
Abstract
Sesamol (SM), a nutritional phenolic antioxidant compound present in sesame seeds, protected pBR 322 DNA from gamma radiation–induced damages. SM prevented gamma radiation–induced degradation of covalently closed circular form of plasmid DNA in a concentration-dependent manner. Also SM protected cellular DNA of mouse blood leukocytes exposed to 4 Gy gamma radiation, ex vivo, as revealed by the data from alkaline comet assay studies. SM (5 mM) showed a faster time-dependant decrease of the radiation-induced DNA damage in mouse blood leukocytes following postirradiation incubation ex vivo, which could be attributed to enhanced DNA repair. SM protected the biomembranes from radiation-induced lipid peroxidation. Thus, SM could act as a radioprotector for the biomembranes and cellular DNA against the deleterious effects of ionizing radiation.
Introduction
Ionizing radiation inflicts deleterious effects to living organisms through the generation of reactive oxygen species, which damages vital cellular targets such as DNA and membrane. 1 The DNA damages include strand breaks, base damage, sugar damage, and crosslinks of inter- and intrastrand types. 2,3 Many natural and synthetic compounds have been found to have considerable radioprotective ability. 4 The radioprotectors can elicit their action by various mechanisms such as inhibiting the formation of reactive free radicals, detoxification of radiation induced species, target stabilization, and enhancing the repair and recovery process. 5
Sesame (Sesamum indicum) is an important oilseed crop with nutritional and medicinal values and its seeds contain lignan compounds—sesamin and sesamolin. Sesamol (SM) (Fig. 1), a phenolic antioxidant compound, is a derivative of sesamolin and is generated on roasting sesame seeds. 6 –9 It exhibits various beneficial biological effects 10 such as chemopreventive, antimutagenic, and antihepatotoxic activities 11 –13 and induction of growth arrest and apoptosis in cancer and cardiovascular cells. 14 The radioprotective effects of SM on gamma radiation–induced DNA damage and lipid peroxidation have been reported in cultured human lymphocytes. SM pretreatment decreased radiation-induced micronuclei, chromosomal aberration, and oxidative stress in human lymphocytes exposed to gamma radiation ex vivo. 15 SM was reported to have radioprotective action in doses above 25 mg/kg body weight (bw) in mice. Pretreatment of mice with SM (50 mg/kg bw) before exposure to 15 Gy gamma radiation significantly reduced dead, inflammatory, mitotic, and goblet cells in irradiated jejunum and also increased crypt cells, thus protecting the gastrointestinal system from radiation-induced damages. 16 In the present study, SM was evaluated for its efficiency as a radioprotective agent against gamma radiation–induced cellular DNA and membrane damage and also its effect on cellular DNA repair under ex vivo conditions.
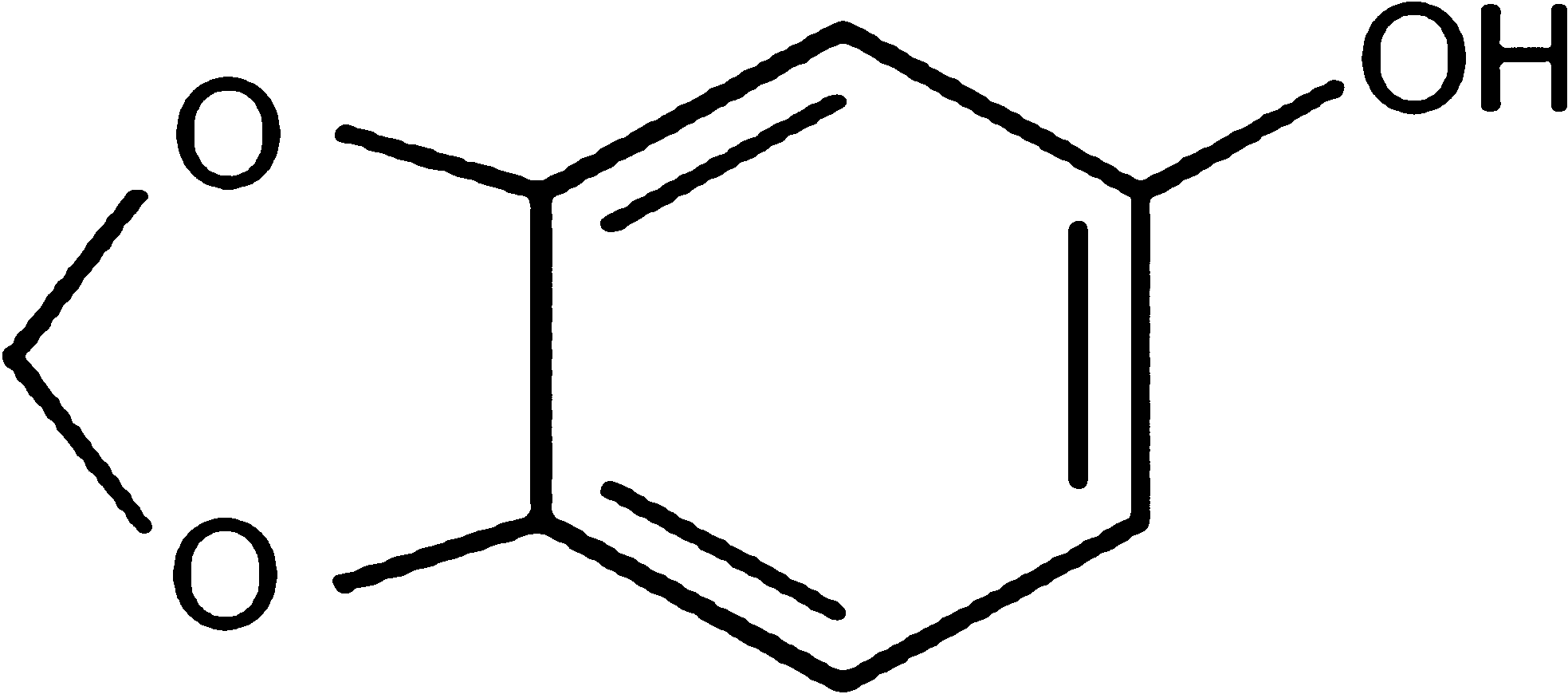
The chemical structure of sesamol.
Materials and Methods
Chemicals
SM (3,4-methylenedioxyphenol) was obtained from Spectrochem. Na2-ethylene diamine tetra-acetic acid and thiobarbituric acid (TBA) were purchased from Sisco Research Laboratories. Plasmid pBR322 DNA was obtained from Bangalore Genei, Dulbecco's modified Eagle's media (DMEM) from Gibco, and fetal bovine serum (FBS) from PAN Biotech GmbH. Ethidium bromide, high-melting-point agarose, and low-melting-point agarose were procured from Sigma-Aldrich. All other chemicals were of analytical grade and procured from reputed Indian manufacturers.
Animals
Swiss albino mice (male, n = 3) weighing 22–25 g were obtained from the Small Animal Breeding Section, Kerala Agriculture University, Mannuthy, Thrissur, India. They were maintained under standard conditions of temperature and humidity in the Centre's Animal House Facility. The animals were provided with standard mouse chow (Sai Durga Feeds and Foods) and water ad libitum. All animal experiments were carried out with prior approval of the Institutional Animal Ethics Committee and were conducted strictly adhering to the guidelines of Committee for the Purpose of Control and Supervision of Experiments on Animals constituted by the Animal Welfare Division of Government of India.
Exposure to gamma radiation
Irradiation was carried out using a 60Co-Theratron Phoenix teletherapy unit (Atomic Energy Ltd.), at a dose rate of 1.88 Gy/minute.
Protection of plasmid pBR322 DNA by SM against different doses of gamma radiation (0–20 Gy)
The plasmid pBR 322 DNA (100 ng) in phosphate buffer (0.1 M, pH 7.4) was exposed to various doses of gamma radiation (0–20 Gy) on ice in the presence and absence of SM (1 and 5 mM). After irradiation, the DNA samples were electrophoresed in 0.8% agarose gel containing 100 ng/mL ethidium bromide at 55 V for 2 hours and the DNA damage was analyzed by Digital Gel Documentation and Analysis Software (Biotech R&D Laboratories). 17 From the amount of the supercoiled form of plasmid DNA remaining after the radiation exposure, the dose-modifying factor (DMF) was calculated. DMF is defined as the ratio of the radiation doses to bring down the percentage of covalently closed circular (ccc) DNA to the same extent in the presence and absence of the radioprotector, that is, SM.
Protection against gamma radiation–induced DNA damage in peripheral blood leukocytes of mouse by SM, ex vivo
Blood from Swiss albino mice by heart puncture was collected into sterile heparinized vials and was exposed to 4 Gy gamma radiation in the presence and absence of SM (1 and 5 mM). The extent of DNA damages in the blood samples were measured as single-strand breaks using alkaline single-cell gel electrophoresis (Comet assay). 18,19 After electrophoresis the slides were dried and subjected to silver staining. 20 The comets were visualized using compound light microscope, images were captured, and a minimum of 50 comets per slide, in triplicates for a group, were analyzed using the software “CASP,” which gives % DNA in tail, tail length, tail moment (TM), and olive TM (OTM) directly. The parameter TM is the product of tail length and % DNA in tail, and OTM is the product of the distance between the center of the head and the center of the tail and % DNA in tail. 19 –21 Results are given as mean ± standard deviation. 17,19
Enhancement of cellular DNA repair in mouse blood leukocytes by SM
Mouse peripheral blood was collected and exposed to 4 Gy gamma radiation and comet assay was performed immediately after exposure (for 0 minute). To one of the groups, SM was added up to a final concentration of 5 mM when incubated in DMEM with 10% FBS at 37°C and to the other only medium was added, which served as the untreated irradiated control. Alkaline single-cell gel electrophoresis (Comet assay) was done at various time intervals during the incubation (15, 30, 45, 60, and 90 minutes) to monitor cellular DNA repair.
Ex vivo effect of SM on gamma radiation–induced membrane damage in mouse liver homogenate
Radiation-induced damage to membrane was assessed as peroxidation of membrane lipids according to the method of Buege and Aust. 22 Mouse liver homogenate (10%) was prepared in ice-cold phosphate-buffered saline (pH 7.4). The homogenate was centrifuged at 800 g for 5 minutes, and supernatant was collected and exposed to 25 Gy gamma radiation in the presence and absence of varying concentrations of SM. After irradiation the samples were analyzed for the presence of TBA reacting substance. The lipid peroxidation values are expressed as nanomoles of malondialdehyde (MDA) (using 1,1,3,3-tetraethoxy propane as standard) per mg homogenate protein. Protein was estimated by Lowry's method. 23
Statistical analysis
The results are presented as mean ± standard deviation of the studied group. Statistical analyses of the results were performed using analysis of variance with Tukey–Kramer multiple comparisons test, where each test group was compared with the respective control group.
Results
Protection of pBR 322 DNA from gamma radiation–induced damage
Exposure of plasmid DNA to gamma radiation resulted in induction of strand breaks, which converted the supercoiled form (ccc) of DNA to open circular (oc) form or linear forms. The disappearance of ccc form of plasmid DNA can be taken as an index of DNA damage induced by ionizing radiation. The reduction in the supercoiled form of plasmid DNA was directly related to the radiation dose as can be seen in lanes 1, 4, 7, 10, and 13 and protection by SM can be visualized in lanes 2, 3, 5, 6, 8, 9, 11, 12, 14, and 15 (Fig. 2). The data presented in Figure 3 revealed that the SM at a concentration of 1 mM protected the DNA partially from gamma radiation–induced DNA strand-breaks with a DMF of 1.3, whereas at 5 mM concentration of SM, protection of plasmid DNA was more effective with a DMF of 1.93. These results thus indicated that SM protected DNA in vitro effectively from gamma radiation–induced DNA strand-breaks.
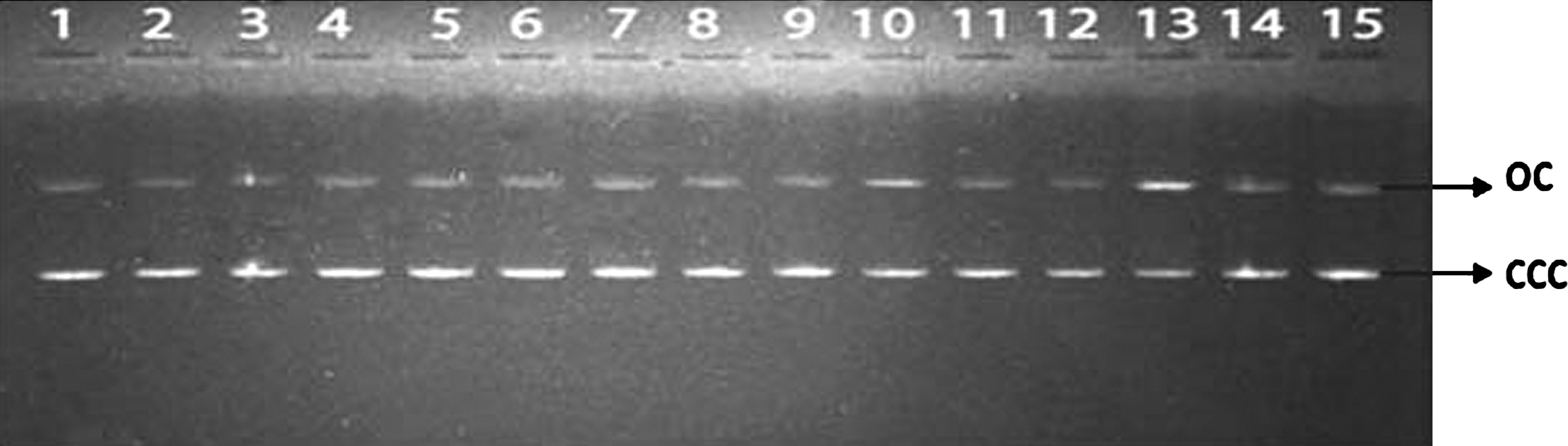
Effect of SM on plasmid DNA (pBR 322) damage induced by gamma irradiation. Lane 1: 0 Gy; lane 2: SM (1 mM) + 0 Gy; lane 3: SM (5 mM) + 0 Gy; lane 4: 4 Gy; lane 5: SM (1 mM) + 4 Gy; lane 6: SM (5 mM) + 4 Gy; lane 7: 8 Gy; lane 8: SM (1 mM) + 8 Gy; lane 9: SM (5 mM) + 8 Gy; lane 10: 12 Gy; lane 11: SM (1 mM) + 12 Gy; lane 12: SM (5 mM) + 12 Gy; lane 13: 20 Gy; lane 14: SM (1 mM) + 20 Gy; lane 15: SM (5 mM) + 20 Gy. SM, sesamol; oc, open circular DNA; ccc, covalently closed circular DNA; DMF, dose-modifying factor.
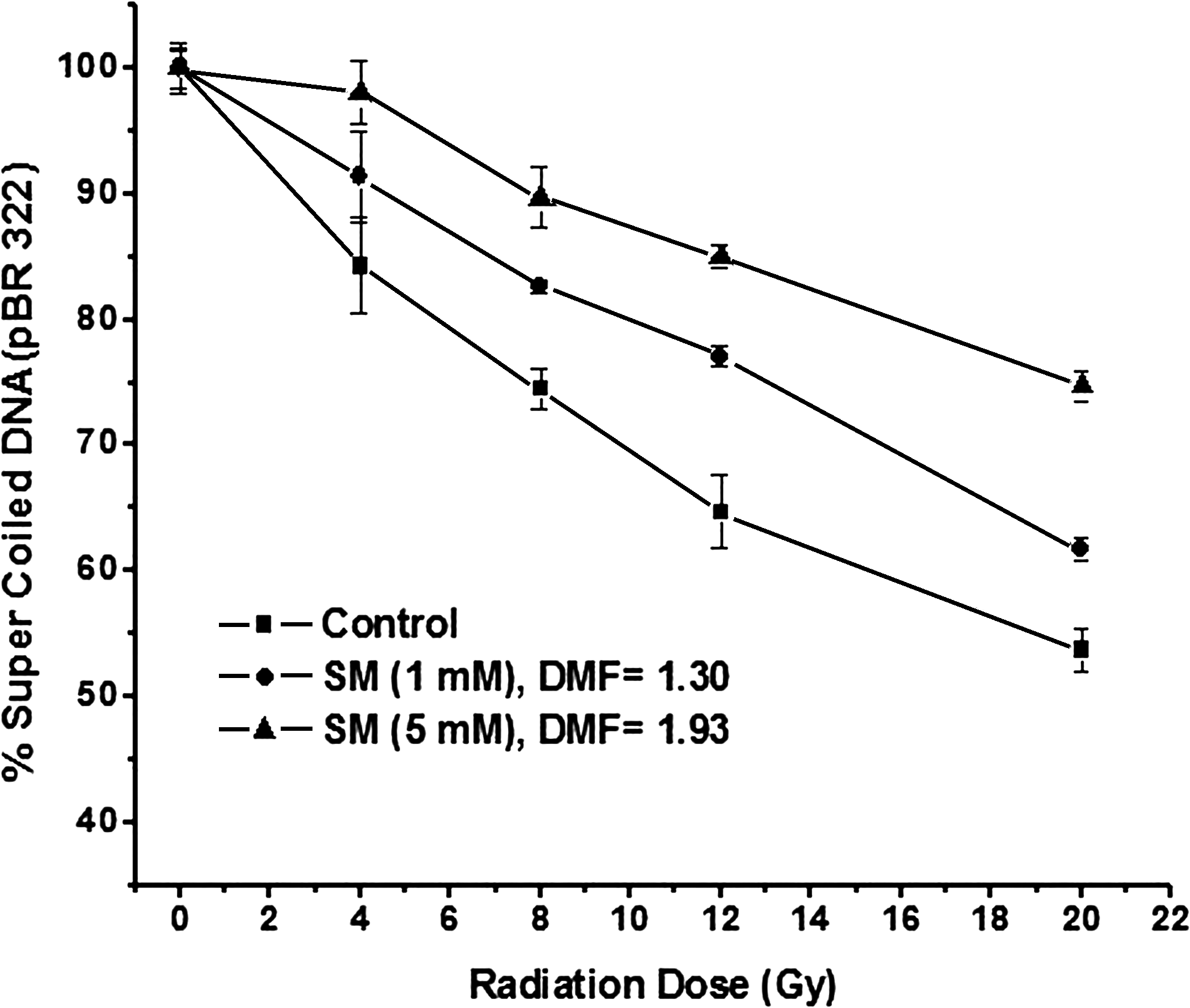
Protection of pBR 322 DNA by SM against different doses of gamma radiation (0–20 Gy).
Estimation of cellular DNA damage in mouse peripheral blood leukocytes irradiated ex vivo
Exposure to gamma radiation ex vivo induced damage to the cellular DNA in mouse blood leukocytes as evident from the comet parameters presented in Figure 4. Exposure of blood leukocytes to 4 Gy gamma radiation resulted in an increase in the comet parameters such as % DNA in tail (from 1.88 ± 0.28 to 10.99 ± 1.88), tail length (from 3.19 ± 0.40 to 16.46 ± 1.26), TM (from 0.25 ± 0.06 to 1.56 ± 0.27), and OTM (from 0.40 ± 0.04 to 1.93 ± 0.39). The presence of SM during radiation protected the cellular DNA. The protection offered by SM was dependent on its concentration, as can be evidenced from the comet parameters: % DNA in tail, tail length, TM, and OTM were brought down to the levels of 4.85 ± 1.30, 7.92 ± 0.72, 0.79 ± 0.10, and 1.17 ± 0.18, respectively (p < 0.001), in the case of 1 mM SM and 2.13 ± 0.15, 3.56 ± 0.66, 0.30 ± 0.09, and 0.46 ± 0.03, respectively (p < 0.001), in the case of 5 mM SM.

Effect of SM on DNA damage in mouse blood leukocytes induced by exposure (ex vivo) to gamma radiation (4 Gy), as assessed by comet assay. Mean of the % DNA in tail, tail length, tail moment, and olive tail moment are presented as mean ± standard deviation. “ns” indicates not significant; *p < 0.001, when compared with respective control.
Effect of SM on enhancement of cellular DNA repair in mouse blood leukocytes exposed to gamma radiation ex vivo
The effect of SM on DNA repair was studied by monitoring cellular DNA damage by comet assay ex vivo in gamma-irradiated mouse peripheral blood leukocytes incubated in DMEM with 10% FBS containing SM (5 mM) for various time intervals. From the results presented in Figure 5, it can be seen that the comet parameters such as % DNA in tail, tail length, TM, and OTM decreased at a faster rate during postirradiation incubation in the SM-treated blood leukocytes than the untreated irradiated controls. The results indicated that SM enhanced the repair of radiation-induced strand breaks in cellular DNA.
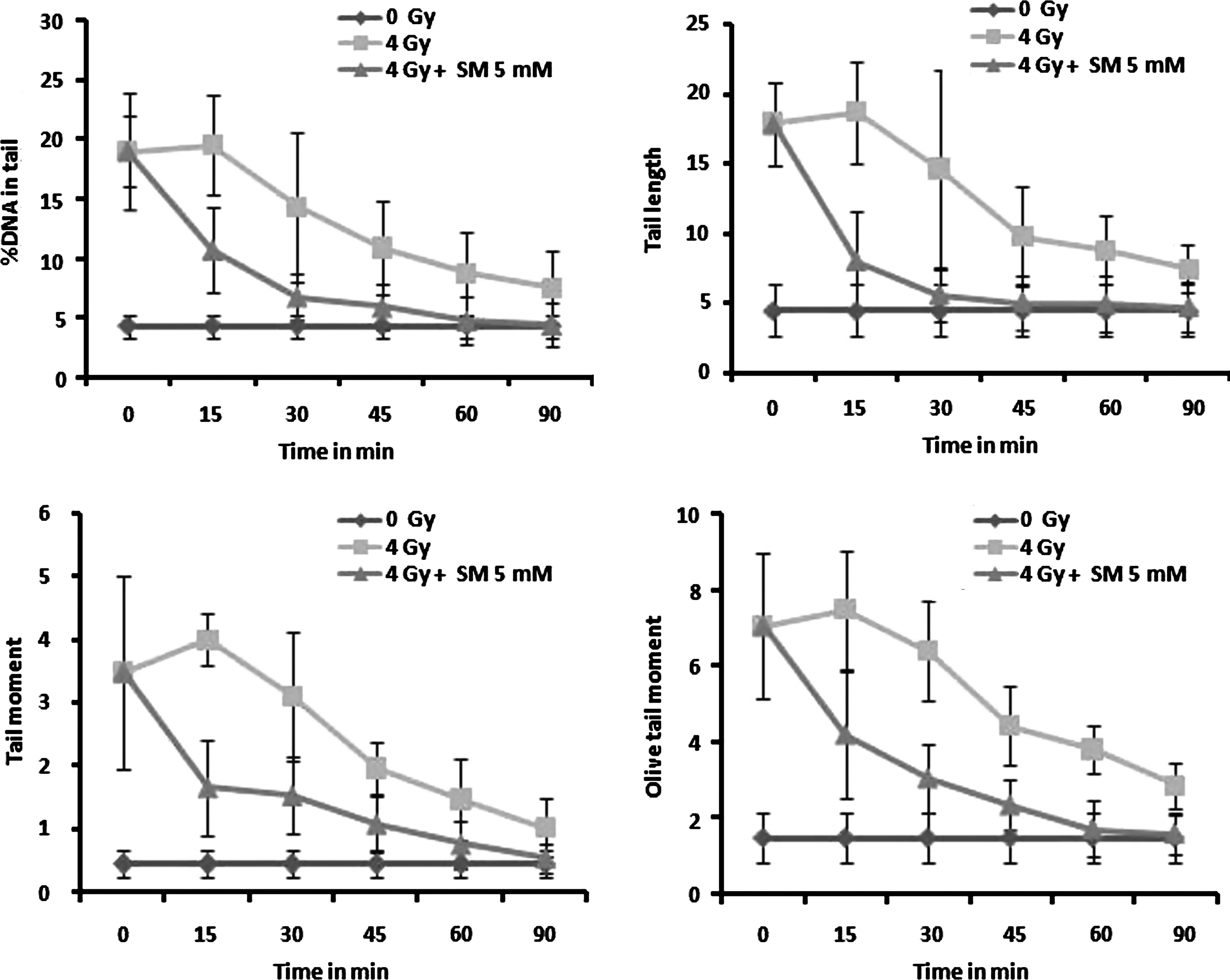
Effect of SM (5 mM) in DNA repair on mouse blood leukocytes exposed to 4 Gy gamma radiation ex vivo in terms of decrease in comet parameters—% DNA in tail, tail length, tail moment, and olive tail moment.
To quantify the efficiency of the cells to repair and rejoin strand breaks in DNA, a relation was derived for cellular DNA repair index (CRI) based on the comet parameters of the cellular DNA. CRI for a particular comet parameter is defined as the percentage decrease from the initial value of the parameter due to repair.
CRI = [1 − (Comet parameter at time t/Comet parameter at initial time t 0)] × 100.
As the rate of decrease in the comet parameters is attributed to cellular DNA repair, the efficiency of the cells to repair DNA strand-breaks following different treatments can be quantified by determining the CRI.
The data on the CRI determined from the comet parameters of mouse peripheral blood leukocytes following 4 Gy ex vivo gamma irradiation and post-treated with SM (5 mM) is presented in Figure 6. The CRI increase reflects the disappearance of DNA strand-breaks or DNA repair. In SM-treated samples (4 Gy), the CRI increased with time at a faster rate than the untreated control (4 Gy) samples. Thus, from the figure it can be observed that SM significantly enhanced cellular DNA repair efficiency.

The DNA repair enhancement expressed as cellular DNA repair index (CRI).
Protection of radiation-induced lipid peroxidation in liver tissue homogenate by SM
Exposure of mouse liver homogenate to 25 Gy gamma radiation showed significant increase in peroxidation of membrane lipids. The presence of SM during irradiation showed effective protection by inhibiting the lipid peroxide formation, measured as TBA reacting substance, illustrated in Figure 7. The MDA level in the unirradiated normal was 1.63 ± 0.67 nanomoles/mg protein, which was increased to 6.35 ± 1.62 nanomoles/mg protein in the 25 Gy gamma-irradiated samples. The presence of 5 mM SM reduced the MDA to normal level, that is, 1.65 ± 0.21 nanomoles/mg protein.
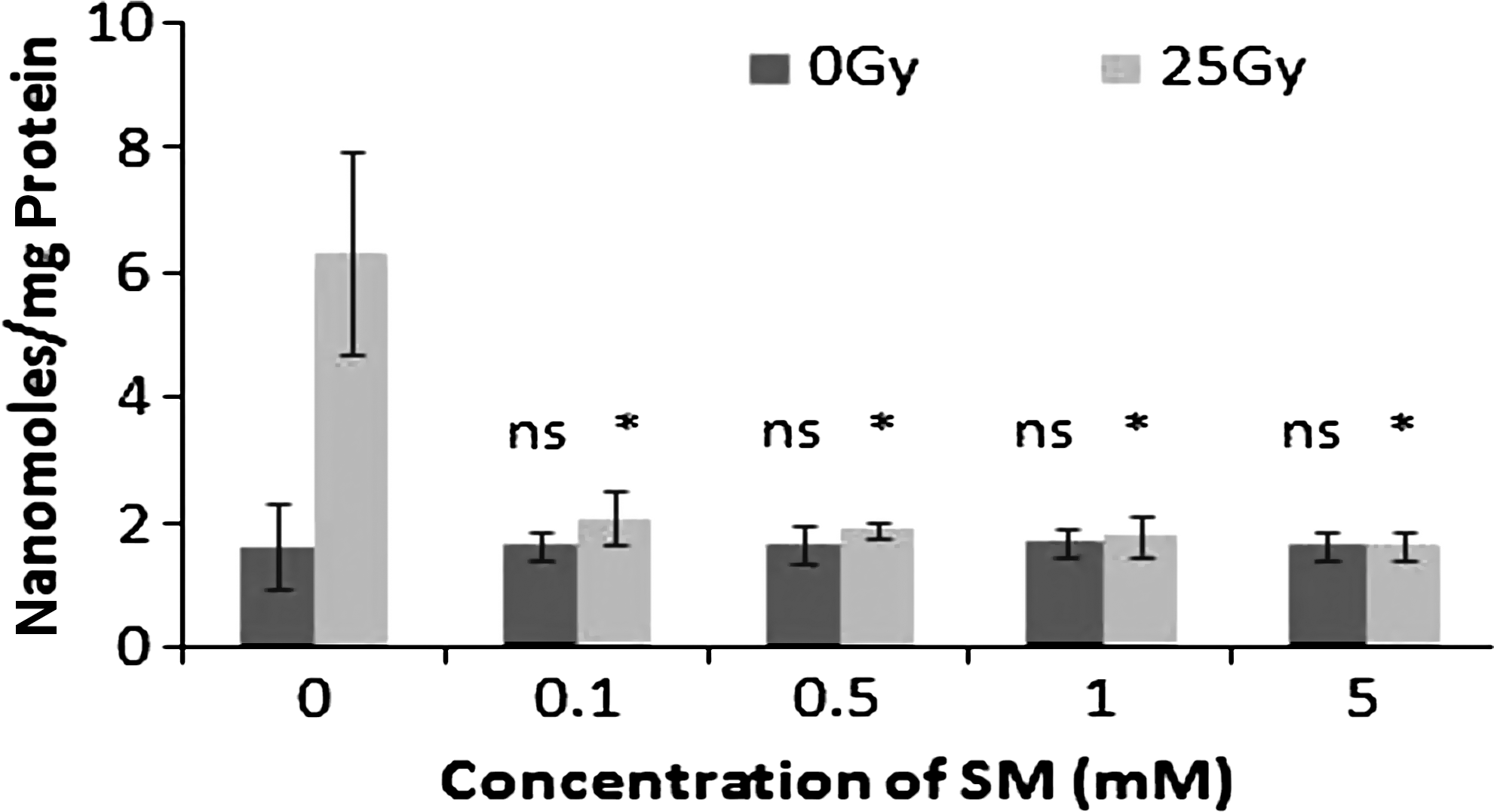
Changes in lipid peroxidation levels expressed as malondialdehyde in nanomoles/mg protein in 25 Gy gamma-irradiated mouse liver homogenate treated with different concentrations of SM. “ns” indicates not significant; * p < 0.001, when compared with respective control.
Discussion
Cellular DNA and membrane constitute primary targets of radiation inactivation in cells. Ionizing radiation–induced damages are manifested by the reaction of free radicals, mainly of the oxidative type, formed in the cellular environment. Thus, antioxidants and compounds that can scavenge free radicals can be found useful as radioprotectors. 24 However, many of the compounds that can prevent free radical activity are highly toxic to cells. Nontoxic compounds that can protect DNA from ionizing radiation are of great importance for their potential use as radioprotectors. 25 The radioprotective compounds can interfere with radiation-induced free radicals or repair the target macromolecular free radicals formed by radiation, by increasing the production of cellular antioxidant free radical scavengers or by enhancing the postirradiation cellular DNA repair process. 5 The present study consists of two aspects: (i) radioprotective effect of SM, that is, protection of DNA due to the presence of SM during irradiation along with DNA, and (ii) effect of SM on DNA repair, that is, effect of SM during postirradiation incubation of irradiated cells. The former study, that is, radioprotection, involved irradiation of cells or DNA (plasmid) in the presence of SM. The latter study, that is, effect of SM on DNA repair, involved incubation of irradiated cells (cells exposed to 4 Gy gamma radiation in the absence of SM) in complete medium containing SM. The results from the present study showed that the cellular DNA and membranes were protected from the deleterious effects of ionizing radiation by SM during in vitro and ex vivo conditions of radiation exposure. The plasmid relaxation assay performed on pBR322 DNA exposed to different doses of gamma radiation (0–20 Gy) in the presence and absence of various concentrations of SM clearly showed that the plasmid DNA was protected in vitro from radiation-induced strand-breaks in a concentration-dependent manner. It has been previously reported that the intraperitoneal administration of SM, 30 minutes prior to radiation exposure, protected mice from radiation. The protection was found to be very effective for gastrointestinal system even after a high lethal dose of 15 Gy. 16 The information on tissue distribution and elimination kinetics of SM following intraperitoneal administration in mice is not available. In the present study, the effect of SM during irradiation was examined. The cells were exposed to radiation along with SM at ice-temperature. It is presumed that the metabolism is negligible at ice-temperature, and what was observed in the present study is the radioprotective effect of SM on cellular DNA. SM was added to the cells at ice-temperature at 15 minutes prior to irradiation. SM pretreatment of lymphocytes (though no information or data are available on the quantity of SM taken up by the cells or remaining in the cells after washing) has been already reported to reduce the radiation-induced genomic DNA damage in human lymphocytes. 15
Single-cell gel electrophoresis or alkaline comet assay is a sensitive technique to monitor cellular DNA damages and is widely used to analyze the DNA lesions such as single strand-breaks, double strand-breaks, alkali-labile sites, DNA repair, and apoptosis. 26 –30 The comet parameters such as the % DNA in tail, tail length, TM, and OTM were increased when the mouse peripheral blood leukocytes were exposed to gamma radiation ex vivo. The presence of SM during irradiation of cells protected the cells from radiation injury and the comet parameters are closely similar to the levels of unirradiated cells. The data from these studies clearly demonstrated the ability of SM to protect cellular DNA from radiation-induced DNA strand-breaks in in vitro and ex vivo conditions. The results of ex vivo DNA repair studies on mouse blood leukocytes exposed to gamma radiation revealed the ability of SM to enhance the repair process. The values of the CRI were higher in the SM-treated group than in the untreated controls.
The studies on the effect of SM on radiation-induced peroxidation of membrane lipids showed that SM significantly reduced lipid peroxidation in liver tissues because of gamma radiation exposure in vitro. The results of the present investigation thus revealed the potential application of SM as a radioprotector.
Footnotes
Acknowledgments
The authors acknowledge the financial support provided by the Kerala State Council for Science, Technology, and Environment (to C.K.K.N.).
Disclosure Statement
No competing financial interests exist.