
Abstract
An agonistic antibody against TNF-related apoptosis-inducing ligand death receptor 5 (DR5) is a practicable candidate drug for antitumor therapy. In this study, a novel murine anti-human DR5 monoclonal antibody, mDRA-6(IgG1-κ), has been generated. This study aimed to explore the caspase-dependent and mitochondrial mechanisms of mDRA-6 in inducing apoptosis in human leukemia Jurkat cells. The apoptotic effects of mDRA-6 on Jurkat cells, which express DR5 on the cell surface, were detected by flow cytometry and western blot after exposure to different doses of mDRA-6 and at fixed doses of mDRA-6 at different times. It was demonstrated that mDRA-6 can induce Jurkat cell apoptosis via caspase- and mitochondrial-dependent pathways. These results indicate that the novel antibody mDRA-6 against DR5 has an antitumor function and may provide a new reagent for tumor therapy.
Introduction
TNF-related apoptosis-inducing ligand (TRAIL), also known as Apo2L, is a member of the TNF superfamily. 1 At present, five TRAIL receptors (TRAIL-R) have been identified. Among them, death receptor 4 (DR4) (TRAIL-R1) and DR5 (TRAIL-R2) have the ability to transduce the apoptosis signal, whereas decoy receptor 1 (TRAIL-R3), decoy receptor 2 (TRAIL-R4), and osteoprotegerin serve as decoy receptors and antagonists of TRAIL-mediated apoptosis. 2,3 All TRAIL receptor molecules were type I transmembrane polypeptides with 2–5 cysteine-rich domains at the extracellular region. However, only DR4 and DR5 contain the intracellular tail, and they mediate the apoptotic signals through their death domain (DD). DR4 and DR5 are two dominant apoptotic receptors of TRAIL. TRAIL activates the DRs by binding to and inducing trimerization of receptors such as DR4 and DR5. Similar to other death domain-containing receptors of the TNF superfamily, activated DR4 and DR5 can drive the Fas-associated death domain (FADD/MORT1) and procaspase-8 and/or -10 into one complex to form a death-inducing signaling complex (DISC) and mediate cell apoptosis. 4,5
The recombinant human TRAIL (rhTRAIL) and monoclonal antibodies (mAbs) against human DRs have been applied for tumor therapy in animal models or in clinical trials. 6 –8 DR4 and/or DR5 are upregulated in most tumor tissues, but not normal tissues, 9,10 unlike TNF or Fas ligand, which induce not only tumor cell apoptosis but also apoptosis in normal tissues 11,12 ; rhTRAIL or antibodies of DR5 and/or DR4 have little cytotoxicity in normal tissues. 13,14 Therefore, the development of a safe and efficacious anti-DR5 mAb will be significant for cancer therapy. 12
In the present study, a novel murine agonistic mAb, designated mDRA-6(IgG1-κ), has been generated by immunizing BALB/c mice with a purified fusion protein containing the extracellular domain of human DR5. This antibody can specifically bind to human DR5 but not other DRs and is able to induce the apoptosis of tumor cells in vitro. 15 –17 In the present study, the TRAIL-sensitive Jurkat cells were used as a target to investigate the apoptotic molecular mechanism induced by mDRA-6.
Materials and Methods
Reagents and antibodies
Jurkat cells were kindly provided by You-Hai Chen (Department of Pathology and Laboratory Medicine, School of Medicine, University of Pennsylvania). mDRA-6 was provided by Prof. Ma (Institute for Immunology of Henan University, Kaifeng, China). Fetal calf serum (FCS), RPMI1640, and methyl thiazolyl tetrazolium (MTT) were purchased from Gibco, and rhTRAIL and dimethyl sulfoxide (DMSO) were purchased from Sigma. The Annexin V-Fluorescein Isothiocyanate (Annexin V-FITC)/Propidium Iodide (PI) Kit was purchased from BD Pharmingen. The JC-1 Kit was purchased from Beyotime Company. Caspase-10 inhibitor Z-AEVD-FMK, caspase-8 inhibitor Z-IETD-FMK, caspase-3 inhibitor Z-DEVD-FMK, caspase-9 inhibitor Z-LEHD-FMK, goat anti-human caspase-10, -8, -3, and -9, and rabbit anti-human caspase-3 mAbs were purchased from R&D Systems. Rabbit anti-human FADD polyclonal antibodies and antipolyADP-ribose polymerase (PARP) mAbs were purchased from Cell Signaling Technology. The primary mAbs against BH3 interacting domain death agonist (Bid), B-cell leukemia/lymphoma 2 (Bcl-2), Bcl-2–associated X protein (Bax), cytochrome c (Cyto c), β-actin, horseradish peroxidase (HRP)-linked mouse anti-goat or anti-rabbit secondary antibodies, and an Enhanced Chemiluminescence (ECL) Kit were purchased from Santa Cruz Biotechnology.
Analysis of apoptotic morphology of Jurkat cells induced by mDRA-6
Jurkat cells were seeded and cultured in a six-well plate at a density of 2 × 105 cells/well in 1 mL of RPMI1640 medium (Gibco) supplemented with 10% heat-inactivated fetal calf serum (FCS; Gibco), penicillin (100 units/mL), and streptomycin (100 units/mL) at 37°C in a humidified atmosphere of 5% CO2 for 8 hours, followed by mDRA-6 treatment at a concentration of 200 ng/mL for 1 hour. After phase-contrast photography, the cells were collected and washed with PBS and fixed in methanol for 10 minutes. The cells were mounted onto glass slides and stained with Giemsa-Wright following the manufacturer's protocol. Cell morphology was observed using a fluorescence microscope (Axioskop 2 plus; Zeiss)
DNA fragmentation assay
After the Jurkat cells were treated as previously described for 0.5 and 1 hour, 5 × 105 cells were collected, washed once with PBS, and centrifuged at 3200 g for 5 minutes. The cells were lysed in 200 μL of buffer containing 10 mM Tris-HCl (pH 7.4), 10 mM EDTA, and 0.5% Triton X-100. Lysates were centrifuged at 12,000 g for 10 minutes at 4°C, and the supernatant was subjected to digestion with ribonuclease A (0.1 mg/mL) and proteinase K (0.1 mg/mL) for 90 minutes at 37°C. The DNA in the sample was precipitated by centrifugation at 12,000 g for 30 minutes at 4°C after treatment with 50% isopropyl alcohol and 0.5 M NaCl for 4 hours at −20°C. The DNA was resuspended in 20 μL of Tris-EDTA buffer and analyzed by electrophoresis on a 1.5% agarose gel in the presence of 0.1 μg/mL ethidium bromide.
MTT assay
Jurkat cells were cultured for 8 hours as described above in a 96-well plate in 100 μL at a density of 2.0 × 105 cells/mL. Groups of cells were treated with mDRA-6 at concentrations of 500, 250, 125, and 0.49 ng/mL. rhTRAIL was used as a positive control at the same concentration. PBS and a cell-free blank well were used as negative controls. Other groups of cells were treated with mDRA-6 at a concentration of 50 ng/mL, and cells were collected after 2, 4, 6, 8, 10, and 12 hours. After mDRA-6 treatment, the absorbance at 570 nm was measured on a microtiter reader (Thermo Labsystems). All samples were assayed in triplicate and were reported as the mean ± SD from a minimum of three independent experiments. The following formula was used to estimate the proliferation inhibition ability: blockage rate (%) = (1 − ODexperiment/ODcontrol) × 100.
Annexin V-FITC and PI staining
Jurkat cells (3 × 105) were seed into 96-well plates and treated with mDRA-6 at a concentration of 200 ng/mL for 0.25, 0.5, 1, and 2 hours. The cells were stained with Annexin V-FITC and PI according to the protocol from BD. The samples were analyzed with a flow cytometer (FACScan; BD Biosciences). For each sample, 10,000 cells were counted, and the results were analyzed with CellQuest software.
Measurement of the mitochondrial transmembrane potential (ΔΨm)
Cells were treated as described above. After washing twice with PBS, the cells were stained with mixed buffer containing 0.5 mL of 5,5′,6,6′,-tetrachloro-1,1′,3,3′,-tetraethyl benzimidazol carbocyanine iodide (JC-1) dye and 0.5 mL RPMI1640 in a 37°C incubator with 5% CO2 for 20 minutes. The cells were washed twice with 1 × staining buffer and resuspended in 400 μL of binding buffer. The early apoptosis rate of the Jurkat cells was analyzed by flow cytometer, and results were analyzed with CellQuest software as described above.
Blocking effect of caspase inhibitor versus mDRA-6–induced apoptosis
Cells (3 × 105) in 1 mL of RPMI1640 culture medium were plated in a 96-well plate. After the cells were maintained at 37°C in the presence of 5% CO2 for 8 hours, caspase-10 inhibitor Z-AEVD-FMK, caspase-8 inhibitor Z-IETD-FMK, caspase-3 inhibitor Z-DEVD-FMK, and caspase-9 inhibitor Z-LEHD-FMK were added at a concentration of 10 μM, and the cells were maintained in the CO2 incubator for 2 hours. Then, the Jurkat cells were incubated with 200 ng/mL mDRA-6 for 12 hours, and the same concentration of mDRA-6 was used as the positive control without caspase inhibitors. PBS and the cell-free blank well were used as negative controls. After the treatment, the absorbance at 570 nm was measured on a microtiter reader as describe above with the MTT assay. All samples were assayed in triplicate and were reported as the mean ± SD from a minimum of three independent experiments. The following formula was used to estimate the apoptosis inhibition ability: apoptosis blockage rate (%) = cell viability percentage of the experimental group − cell viability percentage of the positive control group.
Western blotting
Jurkat cells were treated with mDRA-6 for 0.25–2 hours just as Annexin V-FITC and PI described above. Cytosolic and mitochondrial fractions were prepared as previously described. 18 Briefly, the cells were resuspended in 500 μL buffer (2 mM Hepes-potassium hydroxide [KOH] [pH 7.5], 10 mM MgCl2, 1 mM ethyleneglycol bis(2-aminoethyl ether) tetraacetic acid [EGTA], 1 mM dithiothreitol [DTT], and 250 mM sucrose and protease inhibitor). After homogenization, the unbroken cells, large plasma membrane pieces, and nuclei were removed by centrifugation at 1000 g for 10 minutes. The supernatant was subjected to centrifugation at 12,000 g for 15 minutes. The pellet fraction containing mitochondria was resuspended in 400 μL buffer (10 mM Tris-acetate [pH 8.0], 0.5% Nonidet NP-40, and 5 mM CaCl2) and subsequently incubated on ice for 30 minutes. The supernatant was further centrifuged at 50,000 g for 2 hours to generate the cytosol at 4°C. Cyto c in the fractions of cytosol and mitochondria was detected by western blot. To detect the total cellular protein, the cells were lysed in the buffer (containing 150 mM NaCl, 10 mM Tris-base [pH 7.2], 5 mM EDTA, 0.1% Triton-100, 5% glycerol, and 2% SDS). Equal amounts of protein (50 μg/lane) were fractionated on 12% or 15% SDS-PAGE, followed by transfer to a polyvinylidene difluoride (PVDF) membrane (NEN Life Science). The membranes were blocked with 2% nonfat dry milk blotting buffer (25 mM Tris-HCl [pH 7.4], 0.15 M NaCl, and 0.1% Tween-20) for 1 hour at room temperature, and the membranes were then incubated with the indicated primary antibodies against FADD, caspase-10, -8, -3, and -9, PARP, Bid and truncated Bid (tBid), Bax, Bcl-2, and Cyto c (1:2000) for 2 hours at room temperature. The membranes were then incubated with HRP-conjugated secondary antibodies. The signals were developed in ECL buffer and examined on a Kodak X-ray film.
Statistical analysis
The results were obtained from at least three independent experiments. Student's t-test was used for evaluating statistical significance. SPSS 13.0 software was used, and p-values <0.05 were considered statistically significant.
Results
mDRA-6 induces the apoptotic morphology of Jurkat cells
To determine the apoptotic role of mDRA-6, Jurkat cells were treated with mDRA-6 at a concentration of 200 ng/mL, and the cell morphology was observed using a phase-contrast microscope. In contrast to the cells in the group with no mDRA-6 treatment, which showed a good, round refraction, intact membrane, and a unique cell size, the morphology of mDRA-6–treated cells started to change by 0.25 hour of treatment, and serious apoptotic morphology was observed at 1 hour of treatment, including cell shrinkage, membrane crenation, and blebbing (Fig. 1A, B). Giemsa-Wright staining showed a condensation of nuclear chromatin and nuclear fragmentation (apoptotic bodies) in the mDRA-6–treated Jurkat cells (Fig. 1 C, D).

Morphological observation of the Jurkat cells by inverted contrast microscopy (400 × magnification). Cells were incubated with mDRA-6 at a concentration of 200 ng/mL for 1 hour. (
mDRA-6 induces a DNA ladder of Jurkat cells
To determine whether mDRA-6 could induce cell apoptosis, the Jurkat cells were treated with mDRA-6 for 0.5 and 1 hour. PBS was used as a negative control. The DNA was extracted and separated on agarose gel. The DNA fragmentation was visualized under UV light. As indicated in Figure 2, no DNA fragmentation was observed in the control cells, but in contrast, DNA fragmentation was induced between 0.5 and 1 hour, during which the Jurkat cells were incubated with 200 ng/mL of mDRA-6 (Fig. 2).

Analysis by agarose gel electrophoresis of DNA ladder of apoptotic Jurkat cells with mDRA-6 treatment at a concentration of 200 ng/mL for 0.5 and 1 hour. 1: Negative control; 2: Jurkat cells with mDRA-6 treatment for 0.5 hour; 3: Jurkat cells with mDRA-6 treatment for 1 hour; 4: marker.
The mDRA-6–induced cell death was further analyzed with the MTT assay
The Jurkat cells were treated with mDRA-6 for 12 hours at the following concentrations: 0.49, 0.98, 1.95, 3.9, 7.8, 15.6, 31.3, 62.5, 125, 250, and 500 ng/mL. The death rates of Jurkat cells were 6.7%, 7.5%, 10.9%, 17.5%, 31.3%, 50.1%, 72.0%, 86.0%, 91.9%, 92.9%, and 92.3%; the same concentrations of rhTRAIL were used to inhibit the growth of Jurkat cells, and the death rates of the Jurkat cells were 7.2%, 8.3%, 13.3%, 16.0%, 25.6%, 32.3%, 48.7%, 64.7%, 79.5%, 81.99%, and 85.5% (Fig. 3). The mDRA-6–induced cell apoptosis was also determined by treating the Jurkat cells with 50 ng/mL mDRA-6 for 2, 4, 6, 8, 10, and 12 hours. The death rates for the Jurkat cells were 8.9%, 28.9%, 55.4%, 71.8%, 83.0%, and 85.9%. The same concentrations of rhTRAIL were used to inhibit the growth of Jurkat cells, and the death rates of Jurkat cells were 9.2%, 17.4%, 38.8%, 47.2%, 52.0%, and 58.1% (Fig. 4).

The mDRA-6 and rhTRAIL dose-dependence of Jurkat cell death. The Jurkat cells were treated with mDRA-6 for 12 hours at the following concentrations: 0.49, 0.98, 1.95, 3.9, 7.8, 15.6, 31.3, 62.5, 125, 250, and 500 ng/mL. The death rates of Jurkat cells were 6.7%, 7.5%, 10.9%, 17.5%, 31.3%, 50.1%, 72.0%, 86.0%, 91.9%, 92.9%, and 92.3%, respectively; rhTRAIL was used as a positive control at the same concentrations. The death rates of the Jurkat cells were 7.2%, 8.3%, 13.3%, 16.0%, 25.6%, 32.3%, 48.7%, 64.7%, 79.5%, 81.99%, and 85.5%, respectively, using rhTRAIL. TRAIL, TNF-related apoptosis-inducing ligand.
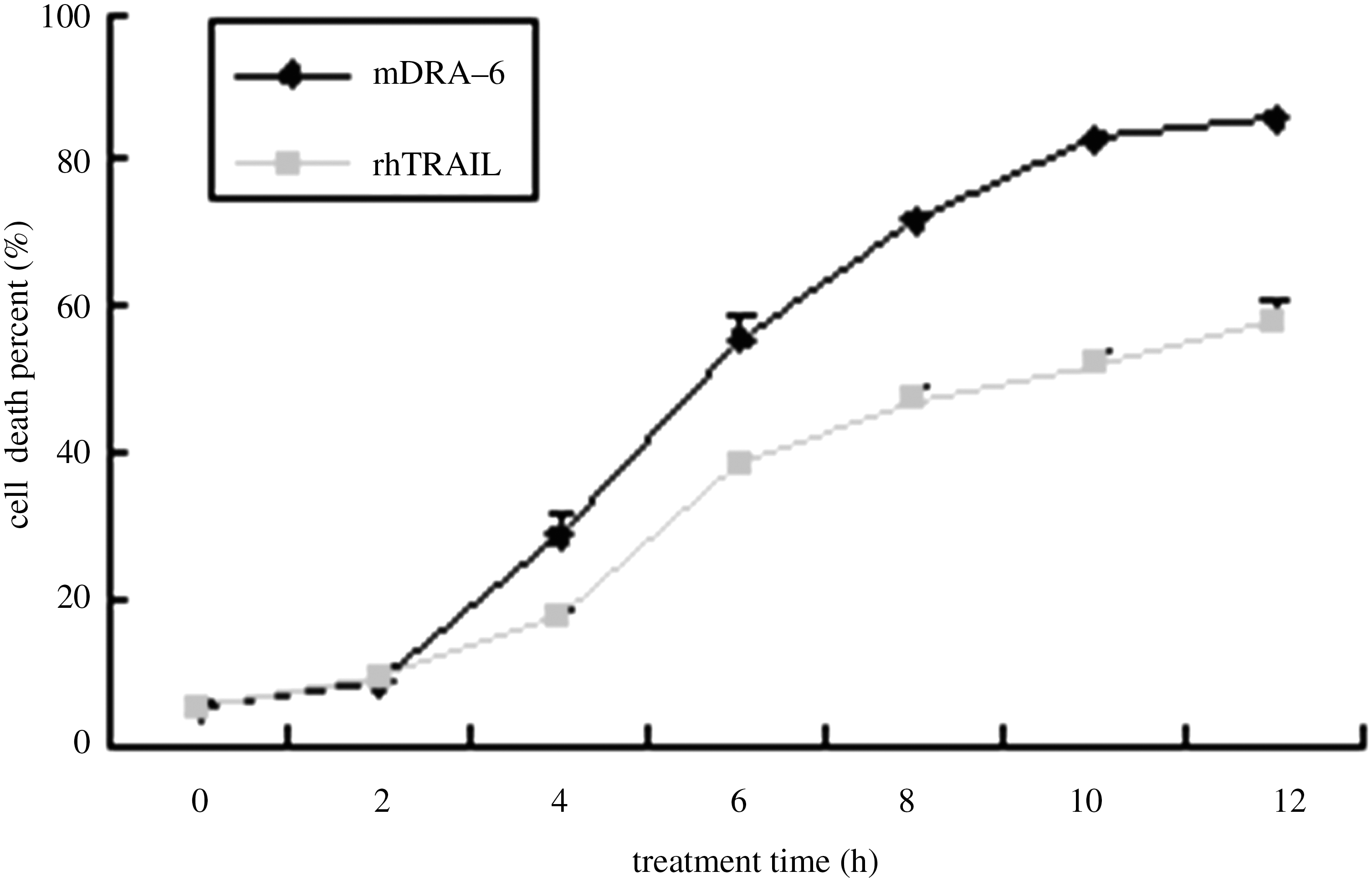
The mDRA-6 and rhTRAIL treatment time-dependence of Jurkat cell death. mDRA-6–induced cell apoptosis was also determined by treating Jurkat cells with 50 ng/mL mDRA-6 for 2, 4, 6, 8, 10, and 12 hours. The death rates for Jurkat cells were 8.9%, 28.9%, 55.4%, 71.8%, 83.0%, and 85.9%, respectively. The same concentrations of rhTRAIL inhibited the growth of Jurkat cells, and the death rates of the Jurkat cells were 9.2%, 17.4%, 38.8%, 47.2%, 52.0%, and 58.1%, respectively.
mDRA-6–induced cytotoxicity of Jurkat cells by Annexin V/PI staining
In contrast to the apoptotic rate of the negative control, the apoptotic rate of Jurkat cells reached 17.1% (the early apoptosis rate was 8.7%, and the late apoptosis rate was 8.5%), 30.2% (early and late rates were 17.4% and 12.9%, respectively), 69.0% (early and late rates were 30.6% and 38.5%, respectively), and 79.6% (early and late rates were 38.6% and 41.0%, respectively) when incubated with mDRA-6 for 0.25, 0.5, 1, and 2 hours, respectively, at the concentration of 200 ng/mL. Different labeling patterns in this assay enabled to identify different cell populations by Annexin V/PI staining: vital cells (PI−/Annexin V−); early apoptotic cells (PI−/Annexin V+); and cells undergoing apoptosis/necrosis (PI+/Annexin V+). This outcome suggested that the novel agonistic mAb mDRA-6 can induce Jurkat cells that are sensitive to TRAIL-mediated apoptosis (Fig. 5).

Annexin V and propidium iodide staining flow cytometry. Jurkat cells were treated with 200 ng/mL of mDRA-6 for different durations. The results showed that the apoptotic rate of Jurkat cells increased by mDRA-6 treatment from 0.25 to 2 hours.
FACS analysis of JC-1
The changes in ΔΨm were monitored with the JC-1 dye. JC-1 emitted light at red and green wavelengths according to the concentrations absorbed into the mitochondria; at high concentrations, the J-aggregated form emits a red light in the mitochondrial matrix, whereas at low concentrations, the monomer form emits a green light in the mitochondrial matrix. Therefore, the red and green florescence of the JC-1 reflects the change of the ΔΨm of the mitochondrial membrane. The decrease in mitochondrial membrane potential was a sign of an early cell apoptosis. Therefore, the fluorescent probe JC-1 could be used as an early apoptosis detection factor. As illustrated in Figure 6, mDRA-6 could induce the apoptosis of Jurkat cells through the mitochondrial pathway. In contrast to the negative control, the early apoptosis rate of the Jurkat cells reached 20.1%, 19.3%, 21.1%, and 30.9% when incubated with mDRA-6 for 0.25, 0.5, 1, and 2 hours, respectively (Fig. 6).

Analysis of Jurkat cell apoptosis upon treatment with mDRA-6 using JC-1 staining by flow cytometry. Jurkat cells were treated with 200 ng/mL mDRA-6 from 0.25 to 2 hours. The results showed that the apoptotic rate of Jurkat cells increased because of the mDRA-6 treatment through the mitochondrial pathway.
The blocking effect of caspase inhibitor against mDRA-6–induced apoptosis
When caspase inhibitors were used to block the cytotoxicity of mDRA-6 in Jurkat cells, the results were different. The cellular viable ratio of caspase-10 inhibitor Z-AEVD-FMK was 11.77%, whereas the ratio in the positive control was 10.74%. The results shown were not considered to be statistically significant (p = 0.672). Then, caspase-8 inhibitor Z-IETD-FMK, caspase-3 inhibitor Z-DEVD-FMK, and caspase-9 inhibitor Z-LEHD-FMK were tested, and the viability ratio of Jurkat cells was found to be 88.6% (p < 0.01), 64.9% (p < 0.01), and 19.5% (p < 0.05), respectively. It was concluded that caspase-8 was the upstream factor in the caspase-dependent apoptosis pathway of Jurkat cells induced by mDRA-6. Caspase-3 and -9 were the downstream factors in the apoptosis of Jurkat cells. Unexpectedly, in this process, caspase-8 was involved as the initial trigger, not caspase-10 (Fig. 7).
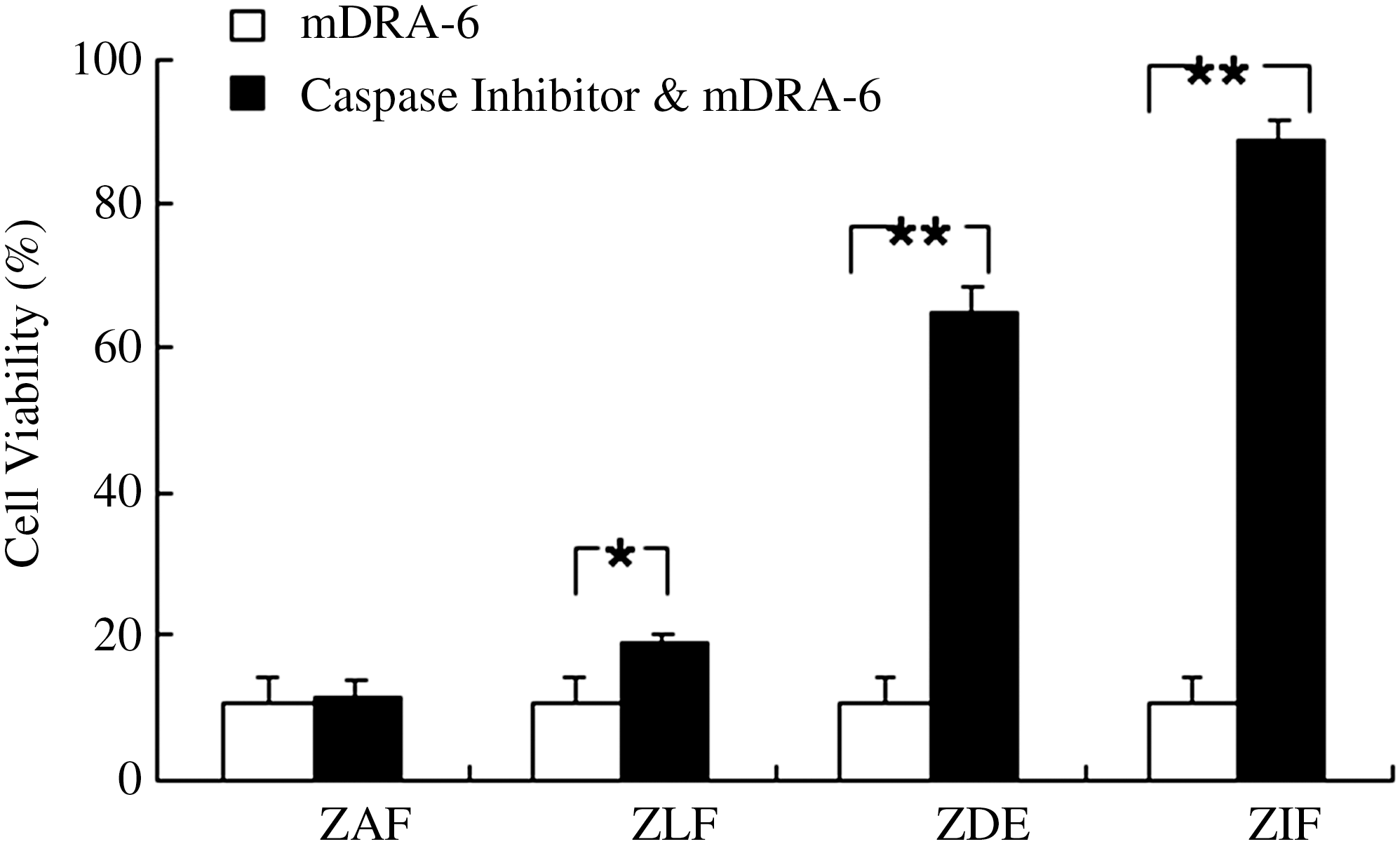
The effect of caspase inhibitors on Jurkat cell apoptosis induced by mDRA-6 as analyzed by MTT assay. Jurkat cells were incubated with 200 ng/mL mDRA-6 for 12 hours, or they were incubated with 10 μM of different caspase inhibitors for 2 hours and then 200 ng/mL mDRA-6 for 12 hours (ZAF represents caspase-10 inhibitor Z-AEVD-FMK; ZLF represents caspase-9 inhibitor Z-LEHD-FMK, ZDE represents caspase-3 inhibitor Z-DEVD-FMK, and ZIF represents caspase-8 inhibitor Z-IETD-FMK). *p < 0.05, **p < 0.01.
The caspase- and mitochondrial-dependent apoptotic pathway of Jurkat cells induced by mDRA-6
The expression of the FADD protein was upregulated between 0.25 and 2 hours. The active cleavage products of caspase-8, −3, and −9 significantly increased compared with untreated cells in a time-dependent manner, whereas mDRA-6 had no effect on the activity of caspase-10 or the corresponding increase in active cleavage products of PARP after mDRA-6 exposure between 0.25 and 2 hours. Likewise, the expression of the Bid protein diminished, whereas tBid increased. Cyto c release from the mitochondria was enhanced concomitant with the related attenuation of Cyto c in the mitochondria. The expression of Bcl-2 protein was downregulated, and Bax was upregulated from 0.25 to 2 hours. TRAIL is known to activate caspases in various cell lines. Naturally, the authors wondered whether mDRA-6 induced the same response. Western blot analysis showed that mDRA-6 activated the caspase cascade in Jurkat cells as did TRAIL. Caspase-8, -3, and -9 and PARP were cleaved in a time-dependent manner within 2 hours, but active caspase-10 was not produced. In this process, caspase-8 is involved as the initial trigger, not caspase-10. These results were confirmed using a caspase inhibitor and showed that mDRA-6 could induce Jurkat cell apoptosis in a typical caspase pathway. At the same time, Bid, Bax, Bcl-2, and Cyto c were differentially changed, and the mitochondrial-dependent apoptosis pathway was also activated (Fig. 8).
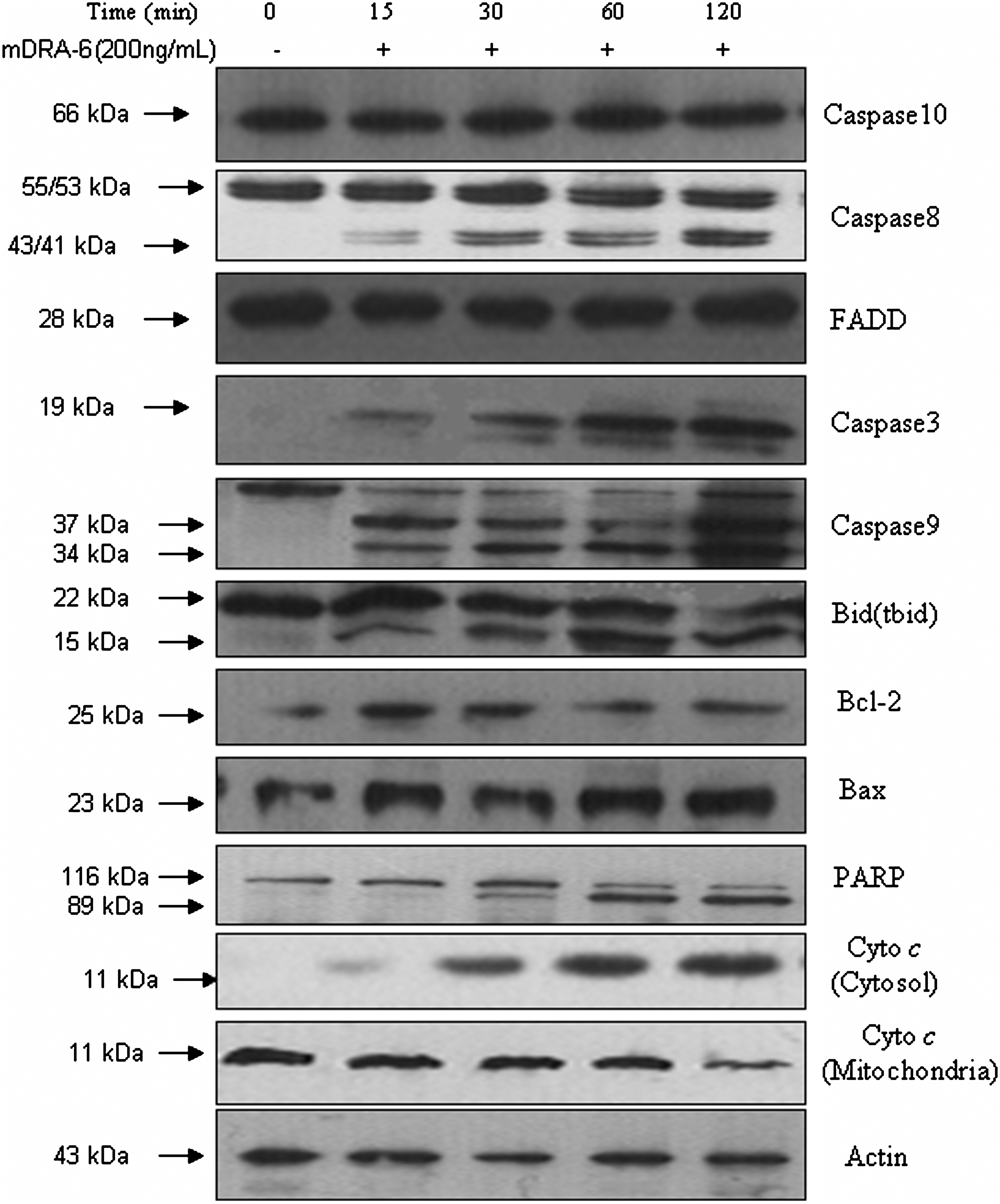
Activation of the caspase cascade and apoptotic proteins in Jurkat cells after treatment of 200 ng/mL mDRA-6 for the indicated times. The activation of caspase-10, −8, −3, and −9, FADD, PARP, Bid (tbid), Bxl-2, Bax, and Cyto c was detected by western blot. Equal amounts (50 μg/lane) of cellular proteins were fractionated on 12% or 15% SDS-PAGE gels and transferred to PVDF membranes. Actin was used as an internal control.
Discussion
In the present study, mDRA-6, a novel agonistic mAb against human DR5, has been developed. This antibody binds specifically to DR5 and does not cross-react with rhDR4, rhDcR1, rhDcR2, and rhFas, and rhDR5 can inhibit the apoptotic effect of mDRA-6 on Jurkat cells. 15 The authors' earlier work showed that mDRA-6 induced apoptosis in various tumor cell lines in the absence of crosslinking 16,17 and had a synergistic effect on tumor cell apoptosis when combined with chemotherapeutic agents. 19,20
The present study showed that mDRA-6 can induce Jurkat cell apoptosis and that Jurkat cells exhibited typical apoptotic features in morphology, including cell shrinkage, bubbling, chromatin condensation, formation of apoptotic bodies (Fig. 1), and DNA fragmentation changes (Fig. 2). The flow cytometry analysis of apoptosis using Annexin V/PI and JC-1 staining methods revealed that at a concentration of 200 ng/mL after 0.25 hour, mDRA-6 induced Jurkat cell apoptosis (Figs. 5 and 6). These results indicated that mDRA-6 could induce Jurkat cell apoptosis.
Receptor oligomerization was considered to be necessary for the death signals mediated by DRs. The fact that some anti-DR mAbs required exogenous crosslinking to ensure their tumoricidal activity in previous studies seemed to favor this argument. However, mDRA-6 as well as the previously reported mAb TRA-813 and mAb AD5-10 21 were able to kill tumor cells without crosslinking, raising questions for this hypothesis.
TRAIL has been thought to be a potential antitumor drug because of its powerful ability to kill tumors. As research continues in the laboratory, clinical application of TRAIL has been halted because of its limitations. The mAbs against DR4 or DR5 may be better candidates as therapeutic agents. It was observed that mDRA-6, similar to TRA-8, activated the conventional caspase cascade in Jurkat cells and caused a classical caspase-dependent cell death. 13 The activation of the caspase cascade is the most important part of the cell signals triggered by TRAIL and constitutes the basic function of DR5. Interestingly, caspase-10 inhibitor Z-AEVD-FMK failed to inhibit the cell death induced by mDRA-6, and caspase-8 inhibitor Z-IETD-FMK completely inhibited the cell death induced by mDRA-6 (Fig. 7), indicating that mDRA-6 activated the classical caspase-dependent signaling pathway. In this process, caspase-8 was involved as the initial trigger, not caspase-10, and western blot analysis of the caspase cascade cleavage (Fig. 8) further demonstrated that typical caspase-8 was the initial trigger to induce cell death in Jurkat cells treated with mDRA-6.
Apoptosis can be triggered by the mitochondrial pathway or the membrane DR pathway. 22 In the membrane DR pathway, of the many disparate death stimuli, the signaling pathway initiated by TRAIL (also Apo-1) has been well characterized. 23 These cytokines are expressed as membrane-bound ligands that can be cleaved into a soluble form. Engagement by these ligands of their respective receptors, followed by the recruitment of an adapter molecule such as FADD, 24,25 leads to direct activation of the caspase cascade. This activation is accomplished by the recruitment of procaspase-8 and/or -10, followed by their proteolytic activation. Once activated, caspase-8 and/or -10 can proteolytically cleave Bid (a pro-apoptotic Bcl-2 family member containing only the BH-3 domain; it is exclusively a cytosolic protein and can be cleaved by caspase-8 or -10), and the truncated Bid targets the mitochondria, induces Cyto c release, and further amplifies the caspase cascade. 26 –28 The engagement of the mitochondria through truncated Bid by receptor-mediated apoptosis further supports the pivotal role of mitochondria in apoptosis. Interestingly, Bcl-2 is a proteolytic target of caspases and is converted in situ to a proapoptotic molecule to promote Cyto c release and apoptosis. 29,30
To reveal the precise molecular mechanism of Jurkat cells induced by mDRA-6, the expression of FADD and Cyto c and the activity of the caspases and PARP, as well as the Bcl-2 family, were observed. The present results showed that mDRA-6 increased the level of FADD, PARP, and Cyto c in the cytoplasm with the corresponding decrease of Cyto c in the mitochondria. To determine whether caspases are involved in mDRA-6–induced apoptosis in Jurkat cells, the catalytic activity and expression of caspase-10, -8, -9, and -3 were measured. The data demonstrated that mDRA-6 could activate caspase-8, -9, and -3. However, the activity of caspase-10, another apoptosis-initiating protease linked to the DRs, was unchanged, and the cleaved fragment of caspase-10 was not detected. mDRA-6 has been suggested to induce apoptosis via both the membrane DR pathway and the mitochondrial pathway. Further, the data showed that mDRA-6 activated the caspase cascade in Jurkat cells just as TRAIL, FADD, caspase-8, -3, and -9, and PARP were cleaved in a time-dependent manner within 2 hours, but caspase-10 was not found to have active cleavage products. In this process, caspase-8 was involved as the initial trigger, not caspase-10.
The Bcl-2 family plays a pivotal role in releasing Cyto c and the apoptosis mediated by the mitochondria. 31 More than 20 Bcl-2 family members have antiapoptosis membranes (including Bcl-2 and Bcl-xL), and proapoptosis membranes (including Bid, Bax, and Bak) have also been identified. Bcl-2 is an important element during apoptosis that is mediated by the mitochondrial pathway and has been identified as preventing Cyto c release from the mitochondria 32,33 and preventing the activation of caspase-3. 34 In contrast, Bax can induce the release of Cyto c from the mitochondria. 32,33 Bax and Bid can promote Cyto c release from the mitochondria and thus activate the caspase cascade. 26 –28 The interactions between pro- and antiapoptotic molecules were found to be the determining factors for cell survival. Not surprisingly, most death modulators function by acting through the Bcl-2 family of proteins that regulate Cyto c release. 35,36 A decrease in the full-length Bid and an increase in tbid between 0.25 and 2 hours of mDRA-6 treatment were observed, and the mechanism may be the cleavage of Bid to tbid. The Bid cleavage occurred at the same time as the activation of caspase-8 by mDRA-6 treatment, and then tbid triggered a time-dependent increased release of Cyto c from the mitochondria (Fig. 8). The present study revealed that mDRA-6–induced apoptosis is accompanied by an increased expression of Bax and a reduced protein level of Bcl-2 in Jurkat cells. The present study's results suggested that the caspase-8–dependent Bid cleavage in mDRA-6–induced apoptosis may trigger the mitochondrial pathway by releasing Cyto c to induce caspase-9 activation in the cytosol. Cyto c release into the cytosol initiates the formation of “apoptosome,” which consists of Cyto c and caspase-9. This complex induces the activation of caspase-9, which sets off a cascade to cleave and activate effector caspases.
Conclusions
In summary, the present study demonstrated that mDRA-6 induced the apoptosis of Jurkat cells via the typical caspase- and mitochondrial-dependent pathways, and the Bcl-2 families were involved in the control of apoptosis (Fig. 9). Different cell signals induced by mDRA-6, mAb TRA-8, 13 and mAb AD5-10 21 and the synergistic action of TRAIL and mDRA-6 in killing tumor cells demonstrate the complexity of the cell signals mediated by DR5. The structure and function of DR5 are much more complicated than what was considered. The present study put forward DR5 as a more attractive research target, and further exploration of the mechanisms of mDRA-6 will add timely new knowledge for research on programmed cell death and cancer immunotherapy. The present study provided evidence to support the humanization of mDRA-6 and subsequent laboratory research. The humanized version of mDRA-6 may be a potential biotherapeutic in future, and the TRAIL-based therapeutic approaches look more promising.
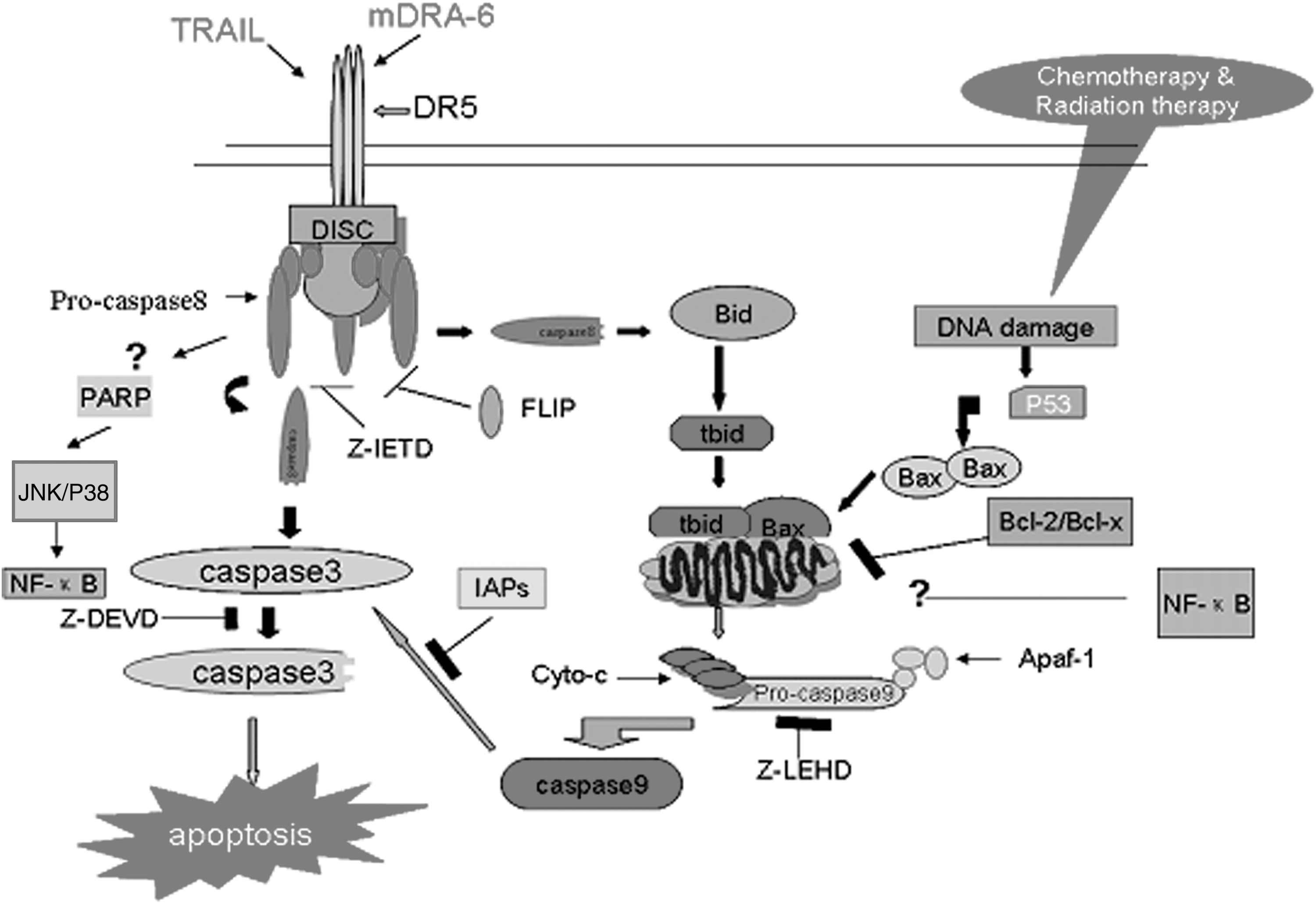
The molecular mechanisms of mDRA-6 inducing apoptosis in Jurkat cells. mDRA-6 induces aggregation of death domains that reside on the cytoplasmic sides of TRAIL-R2 (death receptor 5), which in turn recruit adaptor proteins such as FADD, and thus formed a DISC. The latter binds the prodomains of caspase-8 and may lead to the activation of caspase-3 and cleavage of Bid. Bid cleavage-induced apoptosis may trigger the mitochondrial pathway by releasing Cyto c to induce caspase-9 activation in the cytosol. This finding indicates that caspase-8 is working as an apical initiator upstream of caspase-dependent cell apoptosis in Jurkat cells induced by mDRA-6 treatment, and caspase-8 is also working as an apical initiator upstream of the caspase-independent and mitochondrial pathway.
Footnotes
Acknowledgments
This work was supported in part by the Natural Science Foundation of China (Grant No. 30571697), National High Technology and Development Program of China (863 Program Grant No. 2006AA02A254), and the Outstanding Individual Innovation Foundation of Henan Province (No. 074200510014).
Disclosure Statement
No conflicts of interest exist for any of the authors.