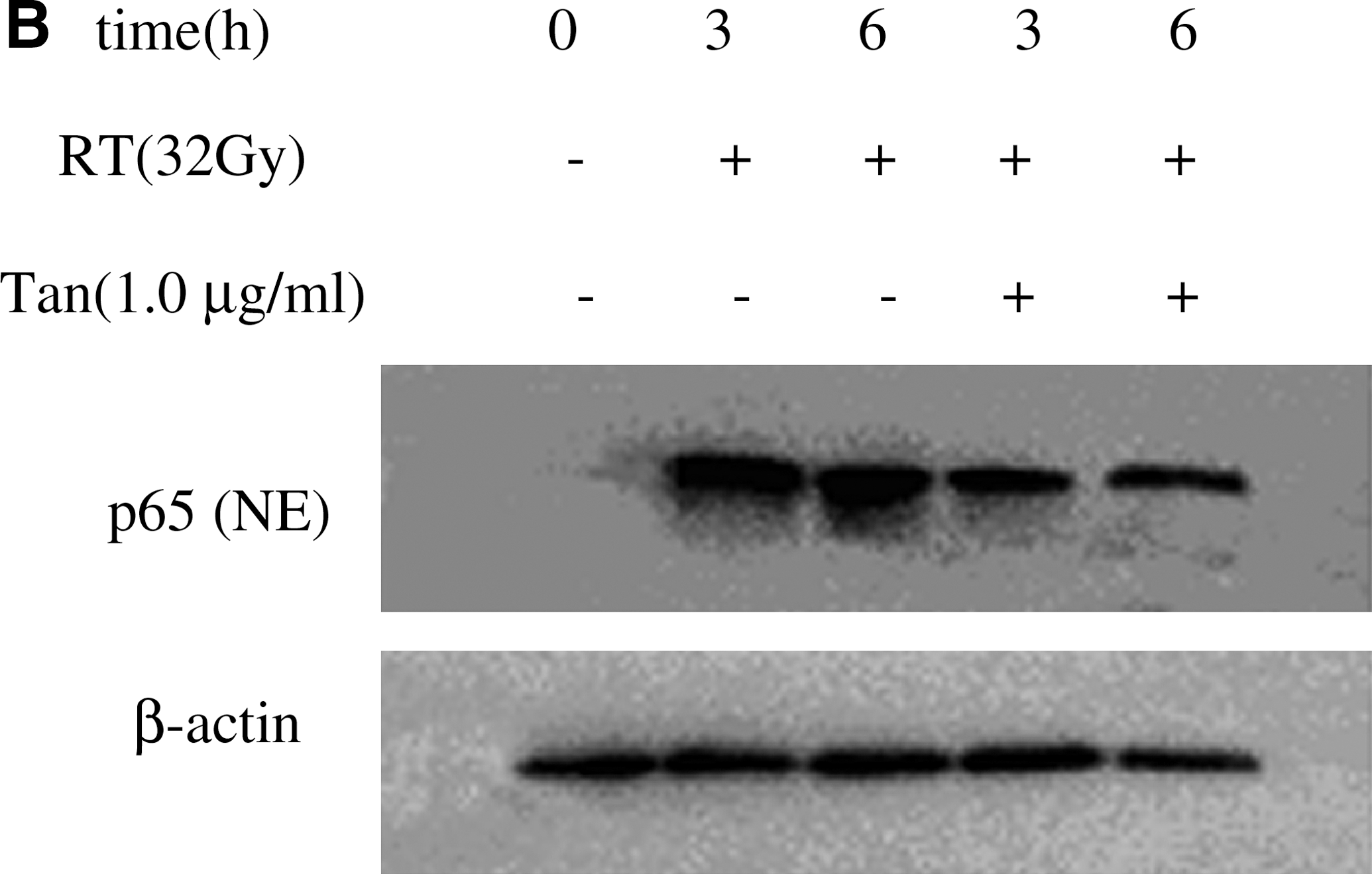
We investigated the translocation of NF-κβ from cytoplasm to nucleus. As shown in Figure 4A, it was observed a significant increase in the p65 subunit of NF-κβ post-8-Gy and 16-Gy irradiation from the nuclear extract in BV-2 cells. No p65 protein expression was found at 2 and 4 Gy postirradiation. This finding suggested that the p65 subunit was involved in the radiation response. Next, we tested whether Tanshinone IIA could prevent radiation-induced NF-κβ pathway activation in microglia cells. As shown in Figure 4B and 4C, treatment with 32-Gy irradiation alone increased the translocation of p65 from cytoplasm to the nucleus at 3 and 6 hours, but Tanshinone IIA inhibited the translocation of p65. Tanshinone IIA significantly inhibited radiation-induced NF-κβ pathway activation.
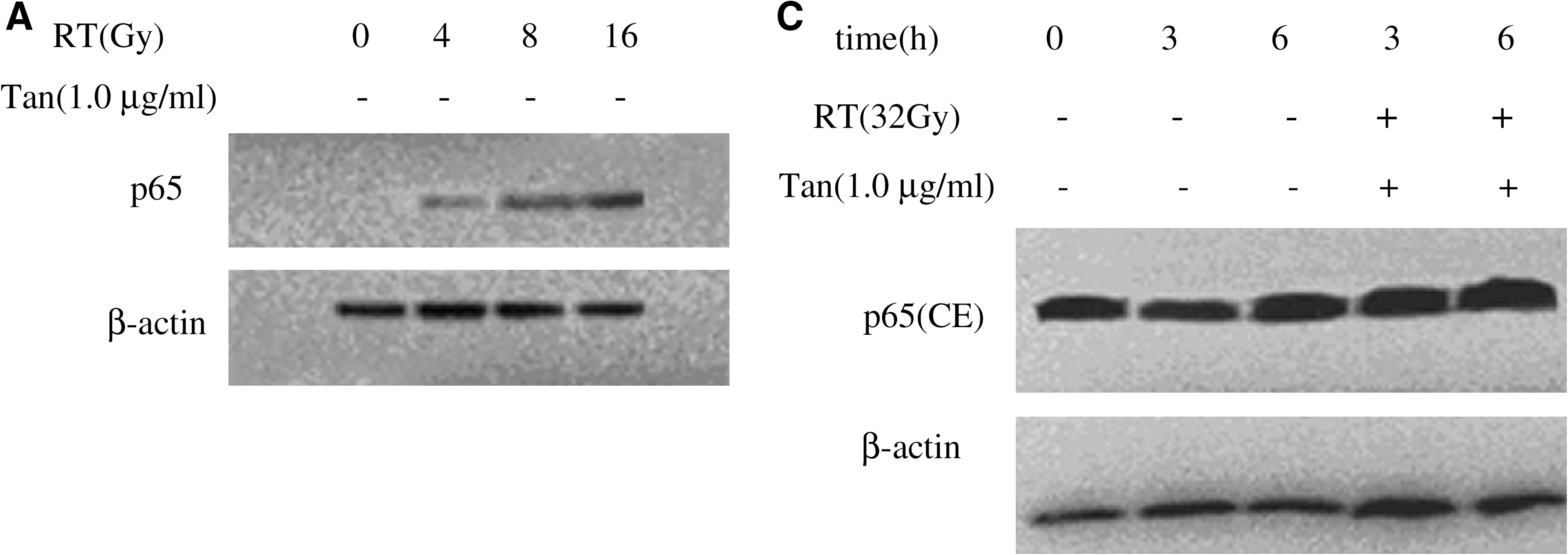
The expression of p65 was analyzed by Western blot. (
Colocalization of γ-H2AX and p65
To investigate the repair of DSBs and the translocation of p65 in BV-2 cells after irradiation, we established the double immunofluorescence staining for γ-H2AX and p65 (Fig. 5). Whereas the sham-irradiation BV-2 cells were almost completely negative for γ-H2AX (Fig. 5A), distinct nuclear γ-H2AX foci were observed in BV-2 cells 3 hours after irradiation (Fig. 5B). The number of γ-H2AX foci decrease with increasing postirradiation repair time. This fast component of foci loss was followed by a slower process at later times, and only very low levels of γ-H2AX foci were observed at 24 hours. The expression of p65 was located in cytoplasm in the control group (Fig. 5A). After 32-Gy irradiation for 3 hours, cell nucleus stained to red presented p65 translocation (Fig. 5B), but when pretreatment with 1.0 μg/mL of Tanshinone IIA for 2 hours, p65 protein translocation was inhibited and the cytoplasm was stained in a red color (Fig. 5C).
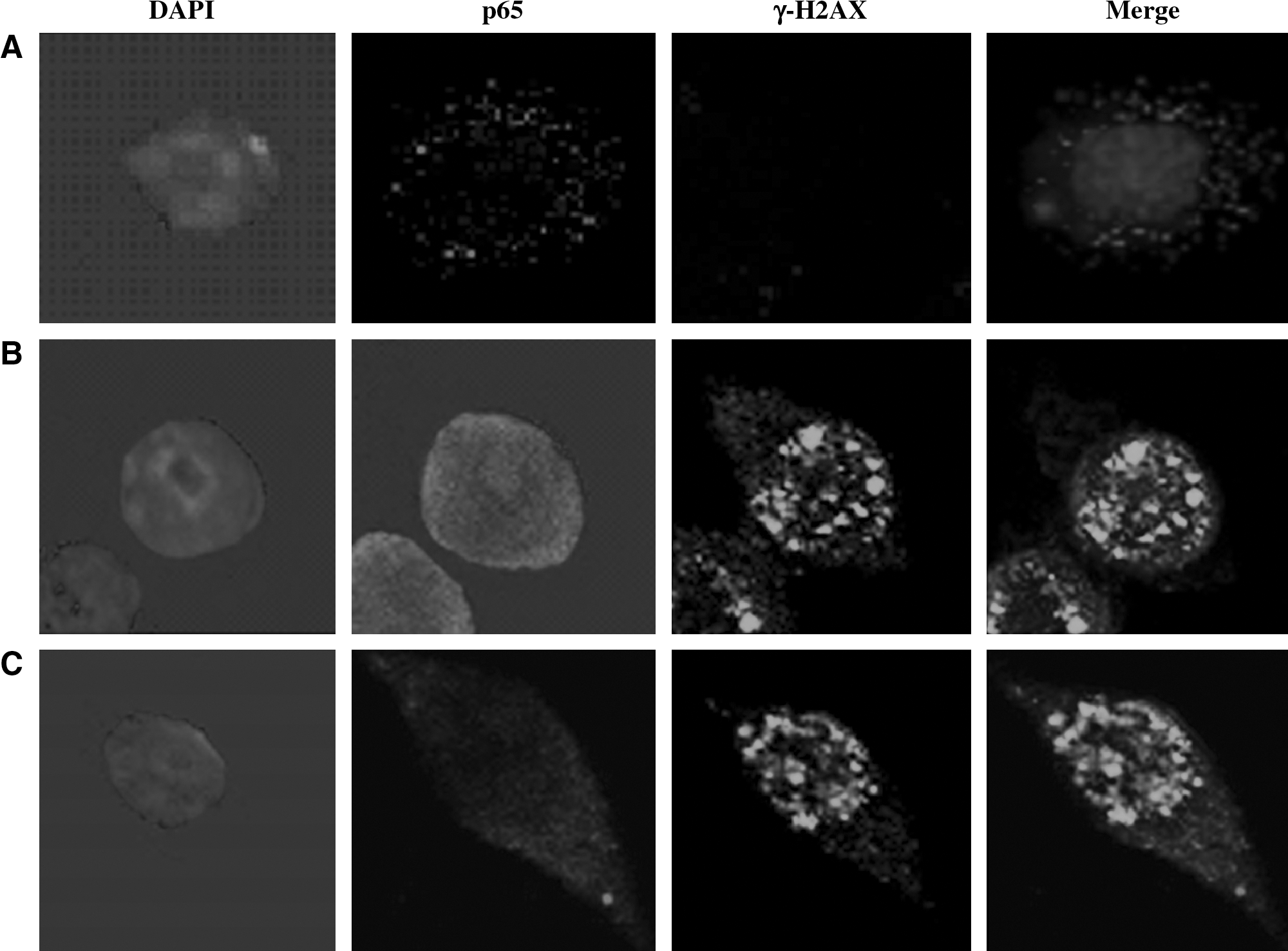
The expression of γ-H2AX and p65 postirradiation. Confocal images of immunofluorescence double staining for γ-H2AX and p65 in BV-2 cells after 32-Gy irradiation at 3 hours with or without pretreatment with 1.0 μg/mL of Tanshinone IIA. DNA counterstained with 4′,6-diamidino-2-phenylindole. Original magnification X600. (
Discussion
It was documented that Tanshinone IIA has anti-inflammatory properties. 12 However, the anti-inflammatory effect on BV-2 cells had not been reported and related exploration on molecular biologic mechanisms of the anti-inflammatory role of Tanshinone IIA remained limited.
In the present study, we showed that the inhibitory mechanisms of Tanshinone IIA on the production of proinflammatory mediators in BV-2 cells was stimulated with irradiation. DSBs are the most deleterious form of DNA damage after ionizing radiation, and deficiencies in repairing DSBs lead to pronounced radiosensitivity. H2AX phosphorylation is an early step in the response to DNA damage, and it has been demonstrated that enumerating γ-H2AX foci (the phosphorylated histone) can be used to measure the induction and repair of radiation-induced DSBs in vitro and in vivo. 15,16 A crucial component of the DNA DSB signaling cascade in mammalian cells is the protein kinase, ATM. 17 It has been suggested that ATM could activate NF-κβ in response to DNA damage. 18,19 NF-κβ plays a critical role in the expression of proinflammatory cytokines. 18 Irradiation has been shown to initiate multiple intracellular signaling events. It has been suggested that NF-κβ could be activated in response to DNA damage. 17,20,21 The NF-κβ signaling pathway mediates a variety of important cellular functions by regulating immune and inflammatory responses. In unstimulated cells, NF-κβ is in the form of a heterodimer of p65/p50 binding to the inhibitor proteins Iκβ. After stimulation, the release of p65/p50 from the Iκβ-α protein and the degradation of Iκβ-α are necessary for p65 translocation into the nucleus to regulate gene transcription. Central to the activation of NF-κβ by stimuli is the Iκβ kinase (IKK) complex that is composed of IKKα/1 and IKKβ/2 and NF-κβ essential modulator (NEMO/IKKγ). In the present study, it was found that Tanshinone IIA could inhibit p65 translocation postirradiation in BV-2 microglia cells, which indicated that NF-κβ pathways might be involved in the suppressive effects of Tanshinone IIA on the release of proinflammation cytokines in BV-2 microglia cells in the radiation-induced microglia activation.
COX-2 is the key enzyme regulating the production of prostaglandins and the central mediators of inflammation. The expression of COX-2 is included by several extracellular signals, including proinflammatory stimuli.
Several proinflammatory gene products have identified that mediate a critical role in inflammation, such as COX-2, IL-6, IFN-γ, IL-1β, and so forth. The expression of all these genes is mainly regulated by the transcription factor NF-κβ. Therefore, suppressing those proinflammatory cytokines and mediators is a key to evaluate anti-inflammatory drugs.
Our studies indicated that irradiating BV-2 cells leads to an increase in proinflammatory responses of the microglia, such as TNF-α, IL-1β, IFN-γ, COX-2, and IL-6, which are consistent with the previous study. 22 Further, the data also confirmed that irradiating microglia can activate the translocation of p65. Tanshinone IIA exerts anti-inflammatory properties by suppressing the transcription of proinflammatory cytokine genes through the NF-κβ signaling pathway.
Microglia cells are resident immune cells within the central nervous system and play a critical role in the development of an inflammatory response within the brain. Microglia cells are activated in response to ionizing radiation, resulting in the induction of proinflammatory cytokines. 23,24 We hypothesized that pretreatment of the microglia cells with Tanshinone IIA would prevent radiation-induced brain injury through inhibiting proinflammatory responses.
Conclusions
In summary, our study indicated that Tanshinone IIA exerts anti-inflammatory properties by suppressing the transcription of proinflammatory cytokine genes through the NF-κβ signaling pathway. We postulated that irradiation causes immediate cellular reaction and DSBs trigger the molecular response that leads to NF-κβ pathway activation.
Footnotes
Acknowledgments
This work was supported by grants from the National Nature Science Foundation of China (30800283).
Disclosure Statement
No competing financial interests exist.