
Abstract
We investigated whether natural killer (NK) cells in the tumor microenvironment have a radiosensitization effect. The radiosensitization effect of combined CpG and Herceptin® (Genentech, Inc., South San Francisco, CA) (CpG/Herceptin), given before or after radiation, was evaluated by using a murine colon cancer cell line overexpressing human HER2/neu, CT26HER2/neu. In vitro radiosensitization effects were investigated by coculture of CT26HER2/neu with splenocytes, CpG, and Herceptin before applying radiation. Tumor cells, cocultured with CpG-pretreated splenocytes and Herceptin, were more vulnerable to radiation damage. In BALB/c mice injected with CT26HER2/neu, CpG/Herceptin administered before radiotherapy was associated with a better retardation of tumor growth than when administered after radiotherapy. The radiosensitization effect was significantly abrogated by NK-cell depletion, indicating that NK cells play an essential role in it. Further, surviving mice treated with CpG or CpG/Herceptin and reverse transcriptase were resistant to renewed tumor challenge, suggesting the presence of an induced immune response to the tumor. Neoadjuvant immunotherapy with CpG/Herceptin may improve response to radiotherapy of HER2/neu-expressing tumors.
Introduction
Immune modulation has been closely associated with tumor growth and reaction to various therapies. 1 –3 The interactions between radiation and immunotherapy are complex. Radiation therapy can be an important adjuvant to immunotherapy by providing danger signals, antigen sources, and activation of innate immunity. 4 Immunotherapy may sensitize the tumor to subsequent radiotherapy and reduce the amount of radiation dose required to eradicate it. 5
Overexpression of Her2/neu is found in 30% of breast cancers and is associated with poor prognosis. 6 On the other hand, it renders monoclonal antibody (mAb) trastuzumab (Herceptin®) active as a single agent or in combination with chemotherapy in the treatment of breast cancers that have Her2 overexpression. 7 Targeted therapy with Her2/neu antibody is known to enhance the radiation effect through the downregulation of the apoptosis threshold. 8
Synthetic oligodeoxynucleotide (ODN), containing unmethylated CpG motifs (characteristic of bacterial DNA), may specifically interact with Toll-like receptor 9 (TLR9) on dendritic cells (DCs) and B-cells, leading to their activation, maturation, and eventual priming of cellular and humoral immunity. 9,10 Direct intra- or peritumoral injection of CpG ODN has been studied extensively, with the expectation that the immune system will select the appropriate tumor antigens and mount an effective response against the tumor. 11,12 The immunostimulatory effects of CpG ODN are broad enough that the tumor microenvironment will support the break in immunotolerance. 13
These activated natural killer (NK) cells could then mediate antibody-dependent cell-mediated cytotoxicity (ADCC) or directly kill NK-sensitive tumor cells. 14 Both agents have been reported to have a radiosensitization effect. 15,16 Several studies have indicated that CpG given with radiation enhances the therapeutic efficacy of the latter apparently through an immune-mediated mechanism. 17,18 The way immunity enhances radiation response is an important question to be addressed, and it is still unclear whether the enhanced response to radiation reported by Mason et al. 18 results from NK-cell activation. In a previous study, we found that the creation of an immunologically favorable microenvironment, as well as the activation of NK cells, are essential for successful radiotherapy. 19 We hypothesize that the interaction of CpG and Herceptin will further attract NK cells into the tumor microenvironment, and that these NK cells are critical for the subsequent radiosensitization effect.
Materials and Methods
Mice and tumor cells
BALB/c mice were purchased from National Taiwan University Animal Center and housed in pathogen-free conditions with 4–6 mice per cage. A BALB/c murine colon carcinoma line overexpressing human HER2/neu, CT26HER2/neu, was used in this experiment. The CT26HER2/neu is a stable human HER2/neu cDNA transfected CT26 cell line. The HER2/neu cDNA gene, with a cytomegalovirus (CMV) promoter in the PCR3.1 eukaryotic expression vector (Invitrogen, San Diego, CA), was transfected into CT26 cells by using Lipofectamine (Invitrogen) and selected by G418, according to the manufacturer's instructions. Tumor cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Invitrogen), supplemented with 10% fetal bovine serum (FBS).
ODNs and Abs
The active CpG ODNs 1826 (sequence TCCATGACGTTCCTGA CGTT) has a nuclease-resistant phosphorothioate backbone (Prisma Biotech, Taipei, Taiwan). A non-CpG-containing ODN was used as a control (GGGGGAGCATGCTGGGGGGG). ODNs were injected peritumorally in a volume of 100 μL to achieve a dose of 100 μg per mice. The humanized anti-HER2 mAb, Herceptin, was obtained from Genentech, Inc. (South San Francisco, CA). Herceptin was administered at a dose of 200 μg per mice, diluted in 100 μL of phosphate-buffered saline (PBS). Polyclonal human IgG (huIgG) was purchased from Sigma-Aldrich (St. Louis, MO). The NK-depletion agent, anti-asialo GM1, was purchased from Wako (Osaka, Japan). The optimal amount of antiasialo GM1 for in vivo depletion was based on recommended titrations of the manufacturer and our preliminary assays of the lactate dehydrogenase (LDH) test.
Clonogenic assay
CT26HER2/neu cells were seeded at 1 × 105 cells per 6-cm dish and grown to confluence overnight. CpG ODNs were added to the dish at a concentration ranging from 1 to 10 μM. After 24 hours, cocultured and trypsinized cells were placed into a 15-mL tube and irradiated with 0, 200, and 400 cGy. Triplicate culture cells were placed in 35-mm Petri dishes (Nunc, Naperville, IL) and maintained at 37°C with 5% CO2 in air in a humidified atmosphere. The cells were then cultured in the incubator for 12∼14 days. Individual colonies (>50 cells/colony) were fixed with a solution containing 25% acetic acid and 75% methanol and stained with 5% Giemsa solution. Colonies were counted by using an inverted microscope.
NK-cell activity assay
NK-cell activity was assayed according to the procedures previously described. 19 Briefly, the YAC-1 cell line (tumoral mouse erythroleukemia cells) was used as a target cell. PKH67-GL (Sigma), a green fluorescent cell linker dye in 1 mM in ethanol, was used for labeling the target cells. Murine splenocytes used as effector cells, obtained after spleen dissociation through a cell strainer followed by erythrocyte lysis, were then added at 2 × 106 cells/well in 2 mL of RPMI medium containing 1 μM of CpG ODNs for 48 hours at 37°C. Control conditions consisted of splenocytes treated with control ODNs at the same concentration. The ratios of effector:target were 50:1, 25:1, and 12.5:1, respectively. NK activity was determined in duplicate by using FACSCalibur (Beckton Dickinson, Mountain View, CA).
In vitro IFN-γ releasing assay
For the in vitro coculture assay, wells of a 24-well, flat-bottomed culture plate were seeded with CT26HER2/neu at a density of 5 × 104 tumor cells/well. Tumor cells were grown to confluence overnight and then treated with 100 μg/mL of Herceptin for 1 hour at 37°C. After washing off unbound Herceptin, splenocytes from BALB/c mice were then added at 2 × 106 cells/well in 2 mL of RPMI medium containing 1 μM of CpG ODNs. Control conditions consisted of splenocytes treated with medium alone, Herceptin-coated tumor cells alone, or CpG ODNs alone. Culture supernatants were harvested after 48 hours and analyzed by using the enzyme-linked immunosorbent assay (ELISA) for mouse interferon-gamma (IFN-γ). ELISA kits for murine IFN-γ were purchased from Endogen (Cambridge, MA, USA).
In vitro radiation-induced cytotoxicity assay
CT26HER2/neu with or without 100 μg/mL of Herceptin-coated were seeded at a density of 5 × 104 tumor cells/well in a 24-well plate. Splenocytes 2 × 106 in 2 mL of RPMI medium were stimulated with or without 1 μM of CpG ODNs for 48 hours. After 4 hours of coculture of splenocytes and CT26HER2/neu cells, adherent CT26HER2/neu cells were washed twice with prewarmed medium to remove splenocytes and antibody and fresh medium was added. Control groups included CT26HER2/neu alone, CT26HER2/neu plus splenocytes, Herceptin-coated CT26HER2/neu plus splenocytes, and CT26HER2/neu plus splenocytes, which had been stimulated with CpG ODNs. Plates containing all groups were irradiated with 8 Gy of radiation. The cells were irradiated with 4-MV X-rays, using a Seimens KDS2 unit at a dose rate of ∼3.0 Gy/min. Culture supernatants were harvested after 48 hours and analyzed by the CytoTox 96 nonradioactive cytotoxicity assay kit (Promega, Madison, WI) for LDH production. The LDH production was measured by calculating the absorbance at 490 nm. Percent cytotoxicity was calculated as follows: (experimental − effector spontaneous − target spontaneous)/(target maximum − target spontaneous) × 100%.
Tumor growth curve
For tumor implantation, female BALB/c mice, 4–6 weeks old, were implanted with 1 × 105 CT26HER2/neu cells into the right flank with a 27-gauge needle on a 1-mL tuberculin syringe. Approximately 12 days later, when the tumors reached ∼4 × 4 mm in diameter, 6 mice were randomly assigned to each of the specific treatment groups. CpG ODNs 100 μg peritumorally (p.t.) and Herceptin 200 μg intraperitoneally (i.p.) were injected per mouse over 3 consecutive days. The control group of mice received no treatment (injected p.t. PBS), i.p. Herceptin alone, or p.t. CpG plus i.p. nhuIgG (200 μg/mice). On day 4, 8 Gy of radiation to the tumor was performed. The mice were anesthetized, and the tumor was irradiated while the remainder of the body was shielded with 5-cm-thick lead blocks. The longest (a) and widest perpendicular (b) tumor diameters were measured at regular intervals with calipers. Tumor volume was calculated from using the following formula: V = 1/2 a2b.20 Mice were sacrificed when the tumor ulcerated or reached a diameter >2.5 cm.
NK-cell depletion
In other experiments, in order to deplete NK cells before treatment, animals were given i.p. injections of 40 μL of antiasialo GM1 (Wako, Richmond, VA) diluted to 200 μL of distilled water when the tumor reached ∼4 × 4 mm in diameter, and this was repeated once 3 days later. The third time of injection was on the day of radiation, and the last injection was 3 days later. Other treatments were the same as described above. Control mice received injections of PBS. This technique resulted in >93% depletion of NK cells in the peripheral blood and spleen, as determined by flow cytometric analysis with an antimurine DX5 Ab (BD Biosciences, Mountain View, CA).
Tumor control probability
To evaluate tumor control rate at high doses of radiation, 1 × 105 CT26HER2/neu cells were implanted into the right hind leg to avoid unnecessary damage to normal tissue. To best assess changes in tumor size, we initiated treatment when the tumor reached 6 mm in diameter, instead of 4 mm in diameter, as used in the experiments detailed above. Treatment consisted of p.t. CpG ODNs alone or with i.p. Herceptin. After 3 dose injections every 24 hours, radiation was delivered in doses ranging from 5 to 75 Gy. One group of mice received only radiation. Mice were checked for the presence of tumor in the irradiated leg at 2–7-day intervals after irradiation for up to 120 days. The dose of radiation needed to produce 50% tumor cures in irradiated mice (TCD50) was determined at 120 days after irradiation. Tumor control probability (TCP) was calculated, as previously described. 21
Tumor rechallenge test
Surviving mice free of tumor 120 days after treatment with CpG plus 50 Gy of RT (n = 7) or CpG/Herceptin plus 50 Gy of RT (n = 8) received 1 × 104 viable tumor cells on the opposite flank. Aged-matched untreated mice received 50 Gy of RT to the right hind leg as a control (n = 11). The mice were examined twice- to thrice-weekly for tumor development for up to 30 days after tumor recalling.
Statistical methods
Comparison of tumor size, IFN-γ secretion, and cell-lysis percentages were carried out by the Student's t-test. Survival was assessed from using the Kaplan-Meier analysis. The TCD50 was calculated from using logit analysis 21 by SigmaPlot 8.0. Differences with P < 0.05 were considered statistically significant.
Results
CpG ODNs had no in vitro effect on CT26HER2/neu tumor cells with or without reverse transcriptase (RT)
As shown in Figure 1, there was no difference in clonogenic survival percentages in the presence of CpG ODNs or control ODNs at each radiation dose level. We found that CpG ODNs neither directly act on CT26HER2/neu tumor cell proliferation nor enhance radiation-induced cell death.

Radiation effect of CpG ODNs on CT26HER2/neu tumor cells. Increasing levels of CpG ODNs had no effect on postradiation clonogenic survival of CT26HER2/neu tumor cells.
CpG ODNs enhanced NK cell-mediated cytotoxicity
The effect of CpG ODNs on natural cytotoxicity within the splenocyte population was evaluated by using the NK-cell-sensitive cell line, YAC-1 (Fig. 2). CpG ODNs increased natural cytotoxicity, compared with control ODNs. These data suggest that CpG ODNs act as positive regulators of NK-cell-mediated cytotoxicity in mice splenocytes.
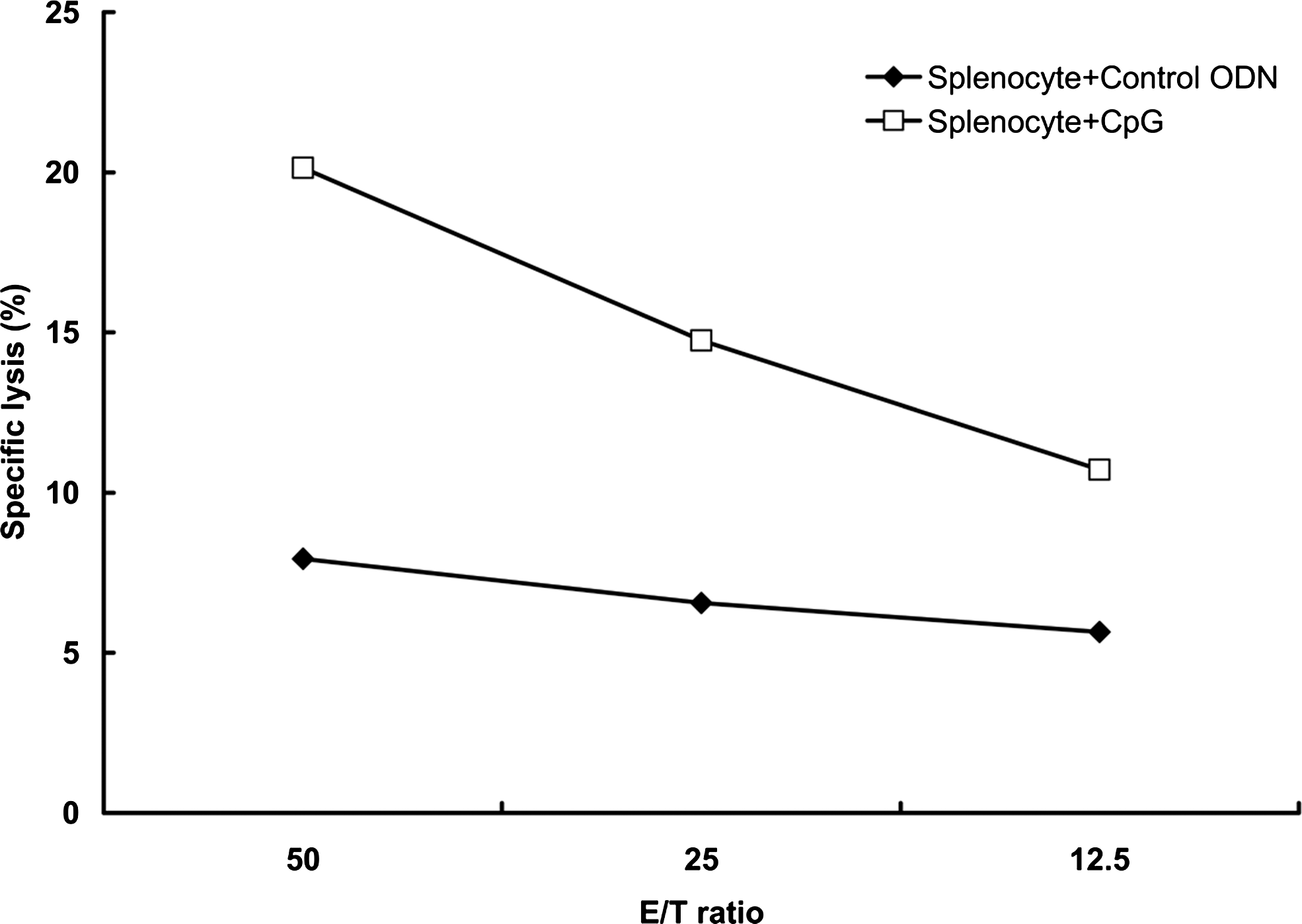
The effect of CpG ODNs on NK cells activity in splenocytes. NK-cell activity was determined by coincubation of effector cells (splenocytes) and target cells (YAC-1) at various “effector-to-target cells” (E/T) ratios, followed by the determination of the PI-positive YAC-1 cells with flow cytometry. Lysis of target cells is shown as a percentage of PI-positive cells. Symbols represent mean of two determinations.
CpG ODNs stimulated splenocytes cultured with Herceptin-coated tumor cells led to enhanced IFN-γ production and radiation damage in vitro
Costimulation of splenocytes with CpG ODNs (1 μM) on Herceptin-coated CT26HER2/neu cells led to potent IFN-γ production (Fig. 3A). The combination of CpG ODNs and Herceptin stimulated more IFN-γ (160 pg/mL) than CpG ODNs alone (130 pg/mL) or Herceptin alone (27 pg/mL) (P < 0.05 and P < 0.01, respectively). The change in radiosensitivity after immunotherapy was also tested. Tumor cells, with or without Herceptin-coating, were cocultured with splenocytes with or without CpG ODNs prestimulation for 4 hours and then irradiated with 8 Gy (Fig. 3B). Four (4)-hour coculture of CT26HER2/neu cells and splenocytes without radiation did not result in detectable cell death. Percentage of cell death was 1.53%, 6.3%, 10.76%, 29.94%, and 44.7% for, respectively, radiation alone, CT26HER2/neu cells cocultured with splenocytes, Herceptin-coated CT26HER2/neu tumor cells cocultured with splenocytes, CT26HER2/neu tumor cells cocultured with CpG ODN-stimulated splenocytes, and Herceptin-coated CT26HER2/neu tumor cells cocultured with CpG ODNs-stimulated splenocytes, all followed by radiation. Activated splenocyte “immunotherapy” renders tumor cells vulnerable to radiation damage.

The effect of CpG ODNs on IFN-γ release by splenocytes and Herceptin therapy (
Neoadjuvant immunotherapy retards tumor growth and induces systemic protection effect
As shown in Figure 4A, radiation resulted in better tumor growth inhibition than treatment with CpG ODNs. Administration of CpG ODNs before RT had a significantly greater tumor growth inhibition effect than after RT, compared to RT alone (P < 0.05). CpG and Herceptin, alone or in combination, all had a radiosensitization effect, which was strongest for the combination of CpG and Herceptin (Fig. 4B). We found that neoadjuvant use of immunotherapy, before RT, is consistently more effective than administering immunotherapy after RT, both for CpG/Herceptin (Fig. 4B) or CpG immunotherapy alone (Fig. 4A). CpG/Herceptin treatment before RT was more effective in delaying tumor growth than CpG/Herceptin given after RT, as well as all other treatment protocols tested with CpG ODNs/Herceptin administered before radiation significantly prolonged survival of CT26HER2/neu tumor-bearing mice, compared to mice treated with RT alone (P < 0.05) (Fig. 4C). Five (5) of 9 mice (55.6%) that received neoadjuvant CpG ODNs/Herceptin were free of tumors 3 months after radiation, as compared to 0% in the RT-alone group.
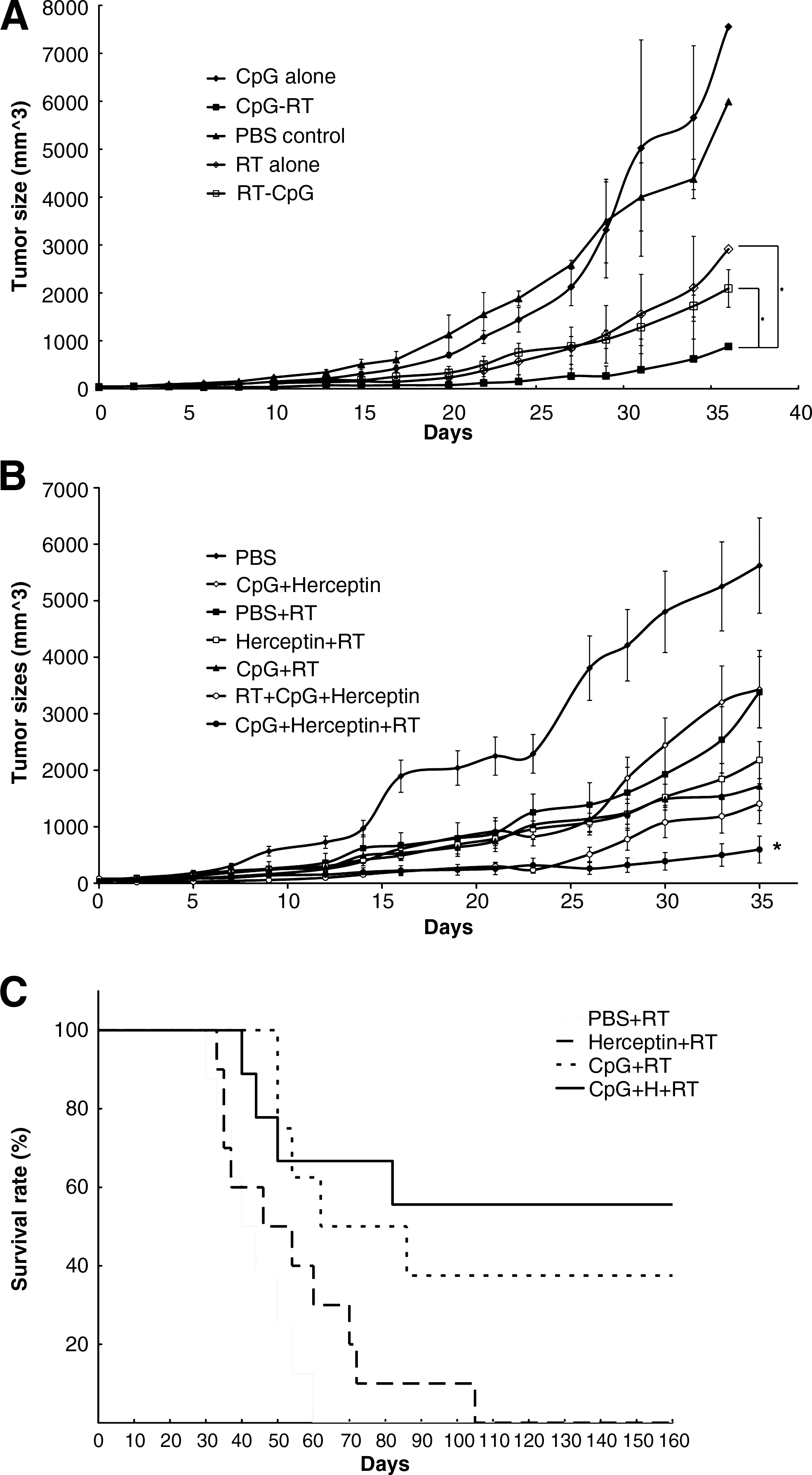
Effect on tumor-growth delay of CpG ODNs combined with Herceptin administered before radiation therapy. Tumor-growth delay assay. The mice were divided into five groups, including local injection of PBS or CpG ODNs, local irradiation with 8 Gy, local injection with CpG ODNs, followed by 8 Gy of RT or local injection with CpG ODNs after 8 Gy of RT (n = 6 in each group), (
Effect of CpG ODNs and Herceptin on TCD50
As shown in Figure 5, the percentage of tumors controlled increased exponentially as the radiation dose increased and reached a plateau at high doses. The TCD50 value was reduced from 54.2 Gy for RT alone to 45.7 and 35.7 Gy for CpG ODN alone and CpG ODNs/Herceptin, respectively.

Tumor control probability. Percentage of tumor control plotted as a function of radiation dose. The CpG ODN and CpG ODNs/Herceptin combination reduced the TCD50 value from 54.2 Gy of RT alone to 45.7 and 35.7 Gy, respectively.
Resistance to tumor cell rechallenge
We tested whether tumor-bearing mice cured by CpG/Herceptin and RT become resistant to tumor rechallenge. As shown in Figure 6, tumor-bearing mice cured either by CpG plus RT or a combination of CpG/Herceptin plus RT were resistant to subsequent tumor cell challenge. In control mice, only 9% of mice were tumor free after 1 × 104 viable tumor cell inoculation. In contrast, 85.7% and 100% of mice were free of tumor in the CpG plus RT group and in the CpG/Herceptin plus RT group, respectively.

Tumor rechallenge after primary therapy. The surviving mice free of tumors 120 days after irradiation combined with CpG ODN treated (n = 7) or CpG ODN plus Herceptin treated (n = 8) were enrolled for the tumor rechallenge test.
The radiosensitization effect of combination of CpG ODNs and Herceptin was abrogated by NK-cell depletion
To determine if there is an NK-cell component responsible for the therapeutic response, NK cells were deleted by the injection of mAbs (antiasialo GM1) against NK cells. Depletion of NK cells was associated with greater growth of CT26HER2/neu tumors, compared to mice that did not receive antiasialo GM1(Fig. 7), suggesting that NK cells are involved in the CpG ODNs and Herceptin-mediated radiosensitization effects. This result demonstrated the important role of NK cells in the therapeutic response to CpG ODNs and Herceptin.
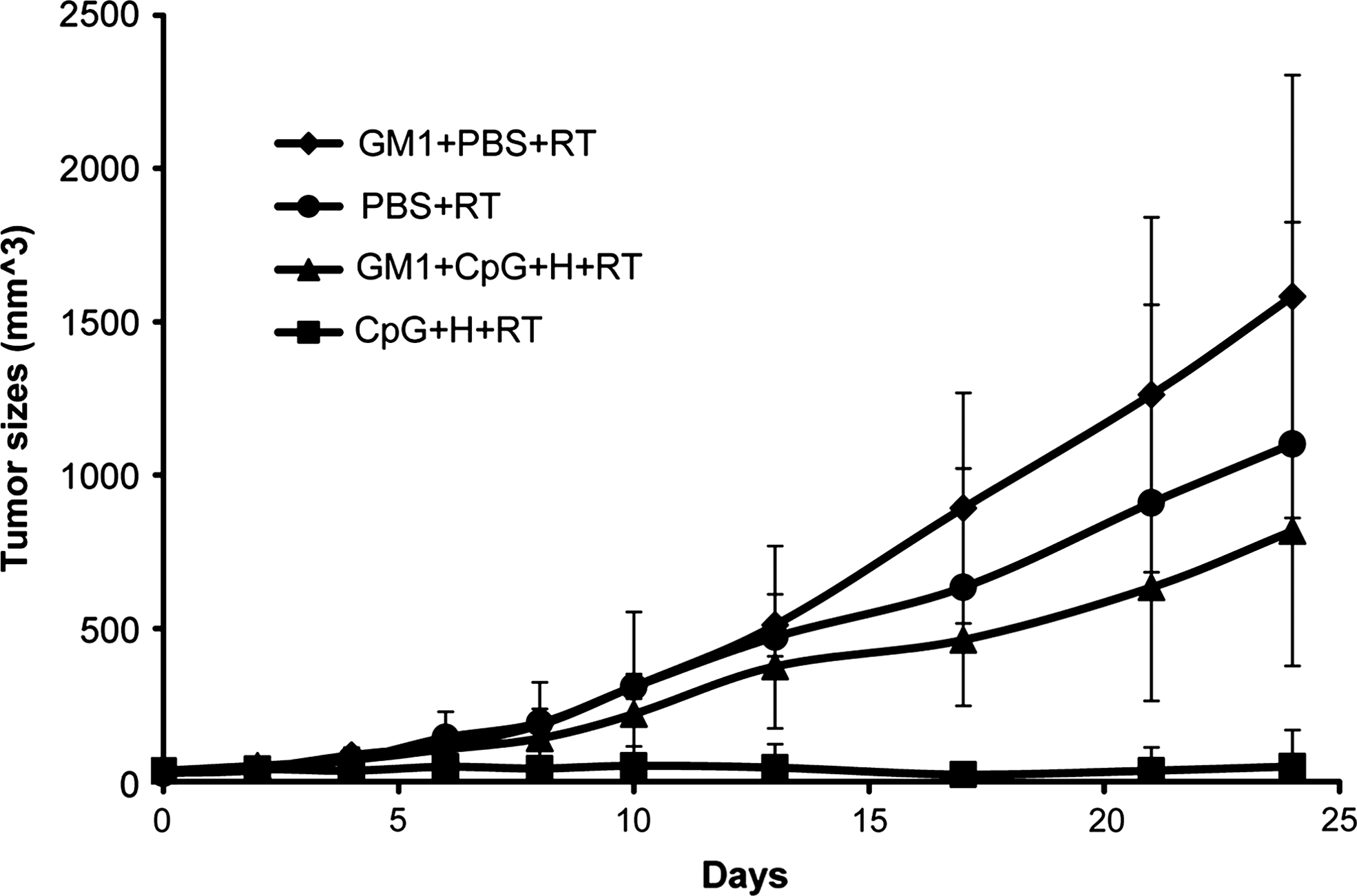
Effect of NK-cell depletion on tumor growth. Mice were divided into four groups, including local injection of PBS, asialo GM1 treated followed by local injection of PBS, local injection with CpG ODNs (100 μg/mice) plus Herceptin (200 μg/mice) or asialo GM1 treated followed by local injection with CpG ODNs (100 μg/mice) plus Herceptin (200 μg/mice). All groups were irradiated with 8 Gy of radiation 24 hours after treatment.
Discussion
Although the current CpG used in this experiment had no direct in vitro radiosensitization effect on CT26HER2/neu tumor cells, results obtained with our animal model suggest an indirect radiosensitization effect mediated through NK-cell activation. A 22% decrease of TCD50 dose and systemic protection can be achieved by the combination of CpG/Herceptin and radiation, and the radiosensitization effect is reduced by NK-cell depletion.
Because CpG stimulates innate immunity and NK cells play a major role in ADCC, the addition of CpG to mAb-based immunotherapy has been suggested by many researchers. 15,22 Recently, Musolino et al. confirmed the role of ADCC on trastuzumab function by revealing that FcγR polymorphism was associated with clinical outcome in patients with breast cancer treated with Herceptin. 23 In the present study, the therapeutic effect of immunotherapy alone, using CpG combined with Herceptin, was not very impressive, but it improved when RT was integrated into the combination. We believe the administration of CpG/Herceptin before RT augments both the radiosensitization effect and the immunomodulating effect. There are three possible mechanisms for this strategy: (1) CpG enhances the direct Herceptin antitumor effect; (2) both agents have radiosensitization effect; and (3) there is an immune-mediated effect. Treatment with CpG/Herceptin significantly enhanced the response of mice bearing large established tumor to subsequent single-dose RT. Treatment with CpG/Herceptin was effective in increasing radiation-induced tumor growth delay and mouse survival rate, compared with radiation alone. Based on these results, we believe that the combination of CpG and Herceptin has potential in RT practice. More important, the mice that achieved local control after combined immunotherapy and RT were resistant to tumor cell rechallenge, suggesting that the treatment was successful in generating systemic antitumor immunity. These observations suggest that systemic antitumor immunity can be generated by local RT once immunotherapy with CpG ODN and Herceptin are integrated.
Our studies demonstrate the role of NK cells in mediating the radiosensitization effects of CpG/Herceptin given before radiation. Depletion of NK cells significantly decreased the tumor growth delay, which suggests a central role for NK cells in the antitumor response. The radiosensitization effect is not completely abrogated by NK cell depletion, which may arise partly from a direct radiosensitization effect of Herceptin and partly from immune modulation mediated by other cells in the immune system. Van Ojik et al. found that the CpG ODN of K-type (CpG ODN 1826 as presently used) works through both NK cells and FcγR1 cells, such as granulocytes and DCs. 24 Meng et al. found that the synergistic effect of combining CpG and RT was not observed in nude mice, underscoring the role of T-cells. 25 Therefore, NK cells, DCs, and T-cells may all play an important role in the systemic protection achieved with the combination therapy investigated in our study.
The interactions between the immune system and radiation are multiple and complex and have been reviewed recently. 26 There are several reasons supporting the addition of radiotherapy to immunotherapy with CpG/Herceptin instead of immunotherapy alone. First, radiotherapy is cytocidal, which may quickly reduce tumor burden and create a less immunosuppressive milieu. Second, irradiation induces tumor cell apoptosis/necrosis with resultant tumor-antigen release for subsequent DC recognition and T-cell response. Third, radiation may increase tumor Fas expression and their susceptibility to cell-mediated lysis. Fourth, radiation may induce the activation of innate immunity through the release of endogenous TLR agonists. 2 Last, the integration of RT into CpG treatment may greatly reduce the risk of CpG-induced tumor promotion. Several epithelial cells and tumor cells were found to have high expression of TLR9. In those cells, stimulation of CpG was shown to increase invasion activity. 27
Few investigators have studied the effect of neoadjuvant immunotherapy on the radiosensitization effect to subsequent RT. We previously provided evidence that the tumor microenvironment, nourished with local released IL-2 and granulocyte macrophage colony-stimulating factor around the tumor site using in situ electrogene therapy, is radioresponsive. 19 We demonstrated in this study that “nontoxic” neoadjuvant immunotherapy significantly increases radiation-induced tumor cell death by culturing Herceptin-coated tumour cells with CpG treated splenocytes for 4 hour before RT. How NK cells render tumor cells more radioresponsive is not fully understood, however.
Conclusions
In summary, our study demonstrates that the combination of CpG ODN and Herceptin improves tumor response to RT, and that NK cells play a pivotal role in this response. The use of neoadjuvant immunotherapy before radiotherapy warrants future clinical trials.
Footnotes
Acknowledgments
This study was supported by grant SKH-8302-95-DR-41 from Shin Kong Wu Ho-Su Memorial Hospital (Taipei, Taiwan).
Disclosure Statement
No competing financial interests exist.