
Abstract
The influence of specific blocking of the Delta-like ligand 4 (DLL4)/Notch signal transduction pathway on the biological behavior of human umbilical vein endothelial cells (HUVECs) has been studied. Recombinant adeno-associated virus (rAAV) vectors expressing an active small interfering RNA (siRNA) (vector 6) targeting the DLL4 (rAAV-DLL4-short hairpin RNA [shRNA]) was used to infect HUVECs. The same cell line infected with empty plasmid (rAAV-EGFP) was used as a control. Stable transfection and expression of DLL4-mRNA in HUVECs were determined by semiquantitative RT–polymerase chain reaction (PCR). Protein expression of DLL4 was examined by western blotting. The distribution of cells in cell cycle was assessed by flow cytometry and cell growth was analyzed by MTT assay. HUVECs were seeded on type I collagen and cultured in a three-dimensional culture system to allow for tubule-like structure (TLS) formation. Compared with negative controls, semiquantitative RT-PCR and western blot analysis showed that the expression of DLL4 mRNA and protein was downregulated in stably transfected cells (p = 0.024, p = 0.033). HUVEC growth and proliferation were stimulated following infection with rAAV vectors containing active siRNA against DLL4, whereas infection with empty plasmid had no specific effect. The proliferation index of rAAV-DLL4-shRNA-infected HUVECs was 39.90% ± 2.19% compared with 25.63% ± 4.54% (p = 0.036) for control-treated cells. TLS formation was significantly induced in cells expressing the rAAV vector; the average length of TLS was greater than the control group (p = 0.028). Altogether, the data suggest that inhibiting the DLL4/Notch signal transduction pathway stimulated proliferation of HUVECs, thereby facilitating angiogenesis.
Introduction
Tumor angiogenesis is closely related to biological characteristics such as tumor growth, metastasis, and recurrence. 1 Therefore, research directions that focus on antitumor angiogenesis and the inhibition of effective microcirculation in tumor tissues are important topics for the comprehensive treatment of tumors. Recent studies have found that the Delta-like ligand 4 (DLL4)/Notch signaling pathway plays an important regulatory role in tumor angiogenesis. Specifically, tumor growth was effectively suppressed by inhibiting DLL4 overexpression, although the microvessel density within the tumor tissue increased. 2 As such, further study on the mechanism of the DLL4/Notch signaling pathway in regulating tumor angiogenesis is warranted and may lead to important theoretical and practical significance for exploring DLL4 as a new target in antiangiogenic therapy against cancer. We used packaged recombinant adeno-associated virus (rAAV-DLL4-short hairpin RNA [shRNA]) expressing shRNA specifically targeting the DLL4 gene to infect human umbilical vein endothelial cells (HUVECs). We observed HUVEC growth and proliferation and its ability to form small tube-like structures (tubule-like structures [TLS]) in vitro. Probing into the possible mechanism by which the DLL4/Notch signaling pathway regulates angiogenesis may lead to new therapeutic targets and is therefore an ideal candidate in the investigation into antitumor angiogenesis.
Materials and Methods
Materials
Adeno-associated virus expressing RNAi fragment specifically targeting DLL4 (rAAV-DLL4-shRNA; with a titer of 5 × 1015 v.g./L) was prepared and stored in our laboratory. Control rAAV-EGFP adeno-associated virus (with a titer of 5 × 1015 v.g./L) was a gift from the Beijing Benyuan Zhengyang Corporation. The HUVEC line was established in our laboratory. RT–polymerase chain reaction (PCR) kits were purchased from TaKaRa Company. M199 medium and standard fetal calf serum were purchased from America Hyclone. Matrigel was purchased from BD Company. Endothelial cell growth additives were purchased from America Sigma Company. Rabbit anti-human DLL4 mAb was obtained from Abcam, and horseradish peroxidase-conjugated secondary antibodies were from America Santa Cruz Corporation. Western blot reagents were purchased from Gemmy Bio-Engineering Co., Ltd., and tetrazolium was from the Sino-American Biotechnology Company. Coulter flow cytometer was developed by the America Coulter Company. The microplate reader used for these studies is an HTS-7000 Bio-analyzer. Other reagents used were analytically pure products, which were imported or domestically produced.
Methods
Cell culture and virus infection
A HUVEC line was developed from HUVECs (provided by Dr. Qiong Wu, Gynecology and Obstetrics, The People's Liberation Army 307 Hospital). The cells that had been cultured in M199 complete media and incubated at 37°C and 5% CO2 for <10 total generations were used during the logarithmic growth phase for the experiments. 3 A monolayer of cells was washed three times with M199 prior to the addition of M199 medium containing rAAV-EGFP with corresponding multiplicity of infection values of 4 × l04, 1 × 105, and 5 × 105 per well. Cells were incubated for 1 hour at 37°C, then an additional 2 mL of M199 was added to each well, and cells were further cultured for 72 hours under the same conditions. rAAV-DLL4-shRNA with the best MOI was chosen to transfect HUVECs according to the literature 4 and the transfecting media was discarded after 24 hours. G418-resistant cells were screened and collected in M199 culture medium containing 300 mg/L of G418. The cells transfected with the recombinant virus vector rAAV-DLL4-shRNA or with the empty virus vector rAAV-EGFP have been abbreviated as HUVECs(+) and HUVECs(−), respectively, whereas nontransfected control cells are called HUVECs.
The variation in DLL4-mRNA expression in HUVECs(+) detected by semiquantitative RT-PCR
HUVECs(−), HUVECs(+), and control HUVECs were collected and divided into four groups. Total RNA was extracted and quantified based on Tripure instructions and RNA was adjusted to a concentration of 1 g/L. Exactly 2 μL of total RNA was used for reverse transcription as previously described.
5
The products of reverse transcription were stored at −20°C in the following PCR reaction mix: 2 μL reverse transcription product (template), 1 μL for two pairs of each primer (choosing housekeeping gene GAPDH as an internal reference), the upstream primer 5′ AATTCAACGGCACAGTCAAGGC 3′, and the downstream primer 5′ GGATGCAGGGATGATGTTCTGG 3′, which yielded a 467-bp amplified fragment. DLL4 upstream primer sequence was 5′ TGCTGCTGGTGGCACTTT 3′, the downstream primer sequence was 5′ CTTGTGAGGGTGCTGGTT 3′, and the amplified fragment was 624 bp. The reaction system used was as follows: 2.5 μL of 10 × buffer, 2 μL of 25 mmol/L MgCl2, 2 μL of 10 mmol/L dNTP mix, and 0.5 μL Taq enzyme; total volume was 25 μL with deionized water. Reaction conditions used were same as that described in the corresponding literature.
5
Ten (10) microliters of PCR product was used for 2% agarose gel electrophoresis, Gel-Doc 2000 gel imaging analysis system was used to scan DNA bands, and Image-glub 4.0 analysis software was used to measure the optical density of each gene band. The ratio of DLL4 intensity to GAPDH internal control strength for each sample was calculated so as to reflect differences in DLL4 expression and calculate
Assaying of DLL4 protein expression
The cells up to 90% growth density were washed twice in 10 mmol/L phosphate-buffered saline (PBS) followed by aspiration. Then, 200 μL of l × SDS loading buffer was added to fully lyse the cells, and lysates were then transferred to new 1.5-mL Eppendorf tubes using a cell eraser. The lysate was heated at 100°C for 10 minutes; cells were then sonicated, centrifuged at 1000 rpm/min for 10 minutes at room temperature, and the supernatant was transferred into a new 1.5-mL tube. The protease inhibitor PMSF was added at a concentration of l0 μL/mL. The protein concentration was measured with BioRad SmartSpec by spectrophotometer and the total protein of each group was diluted to the same concentration before storage at −20°C. Forty (40) micrograms of total protein from HUVECs(+), HUVECs(−), and HUVEC lysates were collected and mixed with an equal volume of sample buffer and denatured at 100°C for 5 minutes. After cooling, the samples were separated by 10% SDS-PAGE electrophoresis gel and then transferred to a nitrocellulose membrane. Membranes were blocked for 2 hours at room temperature in PBST containing l% skimmed milk. Primary antibody (1:100) was added and incubated at room temperature overnight. Membranes were washed in PBST and then incubated with a secondary antibody (1:2000) for an additional 2 hours at room temperature. The results were recorded by a gel imaging system after DAB chromogenic reaction. With β-actin as an internal reference, the change in DLL4 protein expression between each group was detected. The gray value of each band was measured by Mias image analysis system and data analysis was performed using the ratio of the integral optical density value of DLL4 to the β-actin band from the same tube to estimate the level of DLL4 protein expression.
Detection of the inhibition rate of cells by MTT colorimetry
HUVECs(+), HUVECs(−), and HUVECs at logarithmic growth phase were plated at a density of 5.0 × 103 per well into 96-well plates, 24 (3 × 8) wells per cell type. The cells were cultured at 37°C, 50 mL/L CO2, and saturated humidity overnight, and after 24 hours the culture medium was replaced and a 5-day continuous observation was performed. Eight wells were examined at each time point by MTT assay. Briefly, 20 μL MTT solution (5 g/L) was added per well, followed by a 4-hour incubation at 37°C. The supernatant was discarded and the cells were subjected to gentle oscillations for 10 minutes. For each well, the absorption value (A) at 570 nm was measured using ELISA to determine cell proliferation.
Analysis of the influences on cell cycle distribution by flow cytometry
The logarithmic growth phase cells were digested with trypsin, passaged, and inoculated at a density of 1.0 × 108 per liter. Once adherent, the cells were cultured in M199 complete medium for 24 hours, then the medium was refreshed, and the cells were continuously cultured for 72 hours. The cells were then digested into a single-cell suspension by trypsin, washed twice with ice-cold PBS, and centrifuged at 1000 rpm for 5 minutes. After supernatant was discarded, precooled ethanol was slowly added (250 mL/L ethanol precooled at −20°C). The cells were maintained at 4°C overnight, and cell suspension was taken, washed with PBS, and centrifuged at 2000 rpm for 5 minutes. The supernatant was discarded, cells were stained with 1.0 mL PI for 30 minutes, a mesh filter with 300 wells was used to remove impurities in the mixture, the samples were determined at 488 nm excitation wavelength, and PI fluorescence at 620 nm was detected using band-pass filters, collecting more than 10,000 fluorescent signals for each sample. DNA index value of the cell cycle was analyzed by the software MultiCycle, and cell proliferation index was calculated in terms of the following formula:
Formation of TLS in vitro
TLSs were analyzed using Matrigel (BD Company). The stock solution was diluted 1:3 with serum-free M199 medium, and 50 μL of diluted Matrigel solution was then added to each well of an ice-cold 96-well plate. The plates were incubated and placed at 37°C in an incubator, and in 30 minutes, the wells were converted by a gelatinous layer. HUVECs(+), HUVECs(−), and HUVECs were inoculated (1 × 104 cells/group), 100 μL M199 complete medium was added into 21 wells, placed at 37°C, and cultured in 5% saturated CO2 incubator. Media were refreshed every 2 days, and digital images were recorded under an inverted phase-contrast microscope every 12 hours. The length of TLS of 5 pieces as 400 × digital images was measured by Image-Pro Plus image analysis system (calculated in mm/mm2 as indicated).
Statistical analysis
Experimental data were indicated as
Results
Results of semiquantitative RT-PCR
The gene fragments amplified using DLL4 and GAPDH primers were 624 and 467 bp, respectively, based on electrophoresis and EB staining, which corresponds to the designed fragments. The expression of DLL4-mRNA in HUVECs(+) was remarkably lower than that in HUVECs or HUVECs(−) (Fig. 1). The expression of DLL4-mRNA in HUVECs(+) was lower than that of HUVECs(−). The ratios of the intensity of DLL4 PCR products to internal control GAPDH in the control groups and experimental group were 0.972 ± 0.022, 0.948 ± 0.046, and 0.636 ± 0.082. Thus, the ratio of the experimental group to the control group was ∼65.4% (p = 0.024, n = 4).
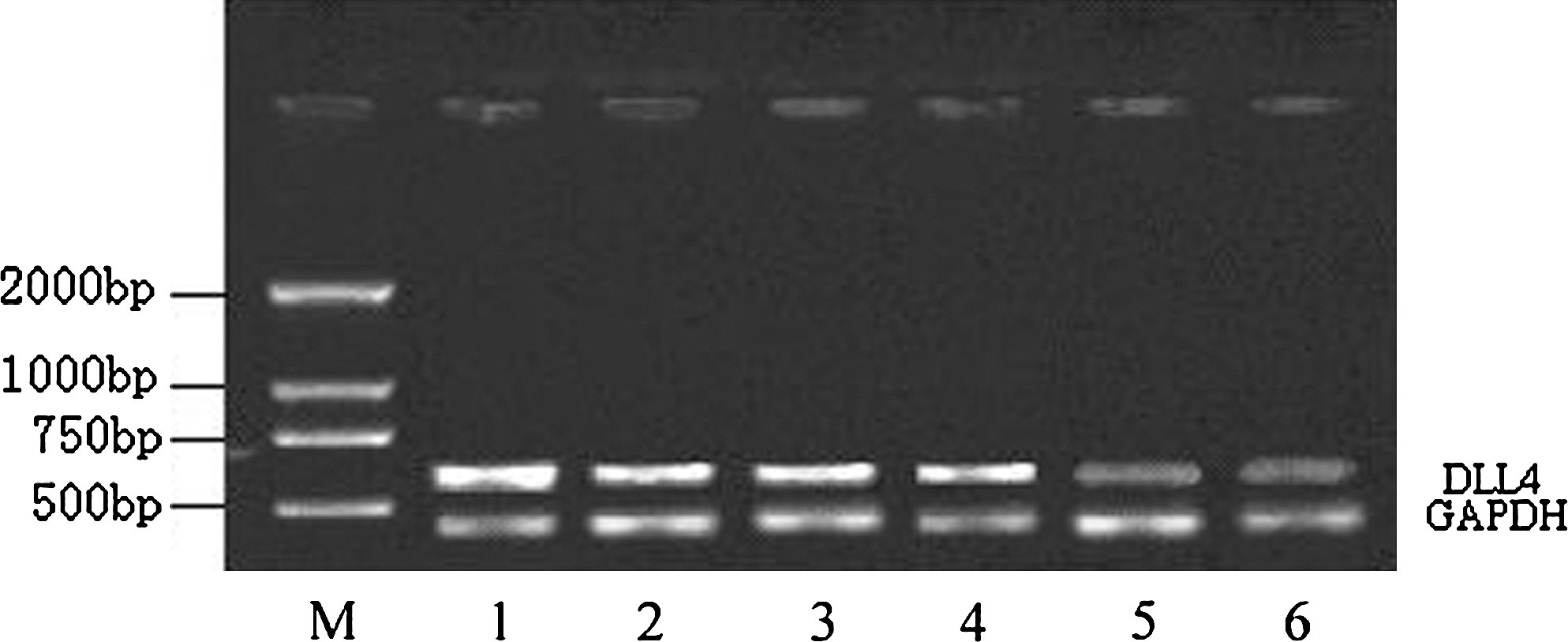
The variation in the expression of DLL4-mRNA in empty viral vector and recombinant virus vector particles before and after HUVEC transfection. Lanes 1 and 2: HUVECs in nontransfected group; lanes 3 and 4: HUVECs (−) group transfected by empty viral vector; lanes 5 and 6: HUVECs (+) group transfected by recombinant virus. DLL4, Delta-like ligand 4; HUVECs, human umbilical vein endothelial cells.
Detection of DLL4 protein expression in each group by western blot
Western blot analysis showed that DLL4 was differentially expressed in the various cell groups, using β-actin as the internal reference. For instance, blank control (HUVECs) and the empty viral vector-transfected group [HUVECs(−)] had relatively high expression of DLL4, whereas DLL4 expression was significantly lower in HUVECs(+) (values were 2.016 ± 0.048, 1.946 ± 0.066, and 0.632 ± 0.052, respectively; p = 0.033; Fig. 2).
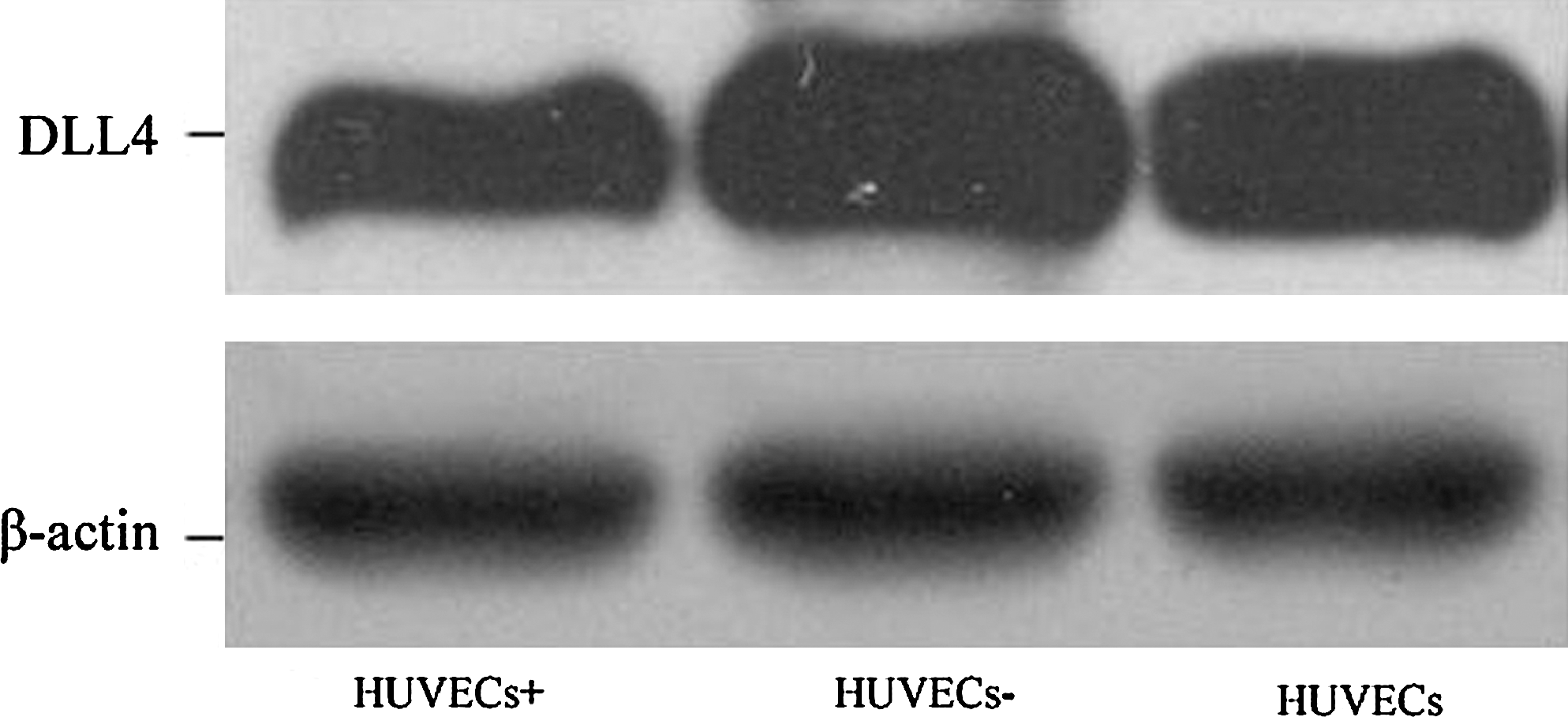
Changes in the protein expression of DLL4 in HUVECs after transfection by recombinant virus. DLL4, Delta-like ligand 4; HUVECs, human umbilical vein endothelial cells.
Determination of cell proliferation by MTT colorimetry
AAV-mediated recombinant virus vector (rAAV-DLL4-shRNA), expressing DLL4 shRNA, significantly promoted the growth of HUVECs, whereas empty virus vector played no role in promoting cell growth (p < 0.05; Fig. 3).
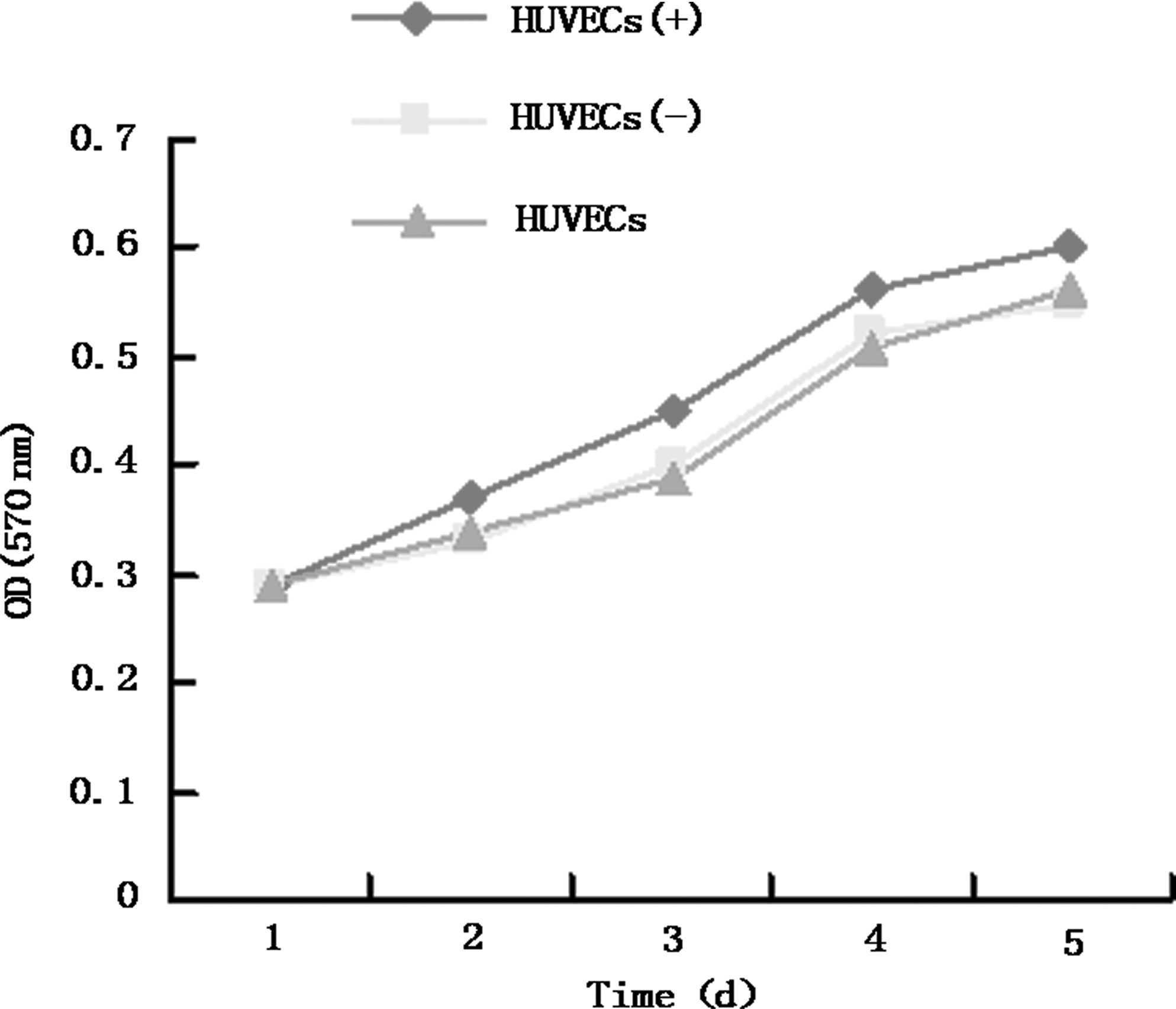
Changes in the cell growth curve after transfection by recombinant adeno-associated virus-Delta-like ligand 4-short hairpin RNA vectors.
The variation in cell cycle distribution measured by flow cytometry
The proportion of G0/G1 phase cells was decreased in HUVECs(+) lines, which were transfected with the recombinant virus vector (rAAV-DLL4-shRNA). On the other hand, the proportion of cells in S phase, as well as the ratio of G2/M cells, was increased and the cell proliferation index was remarkably higher. In contrast, the cell proliferation index of HUVECs(−), which were transfected with empty viral vector (rAAV-EGFP), had no apparent change (Table 1).
Compared with control group, a p = 0.064 and b p = 0.036. The values are shown to 1 decimal place.
HUVECs, human umbilical vein endothelial cells.
Effects of recombinant virus on the formation of three-dimensional TLS by HUVECs
Round or oval-shaped HUVECs were inoculated onto Matrigel cultures. During culture, these cells gradually attached to the surface of the Matrigel and stretched out, extending protrusions and interconnecting by the formation of TLS between adjacent protrusions. The length and the number of branch points of small tube-like structures in the HUVEC line, which was transfected with the recombinant virus vector (rAAV-DLL4-shRNA), were notably greater than that of the control group (p = 0.028; Fig. 4, Table 2).

(
Compared with control group, a p > 0.05 and b p < 0.05. The values are shown to 1 decimal place.
HUVECs, human umbilical vein endothelial cells.
Discussion
The angiogenesis dependence theory proposed by Folkman 1 in 1971 sheds profound light on the relationship between tumor growth and angiogenesis, which led to antitumor angiogenesis becoming an important strategy for comprehensive tumor therapy. Tumor angiogenesis is an extremely complex process that is regulated by factors that induce angiogenesis as well as angiogenesis inhibitors. The common ideal of antitumor angiogenesis involves antagonizing inducing factors or enhancing inhibitors to block tumor angiogenesis so as to control tumor growth. At present, these goals are accomplished mainly through blocking vascular endothelial growth factor (VEGF) and its main receptor, resulting in the suppression of tumor angiogenesis. 6 However, clinical practice has found that a variety of tumors have antagonistic effects on VEGF inhibitors, resulting in ineffective suppression of tumor growth and angiogenesis. 7 Therefore, additional mechanisms for tumor angiogenesis, differentiation, and maturation remain to be explored to increase the understanding of how an effective network of microcirculation is formed. 8
The Notch gene, discovered in 1919, 6 encodes a membrane protein receptor that exists as a heterodimer consisting of a 180-kDa peptide (p180, one peptide containing most of the extracellular domain) and a 120-kDa peptide (p120, another peptide containing the transmembrane region and the intracellular domain). 9 Notch is activated by binding to its ligands, also transmembrane proteins known as Delta and Serrate in Drosophila and Lag-2 in nematode. There are currently four Notch homologs in vertebrates, including Notchl-4. Delta and Serrate homologs in mammals are called DLL and Jagged, respectively. Notch ligands found in mammals consist of single-transmembrane glycoproteins bearing an extracellular structure with different numbers of EGF-like repeats as well as an N-terminus containing a cysteine-rich DSL motif (Delta Serrate-Iag 2), which is necessary for binding and activating Notch. These ligands include DLL1, DLL3, Jagged1, Jagged2, and DLL4. Serrate and Jagged also contain conserved cysteine regions that are lost in the DLL. The intracellular structure of Notch ligands include a short, diversified sequence with unknown function, possibly related to polymer formation. DLL4, cloned from the mouse-derived gene, shares a functional region with other DLL ligands and, like other DLL ligands, also shares many similarities in amino acid sequences between mice and humans. Previous studies found that DLL4 is highly expressed in vascular endothelial cells 10 and overexpressed in human bladder cancer, renal cell carcinoma, and gastric cancer as an endothelial cell-specific ligand. 2,11 The receptors, Notch1 and Notch4, are expressed in vascular endothelial cells. Binding to the endothelial cell-specific ligand, Delta-like 4 (DLL4), it promotes the differentiation and maturation of tumor angiogenesis and the formation of effective microcirculation pathways. 12 –14 Infecting HUVECs with viruses carrying adeno-associated virus-mediated DLL4-targeting RNAi showed that specifically blocking the DLL4/Notch signaling pathway notably encouraged the proliferation of HUVEC lines. Cell cycle analysis suggested that the proportion of G0/G1 phase cells was reduced, whereas the ratio of S phase to G2/M cells increased, indicating that the proliferation index of cells in the experimental group was significantly increased compared with that of the control group.
In 1980, Folkman and Haudenschild 15 found that endothelial cells can form vessel-like structures in a two-dimensional culture system. They observed that the branches had a tendency to form a three-dimensional vascular structure, which was fully consistent with the capillary bed in vivo under the light microscope and electron microscope, resulting in the first establishment of an in vitro model for angiogenesis. Further study detected that endothelial cells cultured in the appropriate extracellular matrix can spontaneously form small tube-like structures, also known as capillary-like structures (CLSs), which stretch and connect to each other to form CLSs and leading to the appearance of original lumen within and between cells. 16 The results indicated that specifically blocking the DLL4/Notch signaling pathway strikingly improved the capacity of a HUVEC line to form a microvascular network, which displayed significant differences when compared with the control group. However, whether the network of new vessels has normal function and structure requires further research.
Conclusions
With the development of biotechnology, virus vector-mediated RNAi has become a research hotspot. Conventional viral vectors mediating RNAi are the adenovirus and retrovirus vectors, of which AAV vector is currently the only vector that does not cause any pathological reaction in humans. 17 In this study, we successfully applied a prebuilt adeno-associated virus vector containing a DLL4-targeting RNA interference fragment to infect HUVECs and specifically block the DLL4/Notch signaling pathway. These studies showed that DLL4 had a negative regulatory effect on the growth and proliferation of HUVECs, as well as the cell's capacity to form TLS on collagen type I Matrigel, but the detailed mechanism remains to be further studied.
Footnotes
Acknowledgments
This study was supported by a grant from the Natural Science Foundation of China (No. 30872468). The authors thank Yun-San Zhao, Ming-Zu Sun, Yang Li, and Hai-Wei Xu for their support with laboratory work.
Disclosure Statement
The authors declare that they have no conflicts of interest.