
Abstract
The in vitro study of cancer has been made easier by the use of stable tumor cell (TC) lines derived from patients to study antigen expression, immunogenicity, and response to both experimental and conventional therapeutic agents. However, the routine generation of these cell lines in some tumor histologies such as non-small-cell lung cancer (NSCLC) is difficult. In many cases, colonies of TCs do not survive, most likely due to a lack of critical growth factors in cell culture medium. Other times, TC colonies are overgrown by fibroblasts, which appear to have less stringent growth requirements. In some cases, cultures are overgrown by bacteria or mold contained in the biopsy arriving from the surgical or pathology suite. This study presents the characteristics of three new NSCLC cell lines and associated autologous clones generated from both adenocarcinoma and squamous cell carcinoma tissue. Different culture media and variable techniques were used to generate these stable TC lines. Limiting dilution analysis resulted in numerous clones, some of which displayed heterogeneity in terms of growth, antigen expression, and the ability to release cytokines. The successes and failures associated with generating TC lines are discussed in this article. Both parental cultures and related clones serve as critical reagents for the continued study of the cellular immune response to NSCLC.
Introduction
Surgery is extremely important in the treatment of non-small-cell lung cancer (NSCLC). 1,2 When performed prior to radiation, chemotherapy, and/or immunotherapy, surgery increases the chances for survival of patients with improvement in their quality of life. 3 Surgery not only reduces tumor burden, but the tissue obtained can also be crucial in learning about the molecular biology of disease, tumor biology, as well as the immunologic response to disease. This knowledge helps improve diagnosis, disease management, and future therapies. Unfortunately, not all patients are surgical candidates, thus the knowledge that could be gained is limited. 4,5
Biopsies from all solid tumor histologies contain viable, apoptotic, and necrotic tumor cells (TCs) as well as variable numbers of host leukocytes; both activated and resting. 6,7 Tumors also contain fibroblasts and endothelial cells, which make up the stromal tissue. 8,9 The interaction of these cells make up the tumor microenvironment. Heterogeneity in the number, type, and viability of the tumor and tumor-associated cells is affected by prior therapies, stage of disease, and anatomic location of the tumor. For these reasons, isolating consistent numbers of viable TCs from biopsies for tumor biology studies can be challenging. Thus, development of consistent methodologies to derive long-term stable TC lines for use in both molecular and cell biologic studies has been an important focus of research. 10 –14 These cell lines can provide a continuous source of cells expressing tumor associated antigens (TAA) and displaying a metabolic phenotype characteristic of the transformed state. In some cases, a small number of TCs isolated from fine needle aspirates is adequate for the development of a stable cell line as shown in melanoma 15 and breast cancer. 16
NSCLC is a major killer of men and women around the world. 17 –19 Generation of long-term stable TC lines in NSCLC is also characteristically very difficult. Apart from the early description of NSCLC TC lines generated by Gazdar and Oie 20 and Oie et al., 21 few reports 22 –24 detail reproducible methodologies to generate in vitro lines from solid tumor biopsies. The present study aimed at developing long-term stable lines to support vaccine immunotherapy studies. 25,26 Generation of NSCLC lines from biopsies of NSCLC, including squamous cell carcinoma, adenocarcinoma, and large cell carcinoma, was attempted. The Cooperative Human Tissue Network (CHTN) and the University of Kentucky hospital provided the biopsy material. Samples were obtained from biopsies of metastatic solid tumors and pleural fluid lavage. A variety of techniques and different cell culture media were used. The generation of three new NSCLC lines and autologous clones by limiting dilution is reported here. Some differences were noted between the parental lines and clones with regard to growth, cytokine secretion, and antigen expression. These cell lines also serve as a critical resource for ongoing studies on the relationship between chemotherapy resistance and the development of specific immunotherapy reagents.
Materials and Methods
Processing of solid tumors for cell culture
Pleural fluid samples and biopsies of NSCLC tumors with accompanying normal adjacent tissue (NAT) were purchased from CHTN. A few samples were obtained from the University of Kentucky hospital with IRB approval. Solid tumor biopsies were finely minced using scalpels under sterile conditions. The minced tissue samples were mixed with enzyme medium (40 mL per g of tissue) and digested to single cell suspensions as previously described. 27 The enzyme medium was composed of RPMI 1640 (Gibco BRL) supplemented with DNAse (30,000 U/mL), collagenase (1 g/L), hyaluronidase (100 mg/L), penicillin (50 μg/mL), streptomycin (50 μg/mL), amphotericin (50 μg/mL), and gentamycin sulfate (10 μg/mL). Following digestion, the samples were strained through a sterile 70-μm mesh filter to remove clumps of fibrous material, centrifuged to collect cells, and washed with phosphate-buffered saline (PBS) containing 5% fetal bovine serum (FBS) to neutralize residual enzymes in the digestion medium. The cells were resuspended in X-Vivo 15 serum-free medium, counted, and assessed for cell viability by trypan blue exclusion technique.
Pleural fluid samples, usually as much as 1 L, were removed from sterile collection vessels and centrifuged at 250 g for 10 minutes, resuspended in PBS, and washed three times. Solid tumor, NAT, and pleural fluid that were <60% viable as assessed by trypan blue exclusion technique were enriched for viable cells using Ficoll Hypaque gradients as previously described. 28 Cells obtained from tumor biopsy, NAT, or pleural fluid that were not used in the initial cell culture studies were cryopreserved by standard techniques developed in the current authors' laboratory. 29
Cell culture medium
The TC line cultures were attempted in two different media,either complete RPMI (CRPMI) 27 or ACL-4. 21 CRPMI consisted of RPMI 1640 (Gibco BRL) supplemented with 10% heat-inactivated (56°C, 30 minutes) FBS, 50 μg/mL penicillin, 50 μg/mL streptomycin, 10 μg/mL gentamycin sulfate, 2 mM l-glutamine, 100 μM NEAA, 1 mM sodium pyruvate, 1 mM HEPES buffer, and 50 μg/mL amphotericin. ACL-4 medium consisted of D-MEM and HAMS F-12 basal medium (Gibco BRL) mixed 50:50 V/V and supplemented with: 0.2% human serum albumin, 1 ng/mL human epidermal growth factor, 10 μM ethanolamine, 10 μM phosphorylethanolamine, 20 μg/mL insulin, 10 μg/mL transferrin, 25 nM sodium selenite, 50 nM hydrocortisone, 0.1 nM triiodothyronine, 50 μg/mL penicillin, 50 μg/mL streptomycin, 10 μg/mL gentamycin sulfate, 2 mM l-glutamine, 100 μM NEAA, 1 mM sodium pyruvate, 1 mM HEPES buffer, and 50 μg/mL amphotericin. Cholera toxin (1 ng/mL) was also added in some instances to discourage the growth of fibroblasts.
Generation of TC lines and clones
For initial growth attempts, single cell suspensions obtained from the biopsy were seeded in T25 flasks coated with Rat-tail Collagen, Type 1 (BD Biosciences). The collagen was diluted to 50 μg/mL with 0.02 N acetic acid and added liberally to the culture flasks to coat the bottom evenly. The flasks were then incubated at room temperature for an hour, at which point the liquid collagen was removed. The plates were rinsed once with PBS to remove any residual acid and used for culture immediately. Single cell suspensions were added at 1.0 × 106 cells per flask into 10 mL of cell culture medium. The flasks were incubated at 37°C and 5% CO2 in air and the growth of TC colonies examined daily. Initially, cells adhered between 24 and 48 hours, when nonadherent cells were washed free from the flask. Overall, generation of TC lines was not successful unless the developing cell lines maintained adherence to plastic.
Parental cultures of the TC lines UKY-29, UKY-116, and UKY-349 were cloned by limiting dilution. In brief, cells were harvested from culture flasks during the log phase of growth, counted, and assessed for viability by the trypan blue exclusion technique. The cells were then diluted to give a final number of 1 or 10 cells per 200 μL of appropriate medium, and the 200 μL volume was added to wells of 96well, flat-bottom sterile cloning plates. The medium used was either CRPMI or ACL-4. Plates were assessed for growth beginning day 10, and clones were harvested using Trypsin-Versene to remove adherent cells before passage to 48- or 24-well plates. In general, wells seeded at 10 cells/well appeared confluent between days 21 and 28; those seeded with 1 cell required 30–45 days. Further expansion of clones and parental lines was done using T75 cm2 culture flasks.
Determination of antigen expression
TAA expression was monitored either by cell surface phenotyping when monoclonal antibodies were available, or by RT-polymerase chain reaction (PCR). In many cases, both techniques were used for verification purposes.
TCs were assessed for cell surface phenotype by direct or indirect immunofluorescence and analyzed using a fluorescence activated cell sorter (FACS). The antibodies used to detect TAA were directly labeled with fluorescein isothiocyanate or phycoerythrin or were unlabeled and used in indirect labeling studies that use a fluorescent tagged secondary antibody. The antibodies used in this study included anti-Her-2neu, anti-cytokeratin, anti-vimentin (all obtained from BD Pharmingen), and anti-CEA (Zymed). In brief, TCs were washed, counted, and adjusted to 2 × 105 cells per tube in 50 μL of staining buffer (PBS containing 2% FBS and sodium azide). The cells were stained for 30 minutes on ice, washed two times in full volume buffer, fixed with 500 μL of 1% paraformaldehyde, and analyzed using FACSCAN and CellQuest software. Data are presented as % positive cells.
For RT-PCR, RNA was harvested from TC lines using standard methodology. In brief, 5 × 106 TCs (parental or clones) were pelleted at 400 g for 5 minutes and lysed with 1 mL of Trizol, with subsequent steps described by the manufacturer. RNA quantity and quality were assessed by spectrophotometry and gel fractionation. All RT-PCR were run using the Promega Access RT-PCR system. A master mix containing nuclease-free water, AMV/Tfl 5x reaction buffer, 0.2 mM dNTPs, 1 mM MgSO4, 0.1 u/μL AMV reverse transcriptase, 0.1 u/μL Tfl DNA polymerase, and 1 μM each of the downstream and upstream primers was added. Following PCR, the reactions were fractionated on a 1.6% agarose gel containing ethidium bromide. DNA Primers were prepared by Integrated DNA Technologies, Inc. (Coralville, IA), using previously published sequences.
Cytokine release analysis
TCs were analyzed for their ability to secrete cytokines following 24 hours of incubation at 37°C and 5% CO2 in air. In brief, TCs were incubated at 1.0 × 106 cells/mL of medium in 24-well plates. After 24 hours the supernatants were removed and analyzed for cytokines using ELISA kits supplied by BD Pharmingen. The data are presented as pg/mL cytokine released per 1 × 106 cells. The cytokines routinely tested were interleukin-10 (IL-10) and transforming growth factor β (TGF-β).
Generation of cytolytic T cells, cytotoxicity, and ELISPOT analysis
Cytolytic T cells (CTL) were generated from UKY-116 fresh tumor using mixed lymphocyte tumor cell (MLTC) culture as previously described. 30 In brief, the UKY-116 TC line was transfected with the gene encoding the lymphocyte costimulatory molecule CD80. Following selection in G418, expression of CD80 (UKY-116.7) was confirmed by cell surface phenotyping. More than 95% of the UKY-116 TCs expressed CD80 (data not shown). Following irradiation of UKY-116.7 (10,000 rads), MLTCs were established using single cell suspensions obtained from thawed, fresh UKY-116 tumor biopsy and UKY-116.7 at a 10:1 responder-to-stimulator ratio. The medium used was CRPMI with recombinant IL-7 (10 ng/mL) as a growth factor. Following 10 days of culture, T cells were harvested, counted, and reestablished in culture using fresh, irradiated UKY-116.7 and IL-7. After 2 days of additional culture, 10 ng/mL of recombinant IL-2 was added to the cultures. T cells were restimulated at 10-day intervals between days 30 and 45 until functional analysis. The levels of CD8 T cells were determined by flow cytometry, and the lytic reactivity and fine specificity of anti-UKY-116 CTL were determined by standard 4-hour 51Chromium release microcytotoxicity assays as previously described. 30
Anti-UKY-116 T cells were also analyzed using interferon γ (IFN-γ) ELISPOT assay for specific cytokine release. 26 ELISPOT plates were coated with anti-IFN-γ antibody in Dulbecco's PBS for a minimum of 4 hours. The plates were washed in PBS and blocked with blocking medium (450 mL RPMI, 50 mL HI hu AB serum, 5 mL HEPES, 5 mL Pen/Strep–filtered) for at least an hour. Following blocking, the plates were washed, and CTL and antigen targets were added at a ratio of 10:1. The plates were then incubated at 37°C for 20 hours, washed, and indicator antibodies and enzyme development system added to complete the biologic reaction. The absolute number of IFN-γ-producing lymphocytes (spots) was counted using an automated Zeiss Axiomat microscope and data analysis system.
Results
NSCLC TC lines
In this study, a previously established NSCLC line, 1650, was used as a control. 1650-TC was graciously provided to the current authors' laboratory in 1995 by Dr. Herb Oie, National Cancer Institute. This stable line is used as a source of apoptotic bodies in vaccine studies 25,26 and has been established as a master cell bank by the lab, verified viral and pathogen free through appropriate testing and analysis. Published immunologic and cell surface antigen characterization reports 31 were used in the current study for comparative analysis.
Sixty-two samples of NSCLC were processed, the majority being derived from solid tumors obtained from CHTN. Fresh tumor tissue or cryopreserved samples of tumor biopsies were placed in cell culture in either CRPMI or ACL-4 medium. The results represent overall success with both medium conditions. In 6 of the 62 cases (9.6%), short-term lines were obtained (UKY- 65, 97, 107, 123, 160, and 343), which contained colonies of TCs early in culture that either died or were overgrown by fibroblasts prior to passage to a larger flask (see Fig. 1). Only 3 of the 62 samples studied (4.8%) resulted in stable, long-term lines. The adenocarcinoma cell lines UKY-29 and UKY-349 were derived using CRPMI. UKY-116, a squamous carcinoma cell line, was derived using ACL-4 medium (see Fig. 2). In 45 of the 62 biopsies tested (72.5%), there was no growth of TCs but an abundance of fibroblasts. In 8 cases (12.9%), cell cultures became contaminated with microorganisms (bacteria, mold, and/or yeast) and could not be rescued for further development of TC lines.
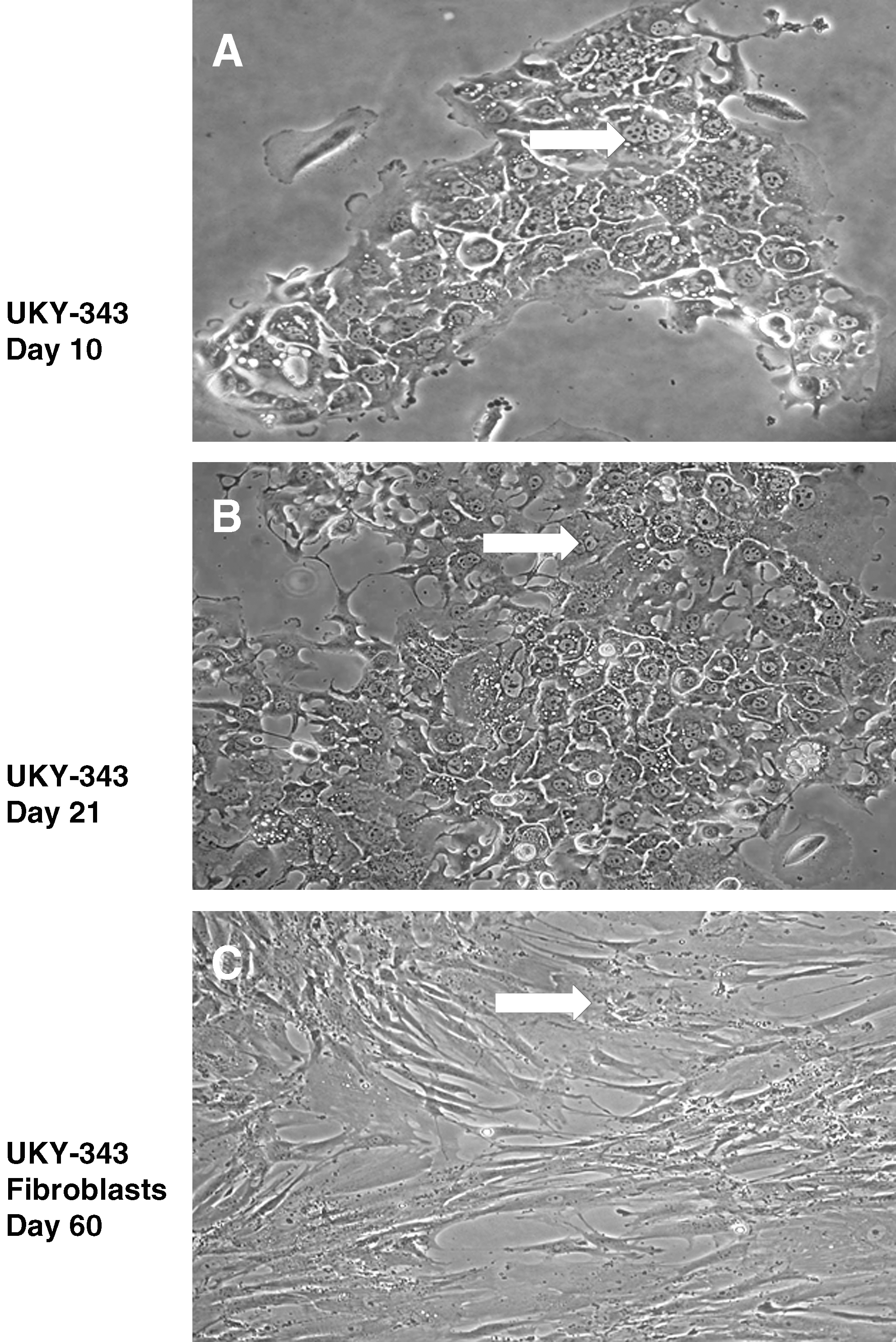
Representative micrographs (40 × magnification, Nikon Coolpix images) illustrating the morphologic characteristics of a short-term NSCLC cell line, UKY-343. TCs from UKY-343 biopsy were established in cell culture in T25 cm2 flasks. (
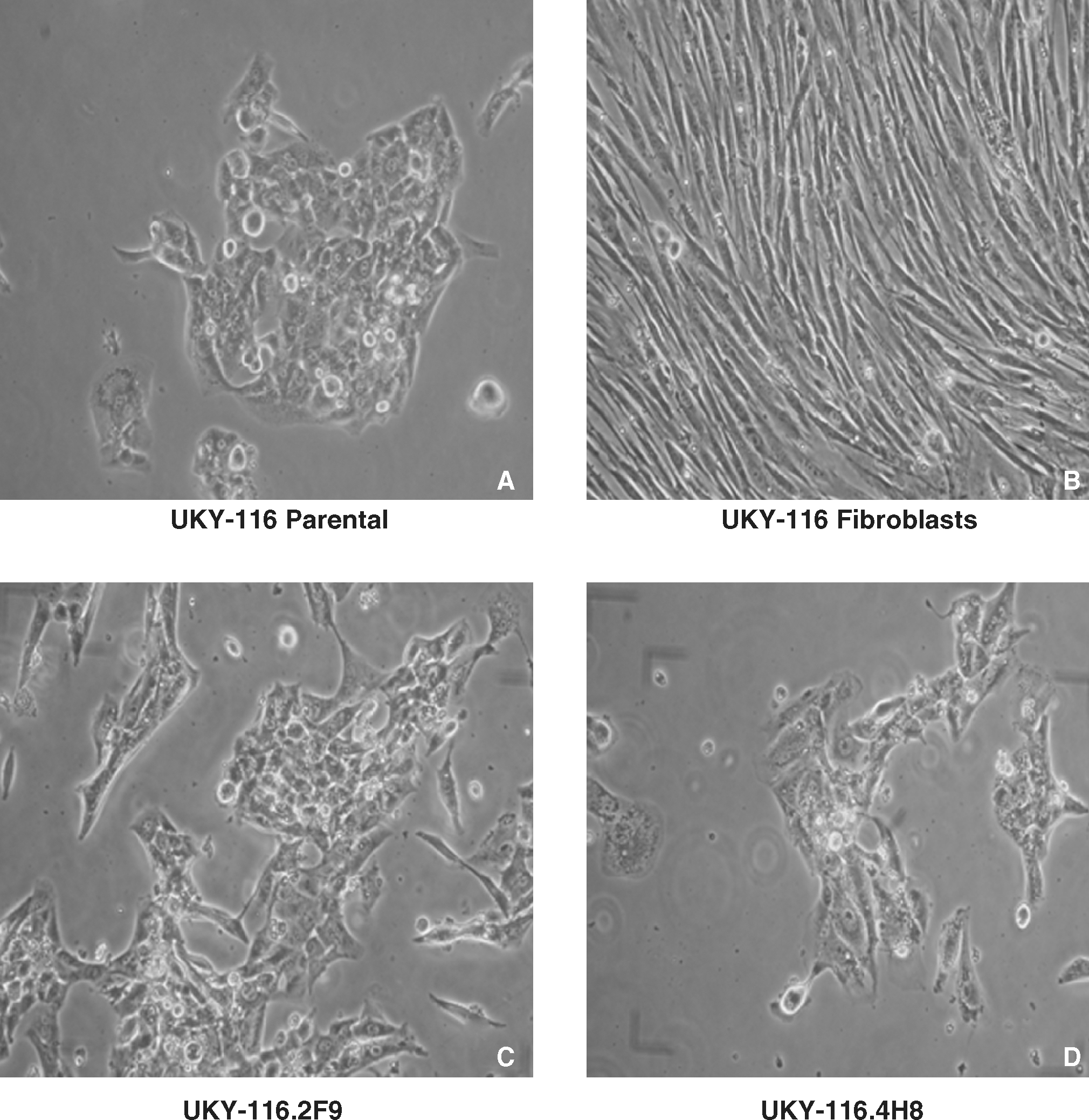
Micrographs (40 × magnification, Nikon Coolpix images) representing the UKY-116 cell line and representative clones derived from 1 cell/well cultures. (
Figure 1 shows the development of the short-term TC line UKY-343 derived from a patient with adenocarcinoma. This result occurred 9.6% of the time. If in vitro passage was not continued, this TC line would have been mistakenly concluded to be stable. TCs in the selected colony appeared to have undergone proliferation as indicated by an increase in colony size between days 10 and 21 (52 cells at day 10, Fig. 1A; 126 cells at day 21, Fig. 1B). However, by day 60 (Fig. 1C), colonies of TCs disappeared with the apparent outgrowth of long, spindle-shaped cells having a morphology characteristic of fibroblasts. The overall morphology of the colony changed, taking on a fibroblastic appearance. Beyond this point, there was no indication of TC colonies in any part of the culture flask. With time, all cells ceased to expand and could not be passaged.
The squamous carcinoma cell line UKY-116 was derived from a biopsy obtained from CHTN. The sample was generated in ACL-4 medium. The parental TC line contained a solitary squamous cell colony (Fig. 2A). The clonal cell lines UKY-116.2F9 (Fig. 2C) and UKY-116.4H8 (Fig. 2D) were obtained by limiting dilution analysis at 1 cell/well. A clone of fibroblasts was isolated in a separate well of 96-well plates (Fig. 2B). Clone UKY-116.2F9 was unique in that its growth kinetics were greater than those of the parental line, while UKY-116.4H8 was the slowest grower of the UKY-116 clones (see Fig. 3). However, no obvious differences in morphology, reflecting growth potential, were evident.

Growth results for 1650-TC, UKY-116, UKY-29, and associated clones. T175 cm2 cell culture flasks were seeded with 1.0 × 106 TCs in 50 mL of appropriate medium. Flasks were observed on a daily basis. When cells reached 75%–100% confluence, TCs were trypsinized, counted, and assessed for viability by trypan blue exclusion. New flasks were then established to provide cells for additional analysis. (
All the cell lines described in the current report were adherent and had some distinguishing morphologic characteristics. In general, smaller, more compact cells represented the squamous cell carcinoma line UKY-116, while larger, more vacuolated cells represented the adenocarcinoma lines 1650-TC, UKY-29, and UKY-349. The TC lines grew as discrete colonies, which, with time, combined with adjacent tumor colonies to form an adherent confluent layer of TCs. During proliferation, TCs appeared to become nonadherent and rounded, and replicated to daughter cells, which then readhered to the plastic. In all cases, distinguishing fibroblasts from TCs in early cultures was difficult until obvious TC colonies appeared. As cell culture time elapsed, fibroblasts were easiest to identify based on their cell shape. They grew as discrete sheets containing swirls of compacted elongated cells (see Fig. 2B).
Growth of parental lines and clones
Following derivation in T25 cm2 culture flasks, the three TC lines developed, UKY-29, UKY-116, and UKY-349, were expanded and passaged in T175 cm2 culture flasks. After the lines were confirmed to be stable, each was cloned by limiting dilution to study TC heterogeneity. UKY-116 and UKY-349 were cloned at relatively early passage numbers (passages 10 and 12, respectively), and UKY-29, derived in 2000, 31 was cloned at passage 102. TCs were cloned at 1 and 10 cells/well. After 30–45 days in the cloning plates, representative clones were expanded and used in studies on the growth, cytokine secretion, analysis of antigen expression, and sensitivity to CTL recognition of cells by lytic and cytokine release assays.
TC line UKY-116 showed heterogeneity in the ability of the clones and the parental line to grow (Fig. 3A). In all cases, UKY-116 and clones selected from 1 cell/well cloning plates did not grow to the levels observed using 1650-TC (Fig. 3A). While the parental line reached 1.3 × 107 cells at day 47, clone UKY-116.2F9 grew better, reaching 3.6 × 107 at day 40. This was a consistent result. In comparison, clones UKY-116.1D7, UKY-116.4H8, and UKY-116.6F5 were slow growers and did not reach comparable levels when examined as late as day 77. Interestingly, supernatants from confluent cultures of the more rapid grower UKY-116.2F9 did not increase the growth potential of UKY-116 parental or selected clones, while supernatants from confluent cultures of the slow grower UKY-116.4H8 did not inhibit cell growth (data not shown).
Figure 3A also shows the growth of UKY-29 parental line and selected clones isolated from 1 cell/well cultures. Growth of the parental line was greater than that observed for 1650; however, it took almost thrice the time to reach that level (day 16 vs. day 42). When analyzed 42 days after the establishment of cell culture, the parental UKY-29 line had reached 5 × 107 cells, with clones UKY-29.15D8 and UKY-29.12B10 reaching 4 × 107 cells. Clones UKY-29.11F8, UKY-29.11G8, and UKY-29.14G10 grew to a lesser extent, with UKY-29.14E5 growing the least when assessed at day 42 of cell culture. These results were confirmed by thymidine incorporation analysis (data not shown). Higher levels of tritiated thymidine incorporation were correlated with increased growth as determined by manual cell counts.
To further examine the heterogeneity of these clones and parental lines, the ability of UKY-29 and derived clones to secrete the immunosuppressive cytokines IL-10 and TGF-β1 was studied. The cells were harvested, adjusted to 1 million/mL of culture medium, and seeded in 24-well culture plates. At 24 hours, the medium was removed and analyzed for the presence of cytokines. The parental UKY-29 TC line secreted a small amount of IL-10 (around 10 pg/million cells, horizontal axis) but close to 2 ng of TGF-β (Fig. 3B). The clones examined secreted variable levels of these cytokines in 24 hours. Levels of TGF-β ranged from 900 pg/mL (clone 12B10) to 2900 pg/mL (clone 11G8) in 24 hours. IL-10 levels, however, ranged from 1 pg/mL for clone 11G8 to 44 pg/mL for clone 11F8. There was no correlation between growth and cytokine release (compare Fig. 3A and B). While clone 12B10 grew similar to the parental line, it secreted roughly half the amount of TGF-β compared to the parental line. Conversely, clones such as UKY-29.11F8, UKY-29.14E5, and UKY-29.14G10, which grew much less than the parental line, secreted varying levels of the immunosuppressive cytokine. As mentioned above, little IL-10 was released by any of the clones, the most significant amount released being 44 pg/million cells, by clone 11F8.
TAA analysis
The expression of TAA was assessed by flow cytometry and/or RT-PCR (see Table 1). 1650-TC was included as a control in these studies, and antigen expression observed was similar to that previously reported. 31 1650-TC expressed Her-2neu and CEA as determined by cell surface phenotyping and verified by RT-PCR. It also expressed vimentin by cell surface phenotyping but was negative for cytokeratin. 1650-TC possessed mRNA for WT-1, Mage 2 and 3, Cage, and survivin. The UKY-116 parental line expressed Her-2neu as determined by both flow cytometry and RT-PCR. It was also positive for WT-1 and Cage as determined by RT-PCR. While majority of the clones derived from UKY-116 were similar to the parental line, UKY-116.2F9 differed notably, as it did not express Her-2neu on the cell surface and was confirmed negative by RT-PCR. Thus, heterogeneity was observed for at least one of the clones with regard to expression of TAA.
Cell surface phenotype performed as described in the Materials and Methods section.
RT-PCR performed as described in the Materials and Methods section.
Cell lines analyzed included the control adenocarcinoma 1650-TC, and three TC lines and clones derived in the current authors' laboratory.
FACS, fluorescence activated cell sorter; PCR, polymerase chain reaction; ND, not determined; N/A, not available.
Analysis of UKY-29 and its clones showed that the parental line was similar to 1650-TC as it expressed Her-2neu, CEA, and vimentin while also expressing cytokeratin as determined by cell surface phenotyping. RT-PCR confirmed the expression of Her-2neu and CEA and also the presence of mRNA for WT-1, Mage 2 and 3, and survivin. There were no differences in antigen expression for any of the UKY-29-derived clones analyzed. Finally, UKY-349 and its clone UKY-349.4B3 were identical in terms of antigen expression. These cell lines expressed Her-2neu and CEA and possessed mRNA for Mage 1, survivin, and Cage.
MHC class I typing was done using PCR. 1650-TC (HLA-A2—; B15—; C7—), UKY-116 (HLA-A2—; B44—; C2—), and UKY-29 (HLA-A2—; B15, 55; C3—) were HLA-A2 positive. Differences were observed in the other A locus as well as in B and C loci, which confirmed the distinct lineages of these cell lines. UKY-349 (HLA-A1—; B7—; C3—) did not express HLA-A2 but was positive for HLA-A1. All of the cell lines were negative for MHC class II in the absence of IFN-γ up-regulation, as determined by cell surface phenotyping analysis.
Use of UKY-116 in MLTC
Earlier studies showed that 1650-TC and UKY-29 could stimulate allogeneic MLTC using CD80 (data not shown). 30 In the present study, following transfection with CD80 and selection in 0.4 mg/mL of G418, the UKY-116.7 clone was used in an autologous setting at a responder-to-stimulator ratio of 10:1 using lymphocytes derived from fresh UKY-116 tumor. Following an initial stimulation, the lymphocytes that grew in the MLTC were restimulated every 10 days with UKY-116.7. At day 35 of cell culture, the lymphocytes were tested for their ability to kill the UKY-116 tumor target. UKY-116 was significantly lysed, while the control cell lines K562 and Daudi and the NSCLC cell lines 1355, 1650 (shared HLA-A2), 838, and 1155 (non-A2) were not lysed to an appreciable degree (Figure 4A). The T cells were also tested for their ability to secrete IFN-γ when exposed to UKY-116 or UKY-116.2F9. As shown in Figure 4B, there were a number of spots indicating IFN-γ-secreting T cells in the wells stimulated with UKY-116 or UKY-116.2F9, suggesting that at least one of the antigens recognized was shared between the parental line and autologous clone.
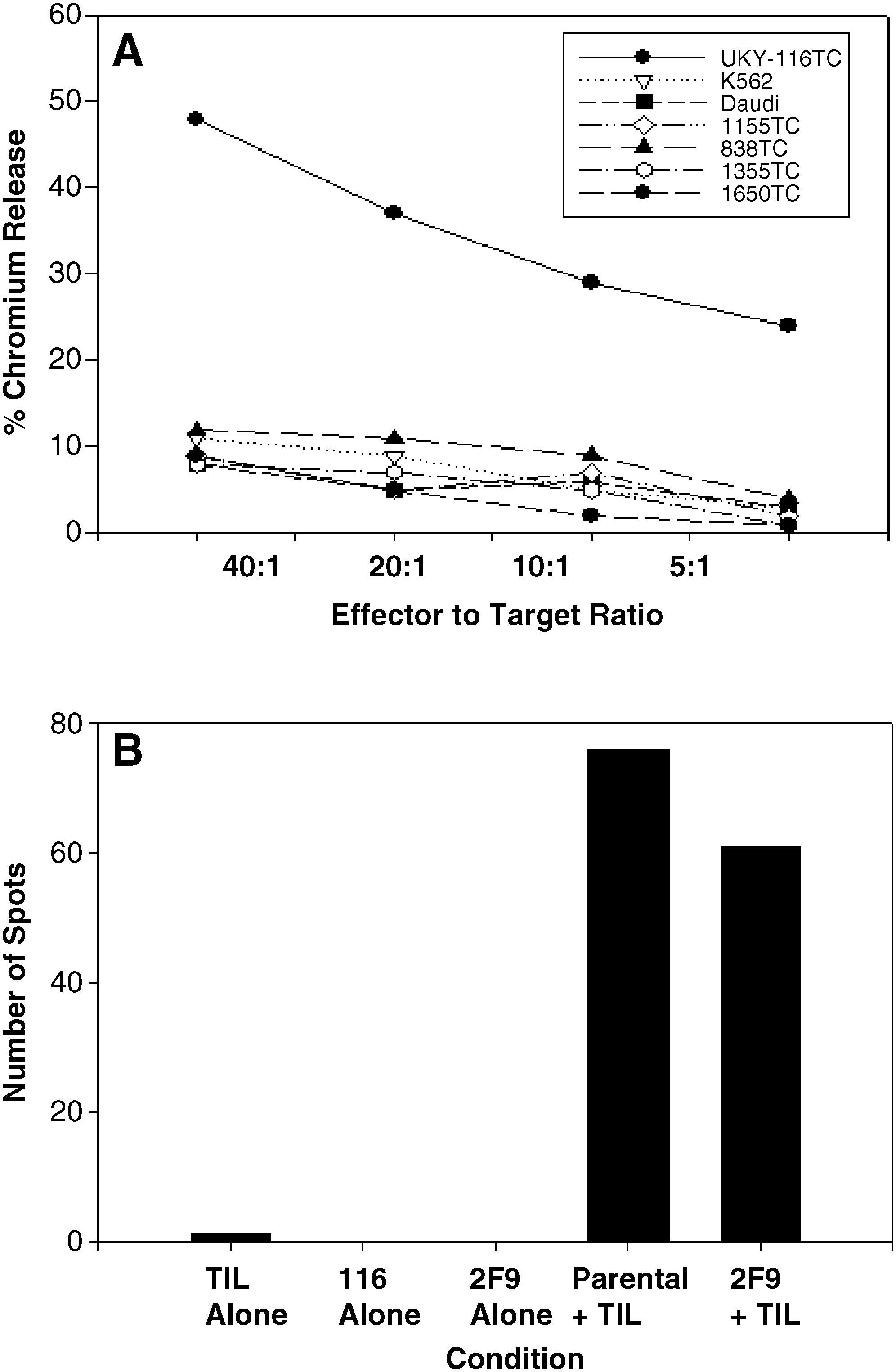
The ability of UKY-116 to stimulate specific T cells studied using MLTC. Functional reactivity was analyzed by cytotoxicity and ELISPOT. (
Discussion
In this study, attempts were made to develop TC lines in NSCLC using two medium formulations and different cell culture techniques. Biopsies were obtained from CHTN or the University of Kentucky hospital. Few reports other than the seminal report of Oie et al. 21 detail successful cell line generation in NSCLC. Many of the NSCLC lines currently available were derived in that unique study. This contrasts with melanoma, whose TCs adapt to in vitro conditions more consistently, almost 60% of the time. 27 As shown in the current report, NSCLC is difficult to establish in long-term stable cell cultures. Only three successful cases were obtained using 62 separate NSCLC samples (4.8% success rate). These stable cell lines could be routinely passaged in vitro, survived cryopreservation and thaw, and maintained NSCLC antigen expression. TCs were evident early in culture and began as a distinct colony of cells, presumably arising from single cell clones. The tumor colonies enlarged, eventually fused with neighboring colonies, and could be repeatedly passaged. Long-term adenocarcinoma and squamous cancer cell lines were obtained from either solid tumor biopsies or pleural fluid samples. Sugaya et al., 24 in a very comprehensive analysis, reported a success rate of 2.6% (15 cell lines in 570 attempts). In their study, success was high with tissue derived from more advanced-stage patients. More recently, Tirino et al. 32 reported the generation of three long-term lines (3 of 89 attempts, 3%) following the isolation and culture of CD133+ TCs, which represent cancer stem cells.
In the present report, the most prevalent outcome was the failure to generate a long-term TC line. In some situations no cells grew from the single cell suspensions. In others, the cultures were overtaken by fibroblasts at early stages or were overgrown by bacteria, mold, and/or yeast. TC growth was observed in only 9 of the 62 cases attempted, and only two-thirds (6 of 9) of these cultures were short-term lines, thereby providing limited chances for study. There was a sharp delineation between a short-term TC line and a long-term, established line, which could be maintained following extensive in vitro passaging. Figure 1 shows a representative culture with TC colonies that went through a period of proliferation followed by cessation in TC growth, thus allowing the culture to be overtaken by the less stringent fibroblast populations. In the 6 short-term lines obtained, initial TC growth was confirmed by cytochemical staining for cell surface antigens such as Her-2neu, CEA, WT1, and/or survivin. In all cases, tumor antigen-positive cells were mixed with varying numbers of normal fibroblasts. Short-term TC lines provide only limited opportunities for the study of cancer cell biology. Nonetheless, they have been exploited as autologous TC vaccines 7,33 to yield promising but limited results. Immunologic responses might be improved using homogeneous, more established tumor lines containing higher percentage of TCs as opposed to varying levels of normal cells as reported.
The reasons for the difficulty in generating long-term TC lines in NSCLC are not clear. Stringent cell culture requirements appear to be necessary but, at the moment, the identity of growth factors that can be consistently applied is not known. While TCs often appear in early cultures, their maintenance past a critical stage of culture is halted due to the lack of proper growth signals. Hybrid medium formulations such as ACL-4 that contain human epidermal growth factor, selenium, and transferrin have been successful but cannot be universally applied. These agents are adequate to a certain degree in initiating TC growth but are not substantive enough to maintain long-term TC passages.
In this study, cell growth and tumor antigen expression were examined in both parental lines and clones to evaluate TC heterogeneity. While heterogeneity in growth kinetics, and antigen expression and/or function could be attributed to infection with Mycoplasma species, 34 –36 all lines were confirmed clean by PCR analysis. This ensured that the differences in growth reported accurately reflected the growth potential of the TC parental line or clone. Differences were observed between the growth of parental lines and individual clones as shown in Figure 3A and B. When early-passage UKY-116 (passage 20) and late-passage UKY-29 (passage 102) parental lines and representative clones were compared, 3 of 5 UKY-116 and 5 of 7 UKY-29-derived clones grew to <50% of that observed for the parental line. While 2 of the UKY-116 clones grew better, 2 of the UKY-29-derived clones had growth approximating that of the parental line. The heterogeneity observed, while not entirely consistent, suggested that regardless of the passage interval cloned, parental lines contained clones with both high and low growth potential. It remains unclear whether growth potential was affected by a contact mediated mechanism or by one influenced by paracrine and autocrine cytokine release. Limited studies using Transwells (Costar) have shown that poor growing clones do not influence rapid growers and vice versa (data not shown). In addition, bulk cytokine studies (Fig. 3B) showed that while the growth potential of UKY-29 and associated clones was different, so was the expression and secretion of certain cytokines normally secreted by NSCLC. For example, there was no correlation between levels of TGF-β and IL-10 and the ability of UKY-29 clones to grow.
Additional evidence supporting TC line heterogeneity was obtained by cell surface phenotyping and/or RT-PCR analysis for the expression of NSCLC TAA (Table 1). These data, along with MHC class I antigen expression, validated that the parental lines were genetically unique even though some TAA were shared on a population basis. For instance, TAA expression of 1650-TC and UKY-29 showed similarities, but 1650-TC expressed Cage while UKY-29 expressed cytokeratin. UKY-349 differed from all other lines in this study in that it did not express WT-1. UKY-116 differed from both 1650-TC and UKY-29 in that it was derived from a patient with squamous cell carcinoma. Typical of squamous cell cancer, UKY-116 and its clones did not express CEA. Interestingly, most clones expressed similar patterns of antigens as did the parental lines. The only consistent difference was noted with UKY-116 and an associated clone, UKY-116.2F9. This clone lacked Her-2neu expression in repeated analysis and grew consistently better than the parental line. Antigenically, however, it was shown to be similar to the parental line in immunologic assay. In studies designed to test the relevancy of MHC and TAA expression, UKY-116 was transfected with the gene for CD80 and used as a stimulator in MLTC. 30,37,38 Results showed that UKY-116 could stimulate both autologous cytotoxic and cytokine-producing lymphocytes. Bulk populations of UKY-116 tumor-reactive lymphocytes recognized UKY-116.2F9 equally well, indicating that at least one TAA was shared between the two cell lines. This is consistent with previous results using transfected TCs as targets for lymphocyte generation.
Conclusions
The current study demonstrated that it is difficult to generate long-term stable TC lines in NSCLC. Though a large-size sample was used (62 biopsies), the majority of attempts resulted in no growth or the growth of fibroblasts. From the 62 samples studied, only 3 long-term cell lines could be derived. These were cloned by limiting dilution, and some heterogeneity was noted in terms of function. Improved techniques to generate TC lines in histologies other than melanoma will be crucial to the development of immunotherapies such as vaccines in the study of NSCLC. Based on earlier studies of BCG and allogeneic TCs used as vaccines in lung cancer, 39 an allogeneic NSCLC line is currently being used as a source of apoptotic bodies in NSCLC. 25,26 To date, 47 patients have been immunized and, regardless of the stage of disease or prior therapy, 67% of the immunized patients generated a lymphocyte response to the vaccine. More repeatable techniques to develop stable cell lines for individual patients could increase the relevancy and potential for vaccine use. Inclusion of autologous antigens in addition to allogeneic antigens in a vaccine will likely lead to more meaningful laboratory and clinical responses.
Footnotes
Acknowledgments
The author would like to acknowledge the technical support of Ms. Katherine Mitchell, Ms. Giovi Hidalgo, and Ms. Sara Perkins in generating and characterizing the cell lines described in this study. The author would also like to thank the Kentucky Lung Cancer Research Fund (KLCR) as well as the Kentucky Center for Science and Technology, Kentucky Science and Engineering Foundation (KSEF) (KSEF-950-RDE-008) for generously supporting the studies. Sincere gratitude is extended to the NSCLC patients, as well as all patients with cancer who volunteer for clinical studies and provide tissue for laboratory analysis. Without these tissue samples these studies cannot be performed. Finally, the efforts of Ms. Bonnie Sigafus, who passed away in 2008, are acknowledged. Ms. Sigafus helped to maintain the KLCR and was excited to see the accomplishments of laboratory research funded by the organization. Studies such as this are meant to memorialize her long-lasting efforts.
Disclosure Statement
No competing financial interests exist.