
Abstract
The β-catenin gene is a critical component of Wnt signaling pathway. Aberrant activation of Wnt/β-catenin signaling and subsequent upregulation of β-catenin is related to enhancing cell proliferation and developing colon polyps and colon cancer. In the present study, the effect of β-catenin knockdown on the growth and survival of the human colon cancer cell line HT-29 was investigated in vitro. The effect of knockdown of β-catenin on cell proliferation was investigated by MTT assay and colony formation. The cell cycle distribution was investigated by flow cytometry. Apoptosis was measured by nuclear staining and flow cytometry. The change of β-catenin and related proteins were determined by western blotting and immunofluorescence. The results showed that small interfering RNA directed against β-catenin markedly inhibited the expression and nuclear translocation of β-catenin and decreased the expression of known target genes such as cyclin D1 and c-myc; HT-29 cell proliferation was inhibited as indicated by growth reduction, cell cycle arrest in G0/G1 phase, and induction of apoptosis; and the inhibition of cell growth may be associated with switching off cyclin D1 and c-myc expression by small interfering RNA targeted against β-catenin in colon cancer HT-29 cells.
Introduction
Colon adenocarcinoma is one of the most devastating of human malignancies and the second leading cause of cancer-related deaths in developed countries. 1 Numerous studies suggest that activation of the Wnt/β-catenin signaling pathway plays an important role in human tumorigenesis. 2 –4
In the absence of a Wnt signal, β-catenin is mostly associated with the plasma membrane, where in conjunction with E-cadherin and α-catenin it promotes cellular adhesion. Cytosolic β-catenin is normally bound to axin and the adenomatous polyposis coli (APC) protein, phosphorylated at the N-terminal Ser/Thr residues by casein kinase Iα and glycogen synthase kinase 3, and then degraded by the ubiquitination-proteasome system. In response to a Wnt signal, β-catenin accumulates in the cytoplasm and is translocated to the nucleus, where it binds to proteins of the T-cell factor (Tcf)/lymphoid enhancer factor family and regulates the expression of genes involved in proliferation, invasiveness, and angiogenesis. 5,6 Therefore, the stabilization of β-catenin and its translocation into nucleus are the key steps in Wnt pathway activation, which may serve as potential target for colon cancer therapy.
RNA interference (RNAi) is a post-transcriptional gene-silencing mechanism, which was first discovered by Fire and coworkers in 1998. 7 The most extensively characterized system to date is targeted mRNA degradation mediated by small interfering RNAs (siRNAs). siRNAs are 21–25-nucleotide double-stranded RNAs (dsRNAs) with 5′ phosphate groups and 3′ dinucleotide overhangs produced from dsRNAs through the action of the RNase III enzyme Dicer. This process may have evolved as a cellular defense in response to viral infection or the activity of transposable DNA elements. siRNAs can also target mRNAs for destruction. 8 –10 RNAi has been exploited as an experimental tool for silencing gene expression and analyzing the functions of mammalian genes.
In the present study, siRNA against β-catenin was used to investigate the effect on expression of downstream targeted genes, c-myc and cyclin D1, and the effect of knocking-out β-catenin on the growth and survival of HT-29 human colon cancer cells.
Materials and Methods
Main reagents
The human colon carcinoma cell line HT-29 was obtained from the Cancer Institute of the Chinese Academy of Medical Science. The Cycle Test PLUS DNA reagent kit was bought from Becton-Dickinson. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and dimethyl sulfoxide were purchased from Sigma Chemicals. β-Catenin (sc-7199), mouse monoclonal antibody for c-myc (sc-7480), FITC-conjugated fluorescent secondary antibody, β-catenin siRNA (sc-29209), and control siRNA (sc-37007) were purchased from Santa Cruz Biotechnology. Goat anti-rabbit (w3960) and anti-mouse (w3950) secondary antibodies were purchased from Promega. Rabbit polyclonal antibody for cyclin D1/Bcl-1(SP4) (RM-9104) was obtained from Neomarkers.
Cell culture
Human colon carcinoma HT-29 cells were maintained in RPMI 1640 (Gibco) in 75 cm2 flasks at 37°C in a 5% CO2 atmosphere at constant humidity. The medium was supplemented with 10% (v/v) heat-inactivated fetal bovine serum (Gibco), 2 mM
RNA interference
HT-29 cells were cultured in complete medium and transfected at 60% confluence. β-Catenin siRNA and control siRNA were used for transfection by using siRNA transfection medium and siRNA transfection reagent (Santa Cruz Biotechnology) according to the manufacturer's instructions. Briefly, the siRNA duplex solution (for each transfection, 6 μL of siRNA duplex added to 94 μL siRNA transfection medium) was directly added to the diluted transfection reagent (for each transfection, 6 μL of siRNA transfection reagent added to 94 μL siRNA transfection medium), then mixed gently, and incubated for 30 minutes at room temperature. For each transfection, 0.8 mL siRNA transfection medium was added to transfection reagent mixture. After mixing, the mixture was overlaid onto the cells and the cells were washed with 2 mL of siRNA transfection medium. Then the cells were incubated for 6 hours in a CO2 incubator. HT-29 cells were washed once with phosphate-buffered saline (PBS) and cultured with complete medium. After 24–72 hours, the cultured cells were assayed using the appropriate protocol.
Western blotting
After siRNA treatment, HT-29 cells were washed twice with PBS and detached with PBS solution containing 0.25% trypsin and 0.02% EDTA. Western blotting was conducted as previously described. 11
Transfection and luciferase assay
Cells were transfected using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. Briefly, HT-29 cells were seeded into six-well plates at 50% confluence and transfected with β-catenin siRNA. At 12 hours post-transfection, the cells were transfected with TOPflash or FOPflash reporter and pRL-CMV Renilla control reporter (Promega) to control for transfection efficiency. After 48 hours, luciferase assays were carried out using a Dual Luciferase Assay kit (Promega). Luciferase activity was normalized by Renilla luciferase activity.
Immunofluorescence detection of β-catenin
HT-29 cells were seeded onto glass cover slips in six-well plates (Nunc) and incubated overnight. After β-catenin siRNA transfection for 48 hours, the slips were washed with PBS and fixed in methanol for 4 minutes. The intracellular location of β-catenin was determined as previously described. 12 Briefly, after repeated washings with PBS, the cover slips were incubated for 30 minutes at room temperature with rabbit anti–β-catenin antibody and then incubated with a fluorescein isothiocyanate–conjugated secondary antibody. Nuclei were counterstained with 4′,6-diamidine-2′-phenylindole dihydrochloride (DAPI, 10 mL/mL; Roche). Slides were analyzed by confocal microscopy (TE2000U; Nikon) and digitally photographed.
Cell viability
The effect of transfection of β-catenin on cell viability was determined by MTT assay. Briefly, at 24, 48, and 72 hours after β-catenin siRNA transfection, 10% (vol/vol) MTT (5 mg/mL in PBS) was added to each well and incubated at 37°C for 4 hours. The medium was carefully removed and 150 μL of dimethyl sulfoxide was added to each well. The plates were shaken for 10 minutes and the absorbance at 490 nm was measured in an Elx800 Universal microplate reader (Bio-Tek Instruments). The OD value for the MTT assay was calculated to determine the viabilities of control cells, control siRNA cells, and siRNA-treated cells.
Colony formation
At 24 hours post-transfection, HT-29 cells were seeded at 400 cells/well in 24-well plates and allowed to attach for 36 hours. After incubation, the medium was renewed and the plates were incubated for 12 days under the same culture conditions. The cells were then fixed with methanol and stained with Giemsa. Colonies containing >50 cells originating from single cells were counted under an inverted microscope (Olympus CK40).
Flow cytometry
Cell cycle analysis was carried out by the method as previously described with some modifications. 13 Briefly, after treatment with control or β-catenin siRNA for 48 hours, HT-29 cells were harvested, washed three times with cool PBS, fixed with 70% cool ethanol for 2 hours, and stained with propidium iodide (Cycle TEST PLUS DNA Reagent Kit). For each concentration, at least 2.5 × 104 cells were analyzed by FAC Sort flow cytometry (BD Biosciences). The proportions in G0/G1, S, and G2/M phases were estimated using ModFit LT analysis software.
Morphological observation of apoptosis
Morphological changes in HT-29 cells at 48 hours after β-catenin siRNA transfection were assessed by fluorescence microscopy. Changes in the nuclei were investigated by staining the cells with fluorescent DNA-binding dyes. The assay was conducted as previously described with some modifications. 14 Briefly, the cells were harvested and washed with PBS, and 20 μL of cell suspension was mixed with 1 μL DAPI (10 mg/mL). Nuclear morphology was observed by confocal microscopy (Olympus IX70).
Statistical analysis
Statistical analysis was performed using SPSS 13.0 software. All the results were expressed as mean ± standard deviation. Differences between the control and treated groups were evaluated by Student's t-test. A p-value of <0.05 was considered statistically significant.
Results
siRNA inhibits β-catenin expression and its nuclear localization in HT-29 cells
The western blotting results showed that there were no significant changes in the expression of total β-catenin proteins after transfection with control siRNA for 48 hours in HT-29 cells. siRNA transfection did not affect the level of β-catenin within 24 hours (Fig. 1). However, after 48 and 72 hours, the expression of β-catenin protein in HT-29 cells transfected with the gene-specific siRNA was 65% lower than that in cells transfected with the control siRNA (Fig. 1).
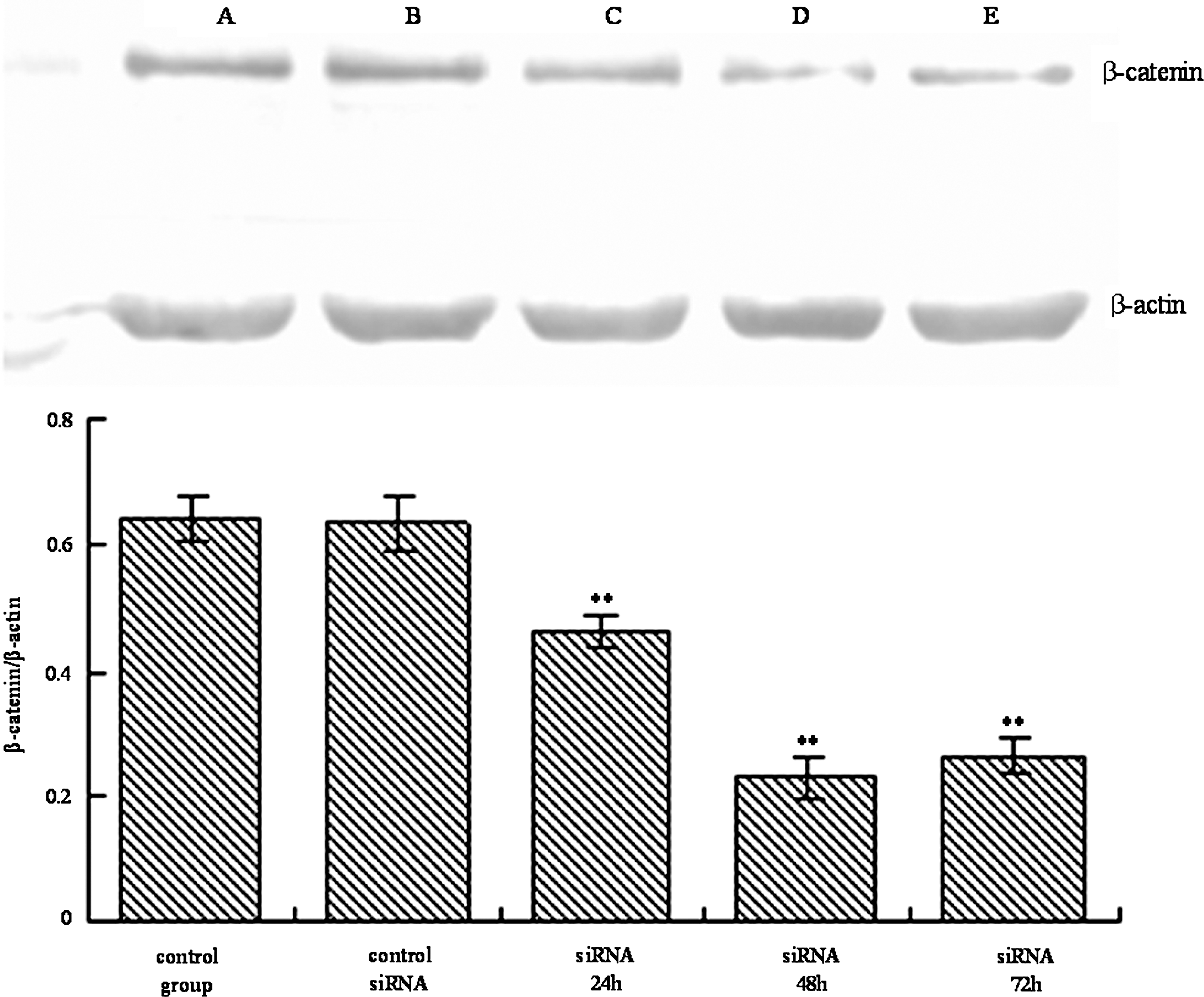
Effects of β-catenin siRNA on expression of β-catenin in HT-29 cells. Western blotting of HT-29 cells for β-catenin. The β-actin gene served as internal control.
To confirm any changes in its location in the nucleus and cytoplasm after β-catenin siRNA or control siRNA transfection, an immunofluorescence experiment was conducted using the anti–β-catenin antibody. The results showed strong nuclear and cytoplasmic staining of β-catenin in HT-29 cells (Fig. 2). No significant changes were observed in both nuclear and cytosolic β-catenin in HT-29 cells transfected with control siRNA for 24, 48, or 72 hours (data not shown). Treatment with specific β-catenin siRNA for 48 and 72 hours significantly reduced nuclear β-catenin but did not markedly affect the cytosolic β-catenin. The level of membrane-associated β-catenin increased in cells treated with β-catenin siRNA for the same time, and this may relate to cell adhesion. These results suggest that β-catenin siRNA transfection may potentially reduce the levels of β-catenin protein in HT-29 cell nuclei.
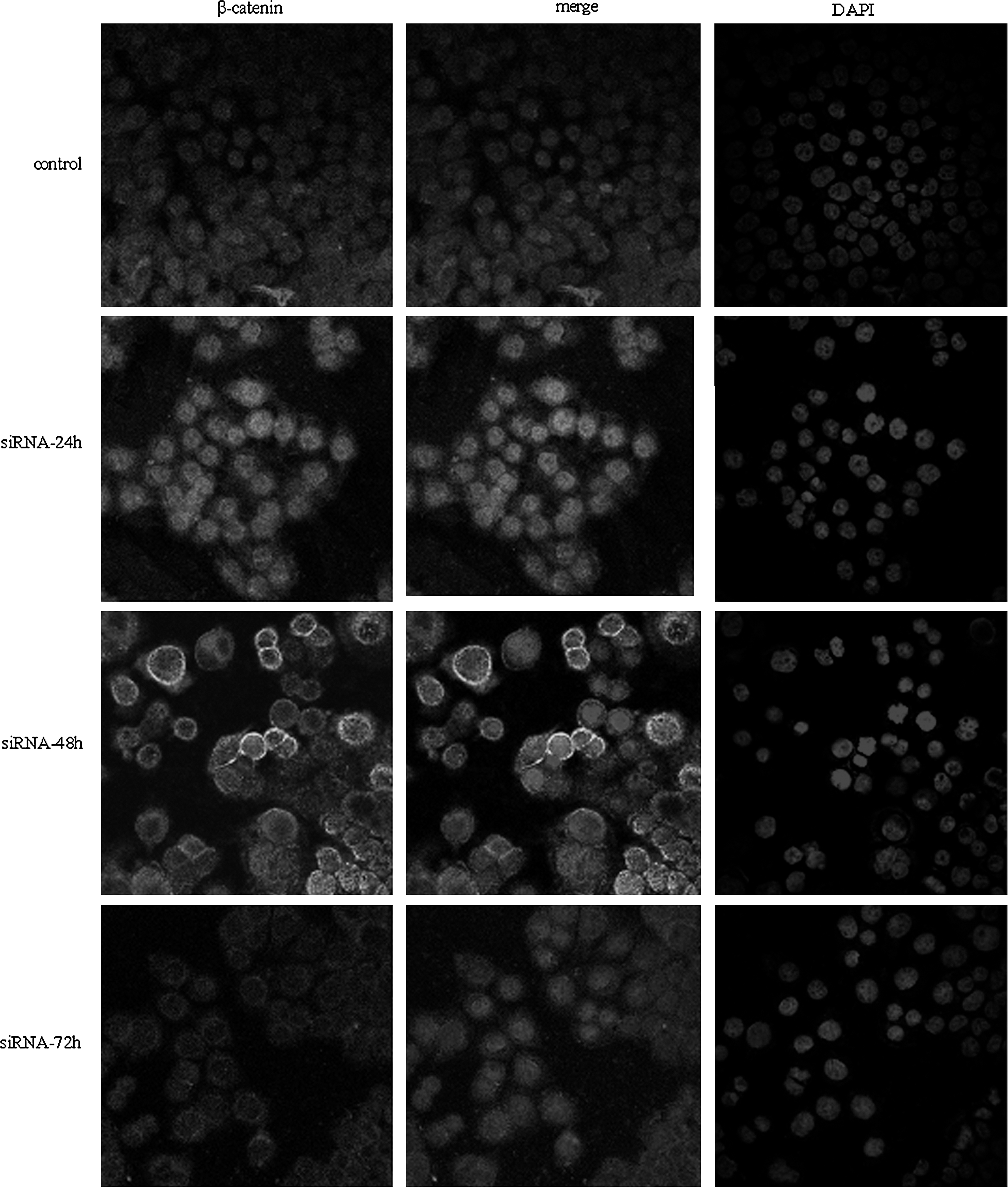
Effects of β-catenin siRNA on subcellular location of β-catenin in HT-29 cells. Immunofluorescence images are shown (original magnification, × 200).
Knockdown of β-catenin inhibits β-catenin/Tcf signaling in HT-29 cells
After 24 hours of transfection with β-catenin siRNA, HT-29 cells were transiently transfected with plasmids encoding luciferase driven by a promoter containing either Tcf-binding sites (TOPflash) or mutated Tcf-binding sites (FOPflash), together with Renilla control plasmid. The transfection with β-catenin siRNA resulted in a dramatic decrease (60%–75%) in TOPflash activity (Fig. 3a). In contrast, FOPflash activity remained unchanged after siRNA treatment. The results suggested that functional binding of β-catenin/Tcf may be important for TOPflash. The results presented in Figure 3A show that β-catenin siRNA specifically suppresses the transcriptional activity of β-catenin/Tcf signaling.
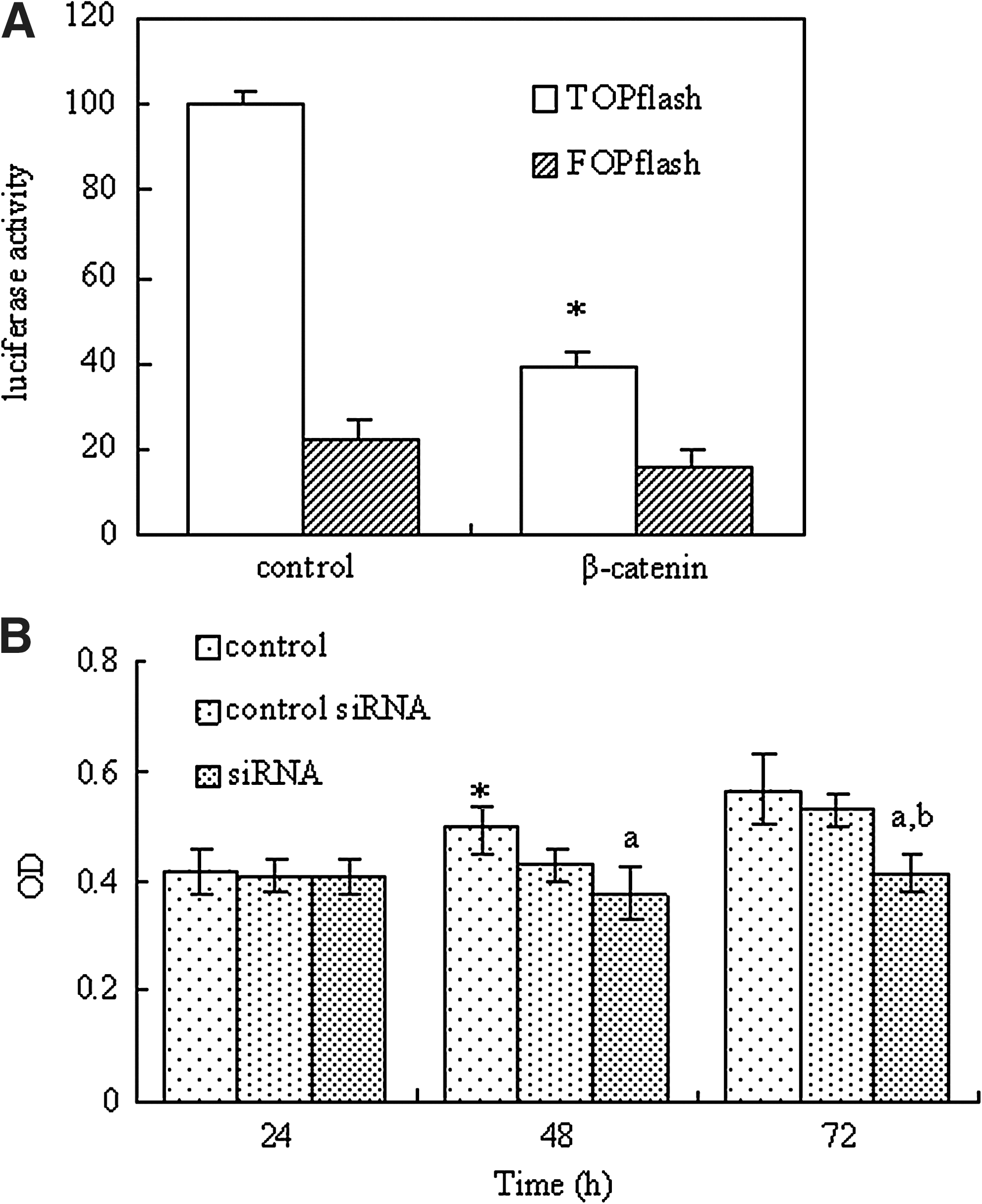
Knockdown of β-catenin inhibits HT-29 cell growth
The effects of transfection with siRNA on the viability of HT-29 cells are shown in Figure 3B. After transfection with control siRNA for 24, 48, or 72 hours, cell viability did not differ from the control group. The statistical analysis showed that the viability of HT-29 cells transfected with β-catenin siRNA for 48 and 72 hours was significantly lower than that of the control group. In particular, only 67.4% cells survived after 72 hours transfection. The differences were more clearly evident under light microscopy (Fig. 4). Comparing with the control group, after β-catenin siRNA transfection for 72 hours, most of the cells showed shrinkage, rounding, and refractile morphology; fragmentation was even observed in some cells, in contrast to the control group.
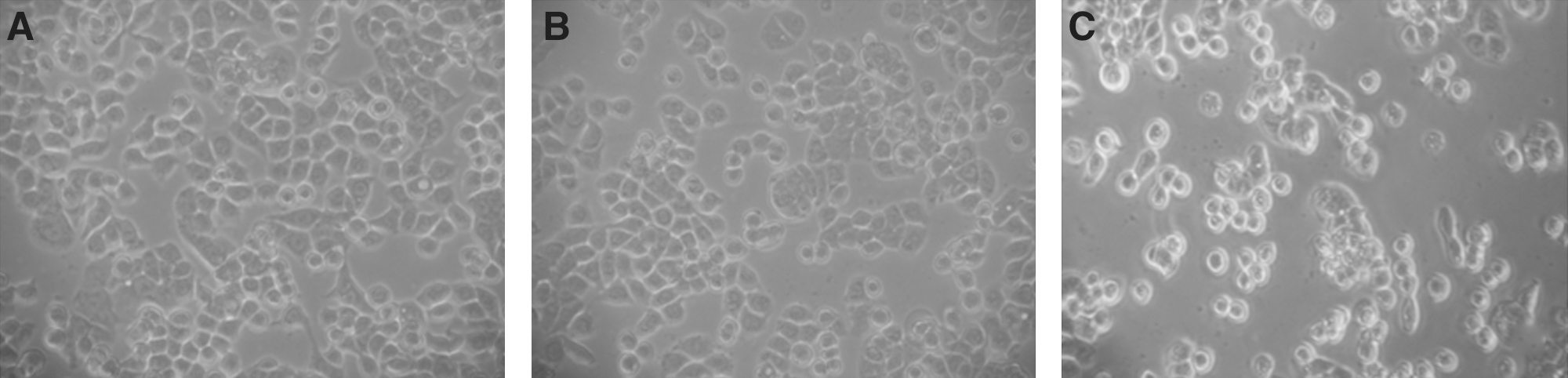
The morphological changes in HT-29 cells treated for 48 hours (original magnification, × 100).
Knockdown of β-catenin inhibits HT-29 cell colony formation
The effects of siRNA targeting β-catenin on the ability of HT-29 cells to form colonies were examined. The colony-forming rates for the β-catenin siRNA-transfected cells, control RNAi, and control cells were 3.2%, 13.7%, and 15.10%, respectively. The results suggest that β-catenin gene silencing decreases the ability of colon cancer HT-29 cells to form colonies (p < 0.05).
Knockdown of β-catenin induces cell cycle arrest and increases apoptosis in HT-29 cells
To further confirm the growth inhibition induced by β-catenin siRNA on HT-29 cells, the cellular DNA content was measured by flow cytometry after transfection with β-catenin siRNA, control siRNA, and vehicle for 48 hours. The effects of β-catenin RNAi on cell cycle progression are shown in Table 1. Percentage of control cells in S phase was 24.53% of total cells. β-Catenin siRNA treatment had a lower proportion of S-phase cells up to 13.70% and the proportion of G0/G1-phase cells was increased from 57.10% to 69.96%. No differences were observed between the control siRNA group and control group (p > 0.05). The results indicate that β-catenin siRNA transfection results in an accumulation in G0/G1 phase and a reduction in S phase in HT-29 colon cancer cells.
p < 0.05 compared with the control group.
siRNA, small interfering RNA.
Moreover, to investigate whether β-catenin siRNA–mediated growth inhibition is associated with apoptosis, treated and untreated HT-29 cells were analyzed by flow cytometry. As shown in Table 1, the apoptosis rates of HT-29 cells were around 15% after transfection with β-catenin siRNA, which was much higher than that recorded for the control group. The number of apoptotic cells transfected with control siRNA was nearly the same as that of control group. In addition, the DAPI staining was used to investigate the morphological changes of the cell nuclei and the results are presented in Figure 5. As shown in Figure 5, control cells showed homogeneous staining of the nucleus, but, after treatment with β-catenin siRNA for 48 hours, apoptotic cells had irregularly stained nuclei indicative of chromatin condensation and nuclear fragmentation. No significant change was observed in HT-29 cells treated with control siRNA. These data reveal that β-catenin siRNA treatment results in apoptosis of HT-29 cells.
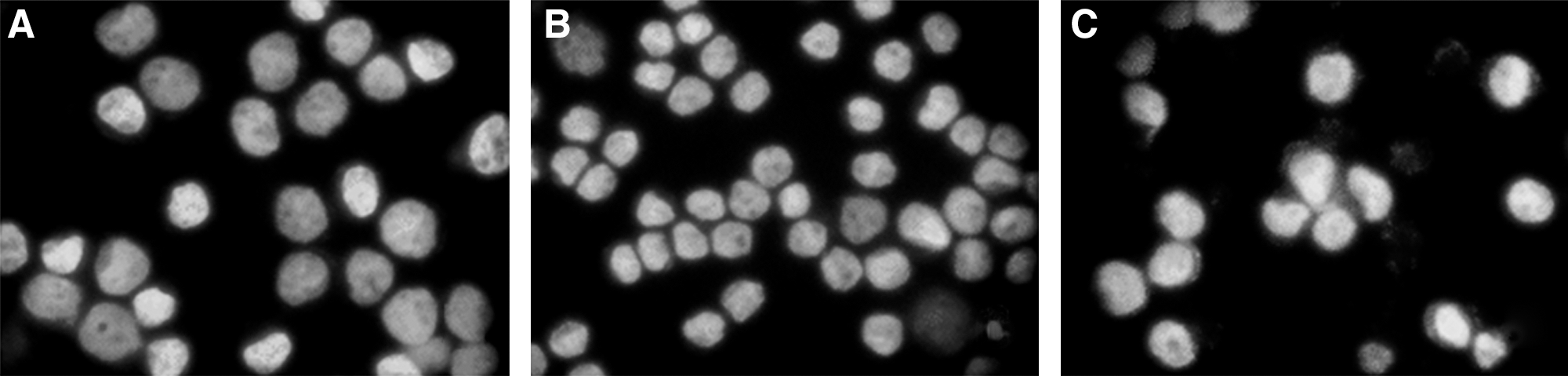
The morphological changes indicating apoptosis in HT-29 cells treated for 48 hours (original magnification, × 200).
β-Catenin siRNA reduces expression of known target genes
Further, the expression of two cellular genes known to be regulated by β-catenin was examined. As shown in Figure 6A and B, no significant changes were found in the expression levels of c-myc and cyclin D1 in HT-29 cells transfected with control siRNA for 24 hours, whereas transfection with β-catenin siRNA for 72 hours resulted in significantly lower endogenous expression of c-myc and cyclin D1 than those of control group (p < 0.05). These results indicate that siRNA specifically decreases the expression of β-catenin–dependent genes.
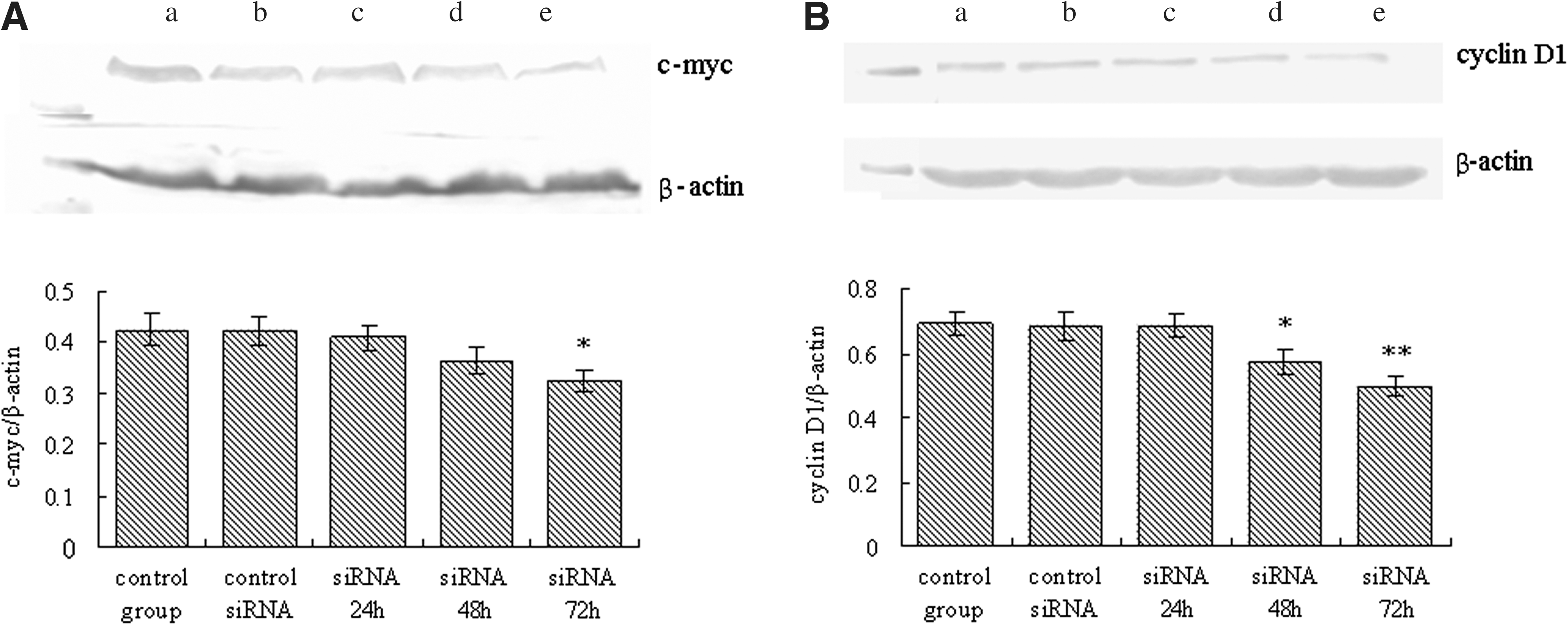
Western blotting of HT-29 cells for c-myc and cyclin D1 at different times post-transfection.
Discussion
The Wnt signaling pathway is deregulated in over 90% of human colorectal cancers. 15 In most colon cancers, mutation in either the APC or β-catenin gene leads to increased level of β-catenin. 2,3,5,16,17 The increased level of β-catenin leads to increased expression of β-catenin/TCF-regulated genes such as c-myc and cyclin D1 18 –20 and promotes cellular proliferation. 21 –23 HT-29 cells with mutations in the APC gene have increased β-catenin levels 17,24 ; this cell line has an activated Wnt pathway. 25,26
RNAi is an important method for analyzing gene function and identifying new drug targets, using dsRNA to knock down or silence specific genes. β-Catenin has attracted much attention as a gene therapy target for carcinomas, including colon cancer. It has been successfully downregulated by RNAi in some previous studies; for instance, Verma and coworkers synthesized siRNA and interfered with the β-catenin gene in the colon cancer cell lines SW480, HCT116, and colo205. 27,28 Another previous study reported that β-catenin RNAi inhibits the expression of β-catenin protein in human hepatoma HepG2 and Hep3B cells. 29 However, the downregulation of β-catenin expression by RNAi in the colon cancer HT-29 cell line has not been reported previously. In the present study, synthetic siRNA duplexes targeted against human β-catenin mRNA were transiently transfected into HT-29 cells. After 48 and 96 hours of transfection, the siRNA significantly suppressed the levels and changed the nuclear localization of β-catenin protein, in contrast to cells transfected with the negative control siRNA and blank controls. The data of the present study also suggest that β-catenin siRNA may significantly reduce the transcriptional activity of β-catenin/Tcf signaling and downregulate β-catenin–dependent gene expression.
β-Catenin is one of the most important proteins in the cadherin–catenin complex, which plays a critical role in intercellular adhesion. β-Catenin binds directly to the cytoplasmic tail of E-cadherin in a mutually exclusive manner. The E-cadherin–catenin complex plays a crucial role in epithelial cell–cell adhesion and in the maintenance of tissue architecture. Perturbation in the expression or function of this complex results in the loss of intercellular adhesion, with possible consequent cell transformation and tumor progression. Human cancers possess both irreversible and reversible mechanisms for inactivation of the E-cadherin–catenin complex, which lead to invasion and metastases. 30 β-Catenin siRNA increased the level of membrane-associated β-catenin in HT-29 cells and then it might restrain the adhesion ability of the cells.
Further studies were conducted on the basis of the efficiency of β-catenin gene silencing. Both the MTT and colony formation assays revealed that β-catenin siRNA significantly decreased tumor cell viability and proliferation. The results of the present study show a correlation between the downregulation of β-catenin expression and inhibition of human colon cell growth. These results support earlier reports about tumor cell lines with mutations in the APC or β-catenin gene. 27,29 The flow cytometry results indicated that the inhibition of cell growth by β-catenin siRNA may be due to cell cycle arrest in G0/G1. A similar result has been reported in colo205 cells. 28 These observations also suggest that G1 arrest following siRNA treatment might be involved in its efficacy of inhibiting colon cancer cell growth. Apoptosis is a cellular suicide process and it is also important for eliminating unwanted or damaged cells and has been recognized as one of the major modes of cell death in cancer therapy. In the present study, the flow cytometry and morphological data showed that treatment with β-catenin siRNA increased HT-29 cell apoptosis. However, this result contrasts with Verma's report, which declared that the decrease in cell proliferation did not depend on increased apoptosis in either SW489 or HCT116 cells. 27 This disparity may be due to the varieties of cell lines. The data of the present study indicate that the specific β-catenin siRNA inhibits cell proliferation and the process is accompanied by cell cycle arrest in G0/G1 and the induction of apoptosis.
Although previous studies, in which β-catenin was functionally inactivated through antisense or siRNA strategies, 27,31 demonstrated the important role of β-catenin in colon cancer development, the mechanism remains unknown. Also, in the present study, a correlation between the expression levels of c-myc/cyclin D1 proteins and β-catenin was observed. The proto-oncogene c-myc encodes a transcription factor, c-Myc, which is of great importance in controlling cell growth and vitality. The quantity of c-Myc is carefully controlled by many mechanisms. 32 Overexpression of c-myc was up to 50% in all human cancers 33,34 and in almost one-third of breast and colon carcinomas. 35,36 c-Myc has been a difficult protein to target because it is a nuclear transcription factor and lacks specific enzyme activity. Currently, no drugs that target c-Myc have been identified for treating human cancers. The c-Myc oncoprotein is strongly induced during the G0 to S-phase transition, and activation of c-myc expression is sufficient to drive quiescent cells into S phase. 37 These increases in cyclin D1 in some but not all colon cancers may result from the heterogeneity of β-catenin expression in tumors and from the fact that β-catenin is not increased in many colon cancer cell lines with mutant APC. 38 Cyclin D1 is a G1-phase regulatory protein that promotes physiological cell proliferation downstream of mitogens and other extracellular stimuli. 39 –41 Constitutive expression of cyclin D1 is likely to contribute to malignant transformation and decreases the requirement for mitogens in the transition through the G1 restriction point. Overexpression of these two genes may stimulate the cells to overcome the cell cycle checkpoints and enhance proliferation. 42 The data of the present study show that β-catenin levels may control c-myc and cyclin D1 transcription in colon cancer HT-29 cells. These results indicate that the siRNA targeted against β-catenin blocks the expression of the target genes cyclin D1 and c-myc. G1-phase arrest and induction of apoptosis may depend on turning off cyclin D1 and c-myc expression in HT-29 cells transfected with β-catenin siRNA.
Conclusions
The results reveal that knock-down of the β-catenin gene decreases the expression and nuclear translocation of β-catenin protein in colon cancer HT-29 cells and suppresses the expression of the β-catenin–dependent genes cyclin D1 and c-myc. Further, the inhibition of cell growth may be associated with switching off cyclin D1 and c-myc expression by the siRNA targeted against β-catenin. The results of the present study suggest that the β-catenin gene may be a useful target for anticancer treatment.
Footnotes
Acknowledgments
The authors thank Prof. Jia-Ren Liu from the Agricultural Research Station, Virginia State University, for her helpful suggestions and assistance. This project was supported by the National Natural Science Foundation of China (No. 30471444) and Development Program for Outstanding Young Teachers in Harbin Institute of Technology (HITQNJS.2009.055).
Disclosure Statement
No conflicts of interest exist.