
Abstract
Ascorbic acid (AA) has been reported to inhibit tumor cell growth through the generation of extracellular hydrogen peroxide (H2O2). However, the clinical utility of AA has been limited by relatively low potency and in vivo efficacy. This study reports that the metalloporphyrin, Mn(III) tetrakis(N-methylpyridinium-2-yl)porphyrin5+ (MnTMPyP), has a potent synergistic cytotoxic effect when combined with AA in a variety of cancer cell lines. In the presence of MnTMPyP, the concentration of AA required to inhibit cancer cell growth was markedly reduced. In vitro (cell-free) experiments demonstrated that AA alone enhanced the Fenton reaction that produces cytotoxic hydroxyl radical (HO•); however, this reaction was limited by the low rate by which AA generates H2O2 (Fenton reaction substrate) from O2. MnTMPyP catalyzed H2O2 generation through the AA-facilitated Mn(II ⇔ III)TMPyP redox cycle and thereby markedly potentiated the Fenton reaction. Accordingly, MnTMPyP and AA resulted in increased cellular levels of H2O2 and HO• in cancer cells, which mediate the synergistic cytotoxicity of this combined treatment. This effect was inhibited by cellular enzymes that metabolize H2O2, such as catalase and glutathione peroxidase, suggesting that selective killing of cancer cells deficient in such enzymes can be achieved in vivo.
Introduction
Many cancer cells have abnormalities in the relative level of expression of important intracellular antioxidant enzymes. 1 For instance, prostatic and pancreatic cancer cells have relatively low levels of superoxide dismutase (SOD) and catalase compared with normal tissue. 2 –4 The disturbance of redox balance in such cancers provides a novel opportunity for the development of redox-dependent therapeutic strategies.
Ascorbic acid (AA) is a redox-active agent reported to selectively inhibit the growth of multiple cancer (but not normal) cell lines. 5,6 The underlying mechanism of action of this effect may be the ability of AA to reduce redox-active transitional metals (such as iron and copper), hence facilitating the redox cycling of these metals, which generates various reactive oxygen species (ROS). It has been shown that hydrogen peroxide (H2O2) is generated by AA and is essential for AA-mediated tumor cell killing. 5 AA may also induce the Fenton reaction [Fe(II) + H2O2 → Fe(III) +HO• + OH−] subsequent to H2O2 generation through the Fe(III ⇔ II) redox cycle, which produces the highly reactive and hence toxic hydroxyl radical (HO•). However, the use of iron chelators to block the reaction has revealed inconsistent effects on AA-mediated toxicity, 5,7 suggesting that the Fenton reaction induced by AA is limited. Given that large doses of AA (4g/kg i.p. twice daily) had limited antitumor effects in mouse models, 6 increasing the Fenton reaction may improve the in vivo efficacy of AA as an antitumor agent. The present study hypothesizes that the rate of AA-induced Fenton reaction is limited by the rate of H2O2 generation, because of the absence (or low efficiency) of a metallocatalyst(s), and/or the varying levels of cellular H2O2-metabolizing enzymes (catalase and glutathione peroxidase [GPX]). The experiments described here were designed to test this hypothesis that metalloporphyrin Mn(III) tetrakis(N-methylpyridinium-2-yl)porphyrin5+ (MnTMPyP) would enhance AA-induced H2O2 production and, subsequently, potentiate AA-mediated cytotoxicity via the Fenton reaction.
Known for its ability to eliminate O2 •−, MnTMPyP has traditionally been considered to be an SOD mimic. However, compared with SOD, its in vitro (cell-free) activity is 2 orders of magnitude lower except when in the presence of reducing agents (such as AA, thioredoxin reductase/NADPH, or GSH) that reduce MnTMPyP from its stable state Mn(III)TMPyP to Mn(II)TMPyP. 8,9 Mn(II)TMPyP is then readily oxidized to Mn(III)TMPyP by reacting with O2 •−, to form the Mn(III ⇔ II)TMPyP redox cycle. MnTMPyP is therefore considered to be an oxidoreductase instead of a dismutase. The product of this oxidoreductase reaction has not been documented, but the present study hypothesizes that O2 •− is reduced to H2O2. Other enzymatic properties of MnTMPyP have been reported, including the reduction of O2 to O2 •− under hyperoxic conditions. 9 This effect under normoxia is unknown, but may be potentiated in the presence of AA by enhancing the Mn(III ⇔ II)TMPyP redox cycle. As a result, MnTMPyP may also enhance H2O2 generation by increasing the substrate (O2 •−) concentration. Because some cancer cells have decreased levels of SOD, 2 –4 the anticancer effect of MnTMPyP has been studied both in vitro and in vivo. 10,11 However, in these studies, MnTMPyP alone had no significant cytotoxic effect. The present study hypothesizes that this was due to the absence of metal redox cyclers, such as AA, to generate O2 •−, reduce Mn(III)TMPyP, and enhance Fenton reaction. In the present study, this hypothesis was tested and the potent synergistic antitumor cytotoxicity of MnTMPyP and AA was demonstrated.
Materials and Methods
Cell culture, reagents, and equipments
Hormone-dependent and -independent (subline) LAPC-4 cells were obtained from Dr. Charles Sawyers at UCLA. All other cells were obtained from ATCC (Manassas, VA). Cells were cultured and treated in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS) unless specified otherwise. MnTMPyP pentachloride was purchased from Alexis Biochemical (San Diego, CA). All other reagents were obtained from Sigma-Aldrich (St. Louis, MO) unless otherwise specified. Equipment used in the experiments described below included fluorescent (SpectraMax, Gemini XS) and spectroscopic (SpectraMax 190) microplate reader (Molecular Device, Ramsey, MN), fluorescent microscope (ECLIPSE TS100; Nikon, Melville, NY), image capture system (MicroPublisher 3.3RTV; QIMAGING, Surrey, BC, Canada), ultrapure water system (NANOpure; Barnstead, Waltham, MA), hypoxia chamber (INVIVO2400; Ruskinn/Biotrace, St. Paul, MN), and flow cytometer (FACScalibur; Becton Dickinson, Franklin Lakes, NJ).
Cytotoxicity assay
Cells were plated in a 96-well microplate at a density of 1000 cells/well. After overnight incubation, the cells were treated as indicated in the figure legends and kept in fresh medium for 3 days before staining with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT).
Cell survival assay
Cell survival was determined using the clonogenic assay as previously described. 12 Briefly, subconfluent cells (250–1500 per 60-mm dish) were plated and incubated overnight before treatment. Colonies with over 50 cells were counted at 1–3 weeks after treatment.
Mitochondrial membrane potential determination
Mitochondrial membrane potential (MMP) was measured using rhodamine 123 staining, as previously described. 11 Briefly, 300,000 cells/well were plated the day prior to treatment in a six-well plate. After the treatment, the cells were stained with 5 μg/mL rhodamine 123 dissolved in medium for 1 hour and then photographed, or analyzed using flow cytometry.
Cell death measurement
Cells (300,000 per well) were plated the day prior to treatment in a six-well plate. After the treatment, the cells (including the floating and attached cells) were collected. Cell death was measured using the trypan blue (0.04%; Invitrogen, Carlsbad, CA) exclusion assay and expressed as the percentage of positively stained/total number of cells.
H2O2 measurement
To measure H2O2 generation in vitro (cell-free), a specific and sensitive marker, Amplex-Red (10-acetyl-3,7-dihydroxyphenoxazine; Biotium, Hayward, CA), was used, which is converted to a red fluorescent product (resorufin) upon reaction with H2O2. 13 The Amplex-Red working solution contains Amplex-Red (100 μM) and horseradish peroxidase (HRP II, 1 U/mL) dissolved in PBS or DMEM. The rate of H2O2 production was monitored as ΔRFU (relative fluorescence unit) over 10 minutes (excitation, 540 nm; emission, 590 nm) after mixing the reagents of the reaction.
O2 •− measurement
O2 •− concentration in the reactions was determined using nitro blue tetrozolium (NBT) colorimetry and expressed as the Δ milli optical density (mOD)/minute at 570 nm over 20 minutes of incubation. The reaction mixture contained 0.75 mM NBT.
Cellular ROS measurement
Cellular ROS was measured using a fluorescent probe 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate, acetyl ester (CM-H2DCFDA; Molecular Probe, Carlsbad, CA). Cells (10,000 per well) were plated in a 96-well plate, incubated the next day with 50 μL PBS containing 10 μM CM-H2DCFDA for 30 minutes at 37°C, and then treated as described in the figure legends. The ΔRFU (excitation, 492 nm; emission, 520 nm) over time was measured using a microplate reader.
Catalase and GPX activity assay
The catalase activity was determined using Amplex®Red Catalase Assay kit (Invitrogen, Carlsbad, CA), following the manufacturer's instructions. GPX activity was determined using UV spectrometry of NADPH that is oxidized to NADP− in the presence of GSH, H2O2, and glutathione reductase. 14 The results were normalized using protein concentration and expressed as U/mg protein.
HO• measurement
In vitro (cell-free) HO• generation was determined using the highly efficient and stable terephthalic acid (TA) assay.
15
The reaction reagents were dissolved in calcium- and magnesium-free PBS containing 60 mM TA. The rate of HO
Results
AA and GSH synergized with MnTMPyP to inhibit cancer cell growth
The synergistic effect of MnTMPyP and AA in the hormone-dependent (LNCaP and LAPC-4) and hormone-independent (PC-3, DU-145, LAPC-4) prostatic, pancreatic (PANC-1), and hepatic (Hep G2) cancer cell lines was tested. The treatment with MnTMPyP (up to 1 μM) or AA (up to 1 mM) as a single agent for 2 hours did not inhibit cell proliferation (measured using the clonogenic survival and MTT assays), whereas the combination of MnTMPyP and AA resulted in significantly decreased cell proliferation in a dose-dependent manner in all cell lines studied (Tables 1 and 2). Notably, both assays demonstrated that AA in combination with MnTMPyP significantly inhibited cell growth at a concentration of 0.2 mM, which is achievable in vivo with oral administration of AA. 16
Cells were treated for 1 hour with MnTMPyP (M) at 0.2 μM in the presence or absence of AA (A, 0, 0.2, and 0.5 mM), and survival was determined using the clonogenic survival assay. Data (mean [of triplicates] ± standard error of the mean) are expressed as the survival fraction (%) compared with control.
p < 0.05 versus control, MnTMPyP, or AA (0.2 mM).
p < 0.05 versus control, MnTMPyP, or AA (0.5 mM). The synergistic effect of MA is represented as the ER of MA over A alone. ER1 is the ER of MA/A (M [0.2 μM] and A [0.2 mM]/A [0.2 mM]), and ER2 is the ER of MA/A (M [0.2 μM] and A [0.5 mM]/A [0.5 mM]).
ER, enhancement ratio.
Cells were treated for 2 hours with AA (A, 0, 0.25, and 1 mM) in the presence or absence of MnTMPyP (M, 1 μM). Cell proliferation was measured using the MTT assay at 3 days after treatment. Data are expressed as the % of OD normalized by the corresponding control group.
p < 0.05 versus control, MnTMPyP, or AA (0.25 mM).
p < 0.05 versus control, MnTMPyP, or AA (1 mM). The synergistic effect of MA is represented as the ER of MA over A alone. ER1 is the ER of MA/A (M [1 μM] and A [0.25 mM]/A [0.25 mM]), and ER2 is the ER of MA/A (M [1 μM] and A [1 mM]/A [1 mM]).
ER, enhancement ratio.
Having demonstrated that the combination of MnTMPyP/AA was cytotoxic in a variety of cell lines, the PC-3 cells were used for further study and it was demonstrated that the effect occurred rapidly (within 0.5 hour) and increased over the 2 hours of treatment time (Supplemental Fig. S1A, available online at

Mitochondrial membrane potential (MMP) and cell death in PC-3 cells treated with MnTMPyP and AA. PC-3 cells were exposed for 1.5 hours to AA (1 mM) and/or MnTMPyP (1 μM). (
Notably, GSH (a major cellular reductant) also synergized with MnTMPyP to inhibit PC-3 cell growth, albeit to a lesser extent (Supplemental Fig. S1B, available online at
MnTMPyP and AA synergistically induced mitochondrial damage
Given the rapidity with which morphological changes occurred, and the sensitivity of mitochondria to oxidative damage, the MMP of cells incubated with MnTMPyP/AA was measured using rhodamine 123 staining. Comparing fluorescent and bright-field micrographs immediately after the treatment (1.5 hours), it was observed that some of the cells treated with MnTMPyP/AA were not stained with rhodamine 123 (Fig. 1A). This change was quantitated using flow cytometry, which demonstrated the loss of about half of the rhodamine 123-stained population compared with either untreated control cells or cells treated with AA or MnTMPyP alone (Fig. 1B, 0 hour). The MMP change was not due to cell death, because there was no difference in trypan blue staining between the AA/MnTMPyP-treated cells and those in other treatment groups at this time point (Fig. 1C, 0 hour). However, at 24 hours after the treatment, rhodamine 123 staining was similar in all treatment groups, but there was a slight increase (3.7%) in dead cells in the AA/MnTMPyP-treated group (Figs. 1B and 2C, 24 hours), suggesting that MMP-damaged cells became nonviable over time.

In vitro (cell-free) generation of H2O2 and O2
•− by MnTMPyP and AA. (
MnTMPyP and AA generated H2O2 through the Mn(III ↔ II)TMPyP redox cycle
The enzymatic activity of MnTMPyP as an oxidoreductase requires the presence of reductants.
8
To determine if MnTMPyP generates H2O2 by this mechanism, a simplified in vitro (cell-free) system that contained MnTMPyP in its stable state as Mn(III)TMPyP and AA (or GSH), which are known to reduce Mn(III)TMPyP to Mn(II)TMPyP, was used.
8,9,17,18
The agents were dissolved in PBS at concentrations that were found to be synergistically cytotoxic as shown in Tables 1 and 2. The reduction of Mn(III)TMPyP to Mn(II)TMPyP in this system was confirmed using spectrophotometry (Supplemental Fig. S3, available online at
It has been reported that O2
•− is the active intermediate in the reduction of O2 to H2O2 by AA,
19
and DMEM and unpurified buffers such as PBS contain sufficient amounts of transition metals (such as iron and copper) to catalyze AA oxidation and generate O2
•−.
19,20
Accordingly, it was observed that addition of AA in both PBS and DMEM (with 10% FBS) caused a significant increase of O2
•−, whereas in purified (deionized) water, no O2
•− or H2O2 was generated by AA (Fig. 2D, and Supplemental Fig. S5, available online at
MnTMPyP and AA synergistically increased ROS in PC-3 cells
Using an intracellular ROS probe, CM-H2DCFDA, ROS generation (as the fluorescence change, ΔRFU) of cells exposed to MnTMPyP or H2O2 (positive control) was measured after loading with CM-H2DCFDA. Addition of external H2O2 to the media resulted in a dose-dependent increase of ΔRFU after 1 hour of incubation (Fig. 3A). MnTMPyP (up to 25 μM) and AA (1 mM) alone did not change ΔRFU within the 1 hour of exposure. However, the two combined markedly increased cellular ΔRFU (Fig. 3B, C). Exogenous catalase inhibited the increase of ΔRFU, indicating that this effect was primarily mediated by extracellular generation of H2O2. When cells were exposed to MnTMPyP/AA in PBS, H2O2 generation was also observed (Supplemental Fig. S6, available online at
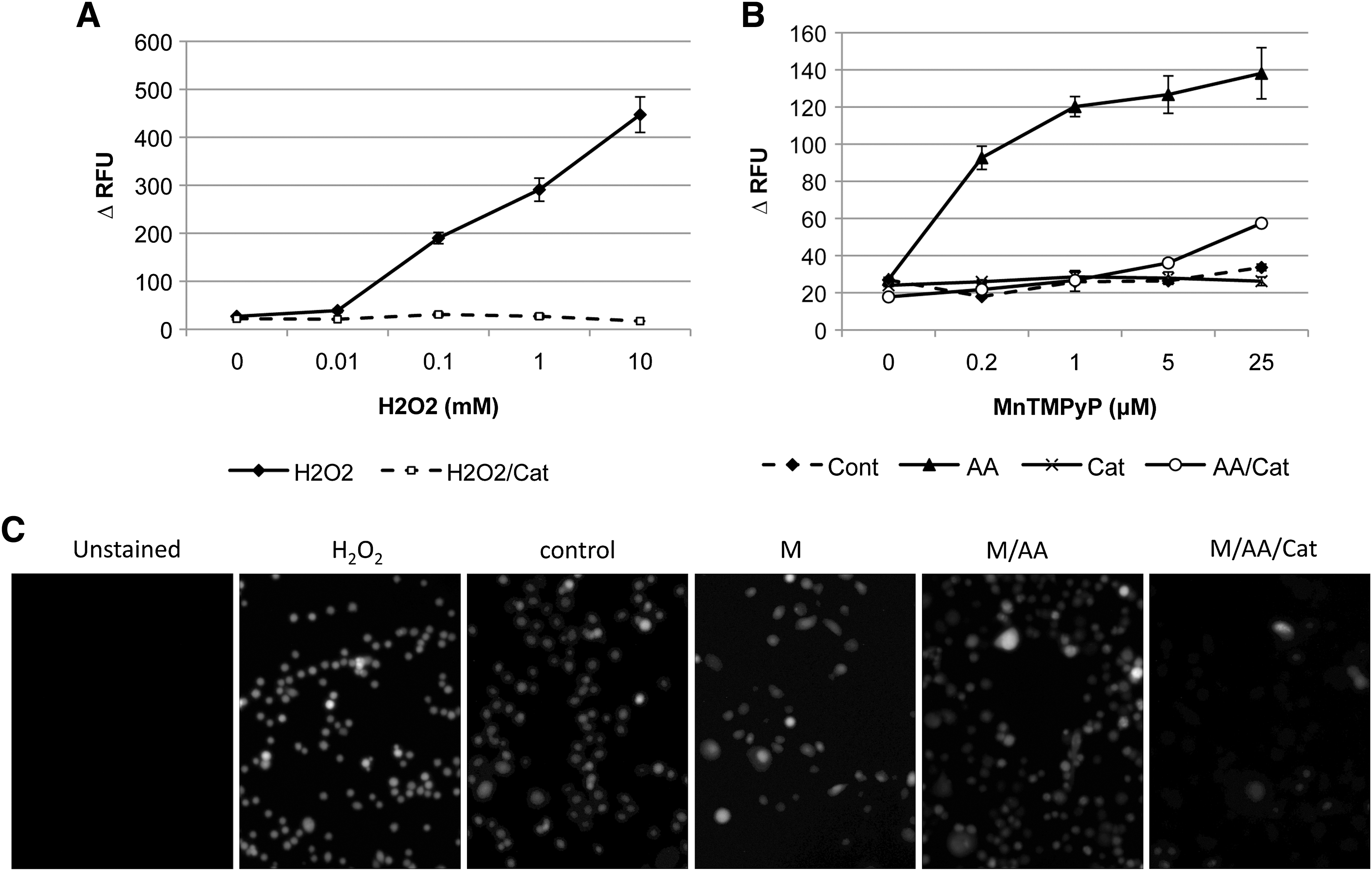
ROS generation in cells treated with MnTMPyP and/or AA. PC-3 cells were incubated with CM-H2DCFDA (10 μM, 30 minutes) and then placed in a medium with different concentrations of H2O2 (
MnTMPyP and AA synergism was dependent on H2O2
Exogenous catalase completely blocked the synergistic effect of MnTMPyP/AA on PC-3 growth inhibition (Fig. 4A), whereas the catalase inhibitor aminotriazole (ATZ) enhanced this effect (Fig. 4B). Both catalase and ATZ alone had no effect on cell growth. AA synergized with external H2O2 to inhibit cell growth in the absence of MnTMPyP (Fig. 4C). This effect was further enhanced by ATZ, which again demonstrates the importance of H2O2 as a mediator of MnTMPyP effects. In a series of prostate cancer cell lines with different levels of activities of H2O2-metabolizing enzymes, the growth of cells treated with MnTMPyP/AA was associated with the catalase and GPX activity (Fig. 4D), suggesting that H2O2 generated by low concentrations of MnTMPyP/AA can be overcome by high activities of endogenous H2O2-metabolizing enzymes.

MnTMPyP and AA synergism was dependent on H2O2. PC-3 cells were exposed for 1 hour to (
MnTMPyP and AA synergism was dependent on the Fenton reaction
The present study hypothesized that the H2O2 generated by MnTMPyP/AA results in the Fenton reaction [Fe(II) + H2O2 → Fe(III) + HO• + OH−] in the presence of Fe(II), which is enhanced by AA that recycles Fe(III) to Fe(II). First, this hypothesis was tested in a simplified in vitro cell-free system that contained the above agents dissolved in PBS at concentrations similar to those used in the cytotoxicity studies (Tables 1 and 2). As a positive control, there was a large increase of HO• within 1 hour of combining AA, Fe(II), and H2O2 (Fig. 5A1). In the absence of AA, the reaction rate was relatively low and only became detectable after 12 hours (Fig. 5A1 and Supplemental Fig. S7, available online at
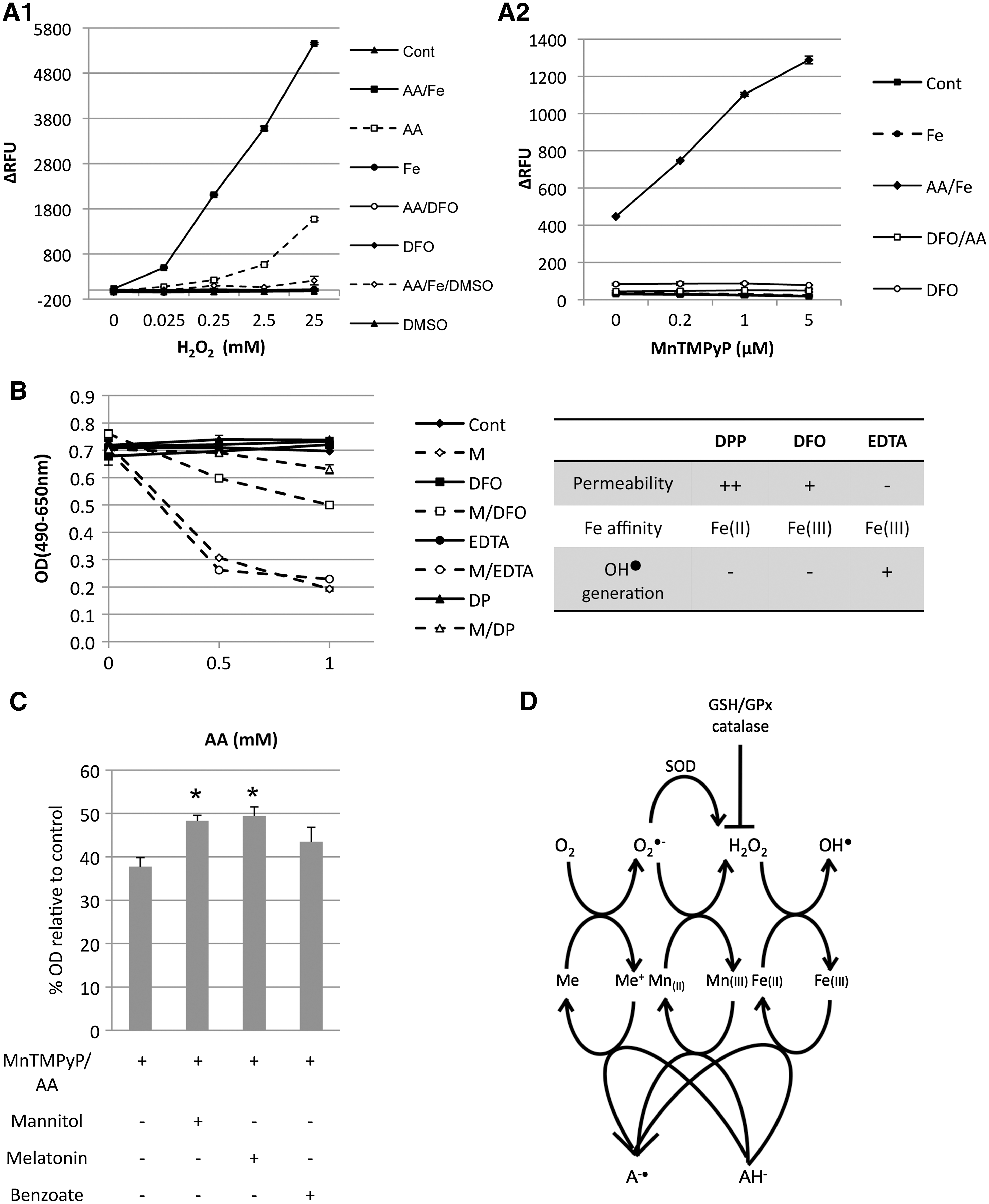
MnTMPyP and AA synergism was dependent on the Fenton reaction. (
To determine if MnTMPyP/AA cytotoxicity was mediated by the Fenton reaction, the iron chelators dipyridyl (DP), DFO, and EDTA were used to remove free iron from the cells. The table in Figure 5B summarizes and compares the membrane permeability of these chelators, as well as their binding affinity to Fe in different oxidation states and the redox activity (HO• generation). Importantly, these chelators did not inhibit O2
•− and H2O2 generation by MnTMPyP/AA (Supplemental Fig. S8, available online at
As O2 is required for HO•-induced cell damage,
23
the effect of hypoxia on MnTMPyP/AA cytotoxicity was also assessed. As expected, hypoxia (0.5 and 2% O2) significantly decreased the growth inhibition induced by MnTMPyP/AA (Supplemental Fig. S9A, available online at
Discussion
Clinical cancer trials in which AA was given orally have yielded inconsistent results. 24,25 More recent studies have demonstrated that AA has in vivo activity against cancer cells only at concentrations achievable with intravenous administration. 6,16 Yet, even with large doses (4 g/kg, twice daily) of intraperitoneal administration, the efficacy of AA was limited in different mouse tumor models. 6 Therefore, alternative strategies are needed to make AA more potent and efficacious and to increase its potential clinical utility. When studying the phenotypes of a K-ras transformed prostate cell line (RWPE-2) overexpressing MnSOD or treated with MnTMPyP, Zhong et al. observed a synergistic inhibition of growth when the cells were treated with MnTMPyP (5 μM)/AA (2 mM) for 72 hours, and they concluded that MnTMPyP enhances the oxidative cellular damage. 26 In addition, Ye et al. recently reported that two other Mn(III) alkylpyridylporphyrins (MnTE-2-PyP5+ and MnTnHex-2-PyP) synergize with AA to induce cytotoxicity. 27 The underlying mechanism of this effect is not clear, but it has been suggested to be mediated by H2O2. Indeed, it has been proposed that this pro-oxidant mode of action of MnSOD and its mimetics may be responsible for their anticancer effects. 28,29 The present study reports a synergistic effect of MnTMPyP with AA, which is much more potent and rapid than that previously reported. The combined MnTMPyP/AA treatment for less than 2 hours resulted in a rapid increase of intracellular H2O2, with secondary mitochondrial injury and morphological changes (within the treatment period), cell death (within 24 hours after treatment), and growth inhibition (within 72 hours after treatment). Importantly, in this combination, the efficacious concentration of AA is markedly reduced to a level that can be achieved and maintained pharmacologically in human blood following oral administration. 16 The pharmacokinetics of MnTMPyP has not been published; however, Spasojevic et al. recently reported the pharmacokinetics of Mn(III) tetrakis(N-ethylpyridinium-2-yl)porphyrin (MnTEPyP), a manganese porphyrin that has an ethyl in lieu of the methyl group in MnTMPyP that is linked with the porphyrin ring. 30 Plasma concentrations of MnTEPyP of 10 mM were reported following a single dose of 10 mg/kg i.p. in mice, which remained above the effective concentration of 0.2 μM in the MnTMPyP/AA regimen for several days. This favorable pharmacokinetic profile suggests that a reasonable dosing regimen may be possible in clinical trials.
In an in vivo environment, the majority of redox active metals, such as iron and copper, are tightly controlled by chelating protein. For instance, the nontransferrin-bound iron (NTBI) measured in body fluids are rarely greater than 1–2 μM and often undetectable in plasma from healthy humans. Given the findings that the AA/MnTMPyP-induced H2O2 generation requires other metal(s) in addition to the manganese in MnTMPyP, in future experiments it will be important to determine if such reaction and subsequent cytotoxicity can be achieved in vivo. Nonetheless, AA-induced H2O2 generation has been documented previously in mouse subcutaneous and tumor extracellular fluid. 6,31 The exact mechanism of the observed redox activity is not clear, but authors of the above articles argue that a putative metaloprotein(s) present in interstitial fluid is responsible for catalyzing the redox reactions. 5
Conclusions
The present study demonstrated that the synergistic cytotoxicity of MnTMPyP and AA is mediated by a cascade of ROS generation (O2 → O2 •− → H2O2 → HO•), which culminates in the generation of highly reactive and toxic HO• radicals (Fig. 5D). AA facilitates each step of the reactions via metal-mediated redox cycling, including the Mn(II ⇔ III)TMPyP redox cycle that markedly enhanced H2O2 generation from O2 •−. As a known reductant, GSH may induce cytotoxicity with MnTMPyP by the same mechanism. However, compared with AA, GSH is less able to reduce transitional metal ions at higher valence at neutral pH. 32 As a result, the cytotoxicity of MnTMPyP/GSH is limited. In addition, at higher concentrations of GSH, the effect becomes less pronounced as H2O2 is more rapidly cleared. It has been reported that GSH, at concentrations (∼0.7 μM) similar to those observed in this experiment, inhibit clonogenic survival in Chinese hamster lung fibroblasts V79 in the presence of copper. 33
Importantly, it was observed that at low concentrations of MnTMPyP (<0.5 μM), the susceptibility of different cell lines to AA/MnTMPyP cytotoxicity is associated with the levels of endogenous catalase and GPX activity. It should therefore be theoretically possible to selectively kill cancer cells with relatively low basal levels of such enzymes within a range of MnTMPyP concentrations in the MnTMPyP/AA regimen that leaves normal cells relatively unaffected. As decreased levels of catalase activity have been reported in prostatic and pancreatic cancer cells, 2 –4 these tumors may be susceptible to this therapeutic approach with an acceptable therapeutic window.
Footnotes
Acknowledgments
This work was supported in part by a grant from the AUA Foundation Research Scholars Program.
Disclosure Statement
The authors claim no conflict of interest associated with this study.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.