
Abstract
The ability of T-lymphocytes to recognize antigens and transduce signals to the nucleus successfully is a key component in the initiation and maintenance of an immune response. The present study addressed the expression status of the signal-transducing proteins in relation to the immune impairment in cervical cancer patients. Immune response was measured by evaluating lymphocyte subpopulations CD3+, CD4+, and CD8+, using flowcytometry, natural killer cell activity, using the single-cell cytotoxicity assay, lymphocyte function, using mitogenic response to PHA and T-cell activation following anti-CD3 stimulation, and production of IL-2. Expression of the T-cell signal transduction proteins, TCR-ζ, CD3-ɛ, zap-70, p56lck, PKC, NFκβ p50, Rel-A, Rel-B, and c-rel, was evaluated by using Western blot assay. A generalized depression of the immune response with respect to the different parameters evaluated was observed. Exogenous interleukin-2 (IL-2) could increase the response in all the controls and in 30% of the patients to different degrees varying from 10% to 90%. Low levels of the signaling molecules (TCR-ζ, CD3-ɛ, zap-70, p56lck, and PKC) and impairment in the transduction of NFκβ components (p50, Rel-A, Rel-B, and c-rel) to the nuclei were observed in these lymphocytes. Decreased CD4+/CD8+ ratio with an increase in suppressor cells, reduced lymphocyte proliferation, and production of IL-2 suggest a defective immune regulation in cervical cancer. Impairment in the translocation of NFκβ p50, Rel-A, and Rel-B to the nucleus and the reduced levels of signal-transducing proteins might be responsible for the decreased production of IL-2 and immune impairment in cervical cancer patients.
Introduction
Women with cervical cancer continue to be at significant risk for the development of recurrent disease, despite surgery, radiation, and chemotherapy. The response to treatment of cervical cancer may be influenced by the host's immune system. In addition, development of effective therapeutic strategies for patients with cervical cancer may be affected by abnormalities in the immune function of tumor-bearing hosts. 1 The mechanism of the reported immunosuppression, and its relation to the tumor, is not yet clear. This study addressed the expression of the signal transduction proteins of T-lymphocytes following stimulation with anti-CD3/PHA in the presence or absence of recombinant interleukin-2 (r-IL2).
Preliminary studies reported by us revealed gross immunologic alterations in terms of increased circulating immune complexes, 2 imbalance of lymphocyte populations, 3,4 lymphocyte functions, 5 serum-blocking effects, 5,6 and kinetics of target cell lysis. 7 We have shown that the patients could be classified into those 1) with adequate immune response and 2) depressed immune response, irrespective of their stage of disease. Further, it was also shown that the immune status correlated well with response to treatment. 8
Natural cell-mediated cytotoxicity (i.e., natural killer; NK) has been implicated as an integral part of the tumor–host interrelationships and may play an active role in vivo. Depressed NK-cell activity has been found in a number of tumor-bearing individuals and is reported to be related to advanced disease. 9 Reports also demonstrate that IL-2- and interferon (IFN)-mediated processes have an activating effect on NK-cell activity, 10,11 and this is being exploited in immunotherapy of a number of cancers.
Levey and Srivastava 1 and others 12 –19 reported signaling defects in T-cells from patients with various cancers and suggested that decreases in TCR-ζ, zap-70, or p56lck protein expression, or an impaired ability to translocate components of NFκβ from the cytoplasm to the nucleus, lead to an inability of T-cells to respond to antigenic stimulation and thereby contribute to tumor progression. Our report describes similar findings in cervical cancer. The levels of proteins associated with signaling in T-cells and translocation of members of the nuclear factor kappa-beta (NFκβ) family are examined in this article.
TCR-associated signal transduction takes place through a cascade of proteins. TCR-ζ protein is an essential component of the TCR complex that binds zap-70 and transduces signals following TCR activation. The protein tyrosine kinase, zap-70, is implicated, together with the Src kinase, p56lck, in controlling the early steps of the T-cell antigen-receptor (TCR) signaling cascade. TCR ligation by antigen/MHC (major histocompatibility complex) or by anti-CD3 monoclonal antibodies (mAbs) induces rapid tyrosine phosphorylation of the immunoreceptor tyrosine-based activated motifs (ITAMS) present in their cytoplasmic tails, an event essential for activation of lymphokine genes. T-cell activation involves other proteins, such as p56lck, TCR-ζ, PKC, and leads to translocation of NFκβ to the nucleus. This, in turn, induces the transcription of a variety of genes, including IL-2. This study addressed the role of these factors in the immune impairment seen in patients with cancer of the cervix.
Materials and Methods
Blood samples were collected prior to treatment from 72 cervical cancer patients, with an average age of 55.7, registered at the Regional Cancer Center (Thiruvananthapuram, India). The patients were all treated with radiation as per the standard protocols followed in this center and were followed up for periods ranging from 15 to 24 months. All patients were staged according to the FIGO (International Federation of Gynecology and Obstetrics) system of classification (stage I–IV). 20 Patients from all four stages of the disease were randomly included in the study, which included 17 patients belonging to stage I/IIA, 23 patients in stage IIB, 25 patients in stage III, and 7 patients belonging to stage IV. As controls, blood samples were collected from 70 age-matched healthy individuals. Ethical consent was obtained from cervical cancer patients and HIs (healthy individuals) prior to blood collection.
Cell culture and stimulation
Lymphocytes isolated from heparinized peripheral blood of cervical cancer patients by Ficoll-Hypaque (Ficoll-Paque, Sigma, St. Louis, MO) density gradient centrifugation were cultured in sterile RPMI-1640 medium (Gibco Life-Technologies, Grand Island, NY) containing 10 μM of gentamycin, 2 mM of L-glutamine (Gibco Life-Technologies), and 10% (v/v) heat-inactivated fetal calf serum (FCS) (Sigma) at 37°C in a humidified CO2 atmosphere. Cells were then incubated with anti-CD3/PHA in the presence or absence of r-IL2 (Sigma), followed by isolation from the culture by a magnet-activated cell sorter (MACS), using anti-CD3-coated magnetic beads (Miltenyi Friedrich-Ebert-Straße 68 GmbH, Germany).
Flow cytometry
Cells were incubated with BD Tri TEST™ CD4 FITC/CD8 PE/CD3 per CP antibodies and Simul test™ control γ1/γ2 [fluorescein isothiocyanate (FITC) and phycoerythrin (PE)], mouse IgG, per CP (Beckton-Dickinson, San Jose, CA) as isotype controls for 30 minutes at 37°C in the dark and incubated with lysing solution for 10 minutes, then washed with sheath fluid (FACS Flow™, Beckton-Dickinson) and resuspended in sheath fluid. Cell-surface expression analysis was assessed with the help of a FACS Calibur (Beckton-Dickinson).
NK-cell assay
Purified lymphocytes depleted of adherent cells (5 × 106) were mixed with 5 × 106 K562 cells as a target (effector:target ratio, 1:1) in RPMI-1640 containing inactivated human AB serum. The mixture was incubated for 10 minutes at 30°C, followed by centrifugation at 400g for 5 minutes. The pellet was gently resuspended in 50 μL of RPMI-1640 along with 50 μL of media-agarose mixture (1 mL 3 × RPMI-160 + agarose type I: agarose type V, 1:5, 2%, maintained at 39°C) and smeared onto agarose precoated slides. The slides were allowed to gel for 5 minutes and transferred to plastic dishes containing RPMI-1640 prewarmed to 39°C. They were incubated for 4 hours in a humidified 5% CO2 atmosphere. This was followed by staining for 12 minutes with 0.1% trypan blue, wash with phosphate-buffered saline (PBS), and fixation for 6 minutes in 0.6% formalin. They were washed again and preserved in 0.3% formalin until observation under a visible microscope. The percentage of target-binding cells was scored by counting the number of lymphocytes binding to K562 among 200 lymphocytes. Percentage of dead conjugated target was obtained by counting 100 lymphocyte-K562 conjugates. Spontaneous target cell death was assessed by counting 200 target cells incubated in the absence of effector cells, and this never exceeded 5%. The percentage of active killer cells was calculated by using the following formula:
Absolute numbers were calculated from the absolute lymphocyte count.
Evaluation of IL-2 production
The lymphocytes (105cells/well) were subjected to 48 hours of stimulation by PHA/anti CD3 mAb at 37°C in a humidified, 5% CO2 atmosphere. Cells were pelleted, supernatants of the cultures were collected after 48 hours, and IL-2 production was measured by (enzyme-linked immuno-sorbent assay (ELISA) (Biosource Int, Camarillo, CA), as recommended by the manufacturer.
MTS nonradioactive lymphocyte proliferation assay
Lymphocytes (105 cells/well) were cultured in 96-well plates in RPMI-1640 medium with 10% FCS and stimulated with anti-CD3/PHA in the presence or absence of r-IL2. MTS assay was performed, as previously reported. 21 The plates were incubated at 37°C for 72 hours in a humidified, 5% CO2 atmosphere. Then, 20 μL of the MTS (Promega Corporation, Madison, WI) solution was added to each well. The plate was incubated at 37°C for 4 hours in a humidified, 5% CO2 atmosphere. Absorbance was recorded at the 490-nm wavelength, using a 96-well plate reader. Proliferation index was calculated as: PI = (mean OD anti CD3/mean OD without anti-CD3) × 100.
Thymidine incorporation assay
To confirm the proliferation index, lymphocytes (105 cells/well) were cultured as in the MTS assay, and 0.5 μCi/well of tritiated thymidine (Board of Radiation and Isotope Technology, Mumbai, India) was added during the last 18 hours of the assay. The cells were harvested onto glass-fiber filter paper (Cambridge Technology, Inc., MA, Cambridge USA), using a cell harvester (PHD™ Cell Harvester; Cambridge Technology, and tritiated thymidine incorporation was measured in a liquid beta-scintillation counter (LKB Wallac 1409 Liquid Scintillation Counter, Turkü, Finland) as counts per minute (CPM). The proliferation assay was performed in triplicate wells, and data were expressed as mean CPM and the cell proliferation index calculated.
Enrichment of T-cells
In order to study the T-cell signal transduction proteins, lymphocytes (10 × 106) were cultured in RPMI-1640 medium in the presence or absence of anti-CD3/PHA/r-IL2 and incubated at 37°C for 48 hours. T-cells were enriched by incubation with anti-CD3-coated magnetic beads and separated through MACS T-cell separation columns. T-cells with 95% purity were obtained by this method. The T-lymphocytes thus obtained were used for the preparation of the cell lysates.
Generation of T-cell lysates
Cytoplasmic fractions of the stimulated T-lymphocytes were prepared as reported earlier. 16 Briefly, cells were harvested by brief centrifugation and lysed in a mixture of 1% NP-40, 10 mM Tris (pH 7.8), 150 mM NaCl, 1 mM phenyl methylsulfonyl fluoride (PMSF), 0.4 mM sodium ortho vanadate, 10 mM NaF, and leupeptin at a concentration of 1 mg/mL. NP-40-soluble cell lysates were cleared by centrifugation at 14,000g for 10 minutes at 4°C. The supernatants, represented the cytoplasmic extracts, were removed and stored at −80°C. The nuclear fractions were prepared as reported earlier.22 Briefly, the pellet was washed with PBS and treated with 25% glycerol, 20 Mm HEPES, 0.4 M NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM phenyl methylsulfonyl fluoride (PMSF), 0.4 mM sodium orthovanadate, 10 mM NaF, and 1 mg of leupeptin per mL, sonicated for 30 seconds on ice, and centrifuged at 14,000 g at 4°C for 10 minutes. The nuclear lysates were stored at −80°C.
Western blot analysis of T-cell lysates
Western blotting was performed by running cytoplasmic lysates on 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gel for TCR-ζ and CD3-ɛ and 10% SDS-PAGE for Zap-70, p56lck, and PKC. The nuclear lysates were electrophoresed on 10% SDS-PAGE gel for NFκβ p50, Rel-A, Rel-B, and c-rel. The gel was transferred to a nitrocellulose membrane and blotted with specific primary antibodies (Santa Cruz Biotechnology, Santa Cruz, CA), and blots were developed using ALP-conjugated secondary antibodies (Dako, Carpinteria, CA). The immune reactions were visualized with BCIP/NBT (5-bromo-4-chloro-3-indolyl phosphate/nitroblue tetrazolium).
Results
Surface phenotyping
A total of 72 cervical cancer patients belonging to various stages of the disease and 70 healthy individuals were included in the study. The percentages of subpopulations of T-lymphocytes are shown in Figure 1, and their CD4/CD8 ratio are shown in Figure 2. Surface phenotyping of CD3+, CD4+, and CD8+ cells showed significant decrease in CD3+ (60 ± 5 in stage I, 57.3 ± 6 in stage II, 53.6 ± 6 in stage III, and 38 ± 7 in stage IV in cervical cancer patients versus 56.84 ± 5 in controls) and CD4+ cells (37.5 ± 6 in stage I, 30.7 ± 7 in stage II, 29.6 ± 5 in stage III, and 24.22 ± 4 in stage IV in cervical cancer patients versus 41.5 ± 7 in controls) and the CD4+/CD8+ ratio (1.53 ± 0.29 in stage I, 1.28 ± 0.3 in stage II, 1.2 ± 0.21 in stage III, and 0.905 ± 0.18 in stage IV in cervical cancer patients versus 2.249 ± 0.21 in controls), with a significant increase in the CD8+ cells in cervical cancer patients (24.5 ± 7 in stage I, 23.8 ± 6 in stage II, 24.6 ± 5.3 in stage III, and 26.5 ± 6 in stage IV in cervical cancer patients versus 18.45 ± 4 in controls) (p < 0.001). All the cancer patients irrespective of the stage showed significantly increased values of the suppressor/cytotoxic cells, as shown in Figure 1. The percentage of CD4+ cells did not differ from the control values in patients belonging to stages I and II, while patients belonging to stages III and IV showed reduced levels of the helper cell population. We also noticed a significant decrease in CD4+/CD8+ ratio in cervical cancer patients, when compared with normal controls, and this defect increased with the advancement of the disease (Fig. 2).
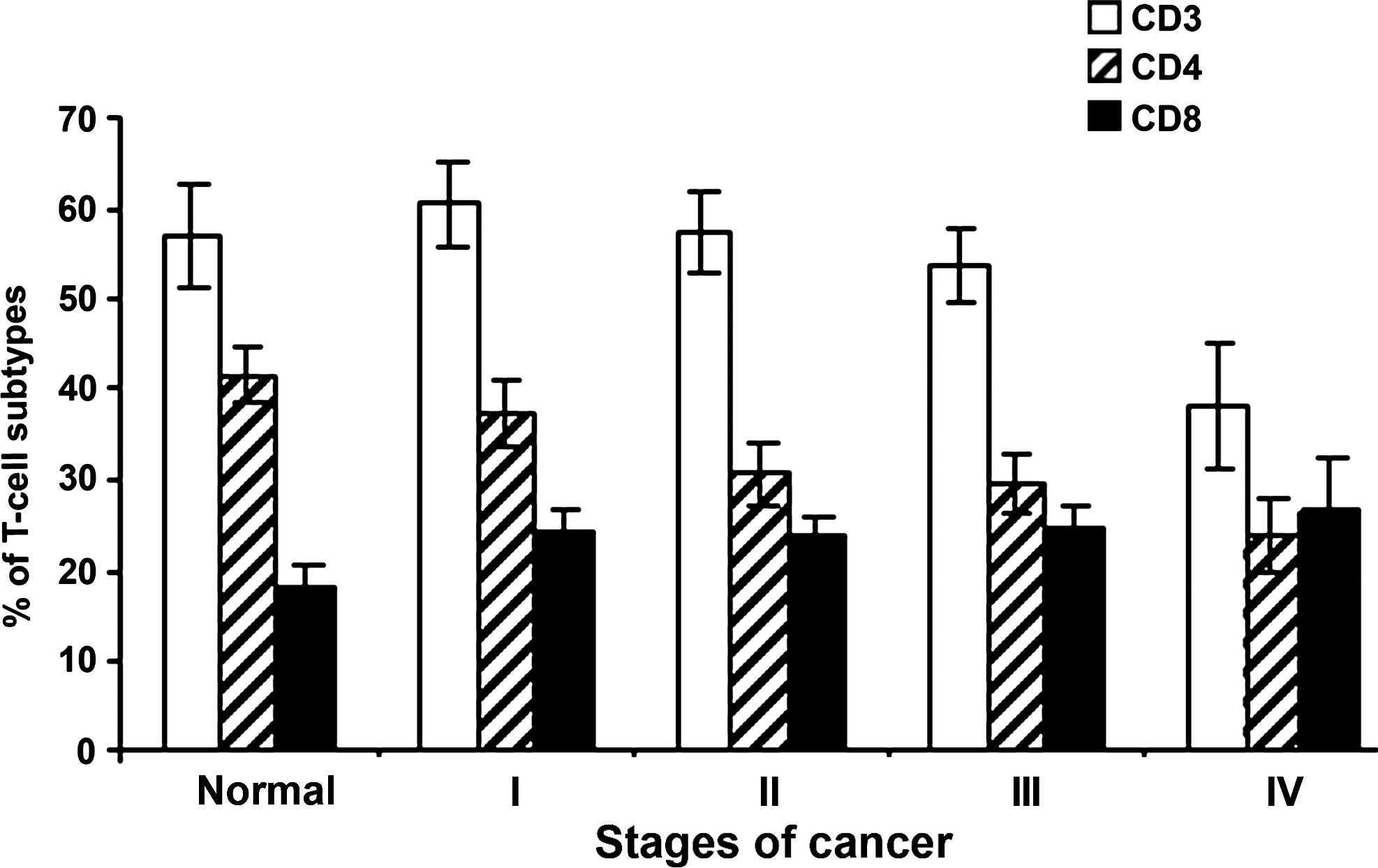
Surface phenotyping of T-cell subpopulations by FACS analysis in patients belonging to FIGO I–IV of cervical cancer, in comparison to normal controls. Cells were incubated with CD3/CD4/CD8 Tritest antibodies and isotype controls for 30 minutes at 37°C in the dark and incubated with lysing solution for 10 minutes, then washed with sheath fluid and resuspended in sheath fluid. Cell-surface expression analysis was assessed with a FACS Calibur (Beckton-Dickinson, San Jose, CA).

Ratio of CD4/CD8 T-cells in various stages of carcinoma of the uterine cervix, compared to normal controls.
NK-cell activity and IL-2 production assay
The results of this study show a significant reduction in the spontaneous cytotoxic potential of the lymphocytes in cervical cancer patients (p < 0.001). The degree of impairment increased with the severity of the disease (Table 1; p < 0.001). Peripheral blood lymphocytes (PBLs) in various stages of carcinoma of the uterine cervix secreted reduced levels of IL-2, when compared to normal controls. This significant defect noticed in the cervical cancer patients was significantly increased in the advanced stages of the disease (Table 1).
All values are mean ± standard deviation.
p < 0.05; ** p < 0.001.
NK, natural killer; IL-2, interleukin-2.
PHA/anti-CD3 stimulated proliferation assay (MTS assay)
Significant decrease in the mean proliferation index (PI) was observed in lymphocytes from cervical cancer patients after stimulation with the anti-CD3, PHA (p < 0.001). Accordingly, the PI of the normal age matched control was 200.2 ± 58 (PHA), 229 ± 64 (anti-CD3), and 248 ± 41 (anti-CD3 in presence of r-IL2) (Figs. 3 and 4). The PI of cervical cancer patients' lymphocyte was 136 ± 33 (PHA), 129 ± 25 (anti-CD3), and 150 ± 34 (anti-CD3 in the presence of r-IL2). The presence of IL-2 showed an augmentation of the proliferative response in 30% of the patients (150 ± 34 in cervical cancer versus 248 ± 41 in controls), showing that lymphocytes from cervical cancer patients were unable to proliferate, to the same extent, as in controls in the presence of the PHA, anti-CD3, and anti-CD3 + r-IL2 (Figs. 3 and 4). Depressed levels of lymphocyte transformation in the PBLs of cervical cancer patients after PHA/anti-CD3 incubation demonstrate the significant impairment in the ability of lymphocytes to respond to antigens and mitogens.

MTS assay showing mean proliferative index of cervical cancer patients and normal control lymphocytes after stimulation with PHA/anti-CD3/anti-CD3 + IL2. Lymphocytes (105 cells/well) were cultured in 96-well plates in, and stimulated with, anti-CD3/PHA in the presence or absence of recombinant interleukin-2 for 72 hours. Then, 20 μL of the MTS solution were added to each well. Absorbance was recorded at the 490 nm wavelength, using a 96-well plate reader, and proliferation index was calculated.
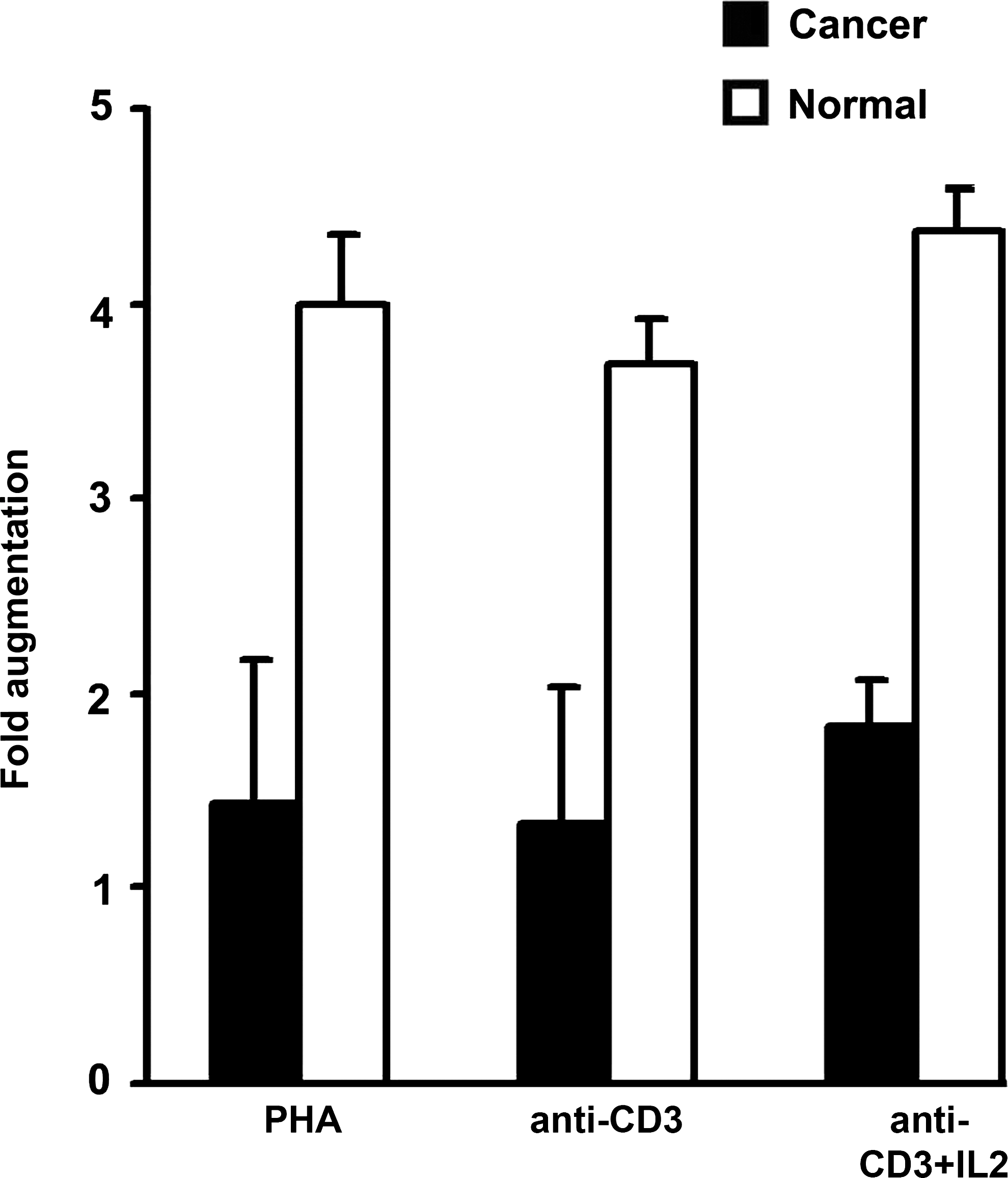
Thymidine-incorporated cell proliferation assay showing mean proliferative index of cervical cancer patient lymphocytes. Lymphocytes were cultured and stimulated with PHA/anti-CD3 in the presence or absence of IL-2 and incubated for 72 hours. Then, 0.5 μCi/well-tritiated thymidine was added during the last 18 hour of the assay. The cells were harvested, and thymidine incorporation was measured. The proliferation assay was performed in triplicate wells, and data are expressed as mean counts per minute, and the cell augmentation index was calculated.
Western blot analysis
The results of the expression of the various proteins in normal controls and cervical cancer patients are shown in Figure 5A and 5B. The results show reduced levels of NFκB p50, Rel A, and Rel B in the nuclear extracts, suggesting that the T-lymphocytes from cervical cancer patients display an impaired ability to translocate these proteins to the nucleus following activation by anti-CD3. These lymphocytes also expressed low levels of the signaling molecules of the T-cell receptor (TCR-ζ, CD3-ɛ, zap-70, p56lck, PKC, NFκB-P50, Rel-A, Rel-B, and c-Rel), when compared to that in the controls.

(
Discussion
Recent studies from several laboratories suggest that alterations in function and expression of signal transduction molecules associated with TCR are responsible for the immune deficiencies observed in various cancers. 1,16 Unfortunately, there are no prospective studies in cancer of uterine cervix examining the clinical significance of an altered CD3-receptor and signal transduction proteins in relation to treatment outcome.
T-cell function and natural cell-mediated cytotoxicity (i.e., NK) has been implicated as an integral part of the tumor-host interrelationships and may play an active role in vivo. The reduction in the number of CD3+ and CD4+ lymphocytes, reduction in the lymphocyte response to mitogens and anti-CD3, and the depression of NK-cell activity in cervical cancer patients demonstrate the inherent defect of the cancer patient's lymphocytes to mount an effective immune response. This depression was stage related, suggesting an influence of tumor or tumor products on these functions. IL-2 is essential for the expansion of antigen-triggered lymphocytes and cytotoxic T-cells, processes necessary for tumor control that are frequently depressed in malignancy. IL-2 secretion was normal in the control groups, but significantly decreased in lymphocytes from the higher stages of disease, when compared to that from normal controls.
The results of the present study showed a significant decrease in the stimulation of lymphocytes from cervical cancer patients. The majority of the cervical cancer patients showed decreased response to PHA and anti-CD3 (Figs. 3 and 4). Production of IL-2 by the lymphocytes was also found to be reduced (Table 1). Exogenous IL-2 could increase the transformation response in all the controls and in 30% of the patients, to different degrees, varying from 10% to 90%. Such a phenomenon has been observed by various other investigators and also from this lab 22 –26 and has been attributed to reduced production of IL-2 and reduced signal transduction. 28,29
The anergy observed in these patients was evaluated further by the capacity of anti-CD3 to elicit the TCR-associated signal tranducing proteins. The results of this study revealed an impairment of the translocation of NFκβ to the nucleus and low levels of other proteins, such as TCR-ζ, CD3-ɛ, zap 70, p56lck, NFκB-P50, Rel-A, Rel-B, and c-Rel, suggesting that the immune impairment noticed could be due to the reduced expression and function of the downstream proteins or defects in TCR itself. Signaling defects in T-lymphocytes of cervical cancer patients emphasizes the fact that their antitumor responses are compromised. This supports the observations of Levey and Srivastava, 1 who reported signaling defects in T-cells from patients with melanoma, renal-cell carcinoma, and colorectal carcinoma, and suggested that decrease in TCR-ζ, zap-70 or p56lck protein expression or an impaired ability to translocate Rel-A from the cytoplasm to the nucleus leads to an inability of T-cells to respond to antigenic stimulation.
Monitoring for TCR-ζ expression is useful for assessing immune competence in these patients and for following changes in immune competence during anticancer therapies. Correlations made between clinical findings, pathologic results, and TCR-ζ expressions in immune cells from different investigators suggest that low/absent TCR-ζ is predictive of poor prognosis and survival in patients with cancer. Thus, TCR-ζ is emerging as a clinically relevant signaling molecule, which also seems to predict a favorable response to biologic therapies and could be helpful in a selection of patients for immunotherapy trials. 26 Among these factors, reduction of TCR-ζ chain activation and inability to translocate NFκβ components from cytoplasm to the nucleus appears to be the main reason for the impaired immune response.
Finally, the present results have strong implications for designing and monitoring treatment strategies for patients with cervical cancer. The molecular defects observed correlate well with immunologic functions, and it will also be of help to predict the ability of tumor-bearing individuals to respond to immunotherapy. Patients without such molecular defects may be more likely to respond to immunotherapy. Reversing this immune defect was possible in 30% of patients by recombinant interleukin therapy, as evidenced from the increased response to anti-CD3 in presence of rIL-2. These data may be of importance for predicting response to treatment of patients with cervical cancer undergoing therapy and may also be useful in generating future immunologic therapies.
Conclusions
To conclude, the results of this study demonstrate an inherent inability of lymphocytes to respond to mitogens and anti-CD3. The results indicate that the impairment of immune response is due to an inability of the T-cells to transduce the signals and also due to the reduced expression of the signaling proteins, especially TCR-ζ, which results in a defective translocation of NFκβ to the nucleus and thereby defective IL-2 secretion and proliferation. This impairment can be corrected in a proportion of the patients with immunotherapy, and these could form the population with potential for interleukin therapy. The results also suggest the impairment to be due to defective signal transduction machinery in cervical cancer patients, hence indicating that these signal-transducing proteins could be potential targets for immunotherapy in cervical cancer patients.
Footnotes
Acknowledgments
The authors thank the Kerala State Council for Science, Technology, and Environment committee (KSCSTE in Trivandrum) of India for their financial support for the project and also wish to acknowledge Tata Memorial Trust for the fellowship offered to SMK to carry out the project.
Disclosure Statement
No competing financial interests exist.