
Abstract
The objective of this study was to investigate the therapeutic potential of 131I added to doxorubicin therapy in multidrug resistance (MDR) mouse colon cancer coexpressing the MDR1 small hairpin RNA (shRNA) and human sodium iodide symporter (hNIS) gene in a single gene construct and to visualize the antitumor effects using molecular nuclear imaging. HCT-15 coexpressing shRNA for MDR1 gene (MDR1 shRNA) and hNIS gene with a single construct was established (referred to as MN61 cell). Inhibition of P-gp function by MDR1 shRNA and functional activity of hNIS gene was assessed using a 99mTc sestamibi uptake and 125I uptake, respectively. Cytotoxic effects by a combination of doxorubicin and 131I were determined in parental (HCT-15) or MN61 cells using an in vitro clonogenic assay. Therapeutic effect of either combination therapy (doxorubicin and 131I) or single therapy (doxorubicin or 131I alone) was evaluated by tumor volume measurement. 99mTc-sestamibi, 123I, and 99mTc-pertechnetate images of mice were acquired to evaluate functional assessment in vivo. Cellular uptake of 99mTc-sestamibi and 125I was approximately 2-fold and 100-fold higher in MN61 cells than in parental cells, respectively. Combination of 131I and doxorubicin resulted in higher cytotoxcity in MN61 cells as compared with parental cells. Scintigraphic imaging showed higher uptake of 99mTc-sestamibi and 123I in MN61 tumor as compared with parental tumor. In mice treated with doxorubicin, there was a slight delay in tumor growth in the MN61 tumor but not in the parental tumor. Cancer treatment with 131I or doxorubicin induced a rapid reduction of tumor volume in the MN61 tumor but not in the parental tumor. Combination therapy further generated a rapid reduction of tumor volume as compared with 131I therapy alone (p < 0.05). A combination hNIS mediated radioiodine gene therapy added to MDR1 shRNA treatment improved the effects of cancer treatment in a MDR cancer model and could enable visualization of the antitumor effects with nuclear imaging.
Introduction
Although single therapy is effective in inhibiting tumor growth in patients with cancer, substantial cases of metastatic tumor or bulky tumors are rarely treated with single therapy alone. Various multimodality strategies have been applied to treat patients with cancer who are not controlled with single therapy either due to resistance to specific therapy or due to serious damage to normal organs. 1 –7
Human sodium iodide symporter (hNIS) gene is responsible for expression of an intrinsic transmembrane glycoprotein that serves as a driving force for iodide uptake. 8 When hNIS gene is introduced to nonthyroidal tumors using nonviral or viral systems, therapeutic radioisotopes such as 131I-sodium iodide and 188Re-perrhenate can be made to accumulate in hNIS expressing tumors without damaging the surrounding cells, leading to impartation of strong antitumor effects. Further, hNIS is proved to act as a nuclear reporter gene that is commonly used in visualizing the antitumor effects of radioiodine gene therapy using gamma camera or PET. 9 Conventional PET reporter genes require the synthesis of complicated substrates and an expensive PET facility, whereas hNIS-mediated gamma nuclear imaging has the beneficial ability to image with radiotracers such as Tc-99m pertechnetate and radioiodine and it would potentiate its clinical application. 10 –12 99mTc-pertechnetate has been commonly used as a substrate for hNIS-mediated imaging, because it is a readily available and cheap radiotracer with good physical characteristics suitable for nuclear gamma imaging.
Chemotherapy using cytotoxic agents plays an important role in the treatment of various tumors. However, cancers that overexpress multidrug resistance 1 (MDR1) gene do not easily respond to chemotherapeutic reagents, because P-glycoprotein (P-gp, a member of the ATP-binding cassette transporter superfamily) encoded by MDR1 gene pumps therapeutic agents out of the cells. 13 Several studies have been performed to block the function of P-gp, and most efforts are being focused especially on chemical compounds that modulate P-gp activity. However, it has not been successful to apply these compounds in the clinical phase, mainly due to its inherent systemic adverse effects. Therefore, an alternative strategy to circumvent MDR is under investigation. Gene therapeutic approaches with antisense oligonucleotides (ODNs), ribozymes, neutralizing antibodies, and, most recently, the application of the RNA interference (RNAi) technology have been investigated. 14,15
RNAi is a conserved cellular mechanism by which double-stranded RNA silences the corresponding homologous gene. Many reports have shown promising therapeutic potential of small hairpin RNA (shRNA) in modulating the function of P-gp in several cancers associated with MDR. 16 –18
It was postulated that combination of hNIS radionuclide gene therapy and chemotherapy mediated by RNAi-mediated MDR1 gene silencing could be a more effective strategy for treatment of patients with cancer who were not responsive to single therapy. Previously, by using the vector coexpressing hNIS gene and shRNA for MDR1 gene, the therapeutic potential of the combination of hNIS radioiodine therapy and shRNA for MDR1 gene (MDR1 shRNA) in MDR cancer cells in vitro was successfully reported. 19 Although this strategy has proved to be a novel method for cancer treatment in vitro, the in vivo therapeutic effects of combination therapy should be further investigated in animal tumor models that could mimic the clinical situation closely for extending it toward the clinical application of combination therapy.
This study was prepared to evaluate the in vivo therapeutic potential of 131I added to doxorubicin treatment in mouse colon cancer model coexpressing MDR1 shRNA and hNIS gene in a single gene construct and to visualize the antitumor effects using nuclear imaging.
Materials and Methods
Mice
The Review board of the university approved the study for the Handling and Use of Animals. Specific pathogen-free 6-week-old male BALB/c nude mice were obtained from SLC.
Construction of vector
The siRNA sequencing targeting MDR1 gene corresponded to the coding region 3491–3509 bp of human MDR1 DNA (Gene Bank: NM_000927). The oligonucleotides with a sequence predicted to induce efficient RNAi of MDR1 (containing sense and antisense sequences linked by a hairpin loop) were synthesized using primers (Forward: 5′-gatccc GGCCTAATGCCGAACACAT CGAA ATGTGTTCGGCATTAGGCC ttttttccaac-3′; Reverse: 5′-tcgattggaaaaaa GGCCTAATGCCGAACACAT TTGG ATGTGTTCGGCATTAGGCC gg-3′). These oligonucleotides were annealed in STE buffer (10 mM Tris pH 8.0, 50 mM NaCl, and 1 mM EDTA) at 94°C for 5 minutes and gradually cooled. The double strand RNA for MDR1 gene was cloned downstream to the human U6 promoter and expression of human NIS gene was induced by CMV promoter in the pRNAT U6 vector (GenScript Co.).
Establishment of stable cells
The human colon cancer HCT-15 cells were cultured in RPMI 1640 (GibcoBRL Co.) culture media containing 10% fetal bovine serum (Hyclone) and 1% antibiotics. HCT-15 cells were grown to 50%–60% confluency and transfected with purified recombinant plasmid using lipofectamine and reagent (Invitrogen) according to manufacturer's guidelines. The transfected cells express MDR1 shRNA, hNIS, and neomycin resistance gene. After treatment with 500 μg/mL G418 (Duchefa Biochemie Co.) for 2 weeks, the stable cell line (referred to as MN61 cell) was established.
RT-PCR analysis
Cells (HCT-15 and MN61 cells) were lysed in Trizol solution (Invitrogen), and RNA was extracted according to manufacturer's instructions. Reverse transcription was performed using the Superscript First-Strand cDNA Synthesis kit (Fermentas Co.). Briefly, 2 μg total RNA was reverse transcribed in a final volume of 20 μL containing 200 ng random hexamer primer, 1 × reverse transcription buffer, 2.5 mM MgCl2, 1 mM dNTP mixture, 10 mM DTT, 20 unit ribonuclease inhibitor, and 50 units of Superscript reverse. The reaction mixture contained 2 μL of cDNA template, 1.5 mM MgCl2, 2.5 units of rTag polymerase, and 20 pM of MDR1 primer (forward: 5′-ATATCAGCAGCCCACATCAT-3′; reverse: 5′-GAAGCACTGGGATGTCCGGT-3′) and NIS primer (forward: 5′-GGGGCTCTCTCACTCATCTACGGAT reverse: AAGCTCTAGATCAGAGGTTTGTCTCCTGC T). GAPDH primer (forward: 5′-GCCAAAAGGGTCATCATCTC-3′; reverse: 5′-GTAGAGGCAGGGA TGATGTTC-3′) was used as an internal control. PCR products were electrophoresed on 1.5% agarose gels, and PCR fragments were visualized using ethidium bromide staining.
99mTc-sestamibi uptake assay
To assess functional expression of P-gp, 99mTc-sestamibi uptake was measured in vitro. Cells were plated at 2 × 105 cells per well in 24-well plates. One day later, the cells were washed once with 2 mL of PBS and then initiated by adding 0.1 mL of PBS containing 370 kBq of 99mTc-sestamibi (Cardiolite; Bristol-Myers Squibb Medical Imaging) per well. After incubation for 30 minutes at 37°C in 5% CO2, the cells were washed twice with ice-cold PBS. Cells were lysed with 2% SDS, and the radioactivity was measured with a well γ-counter (Packard). The radioactivity of cells was normalized with total proteins, which was measured using bicinchoninic acid Protein Assay Kit (Pierce Chemical Co.), and cellular uptake of 99mTc- sestamibi was calculated as cpm/mg protein.
In vitro 125I uptake assay
Cells were plated at 2 × 105cells per well in 24-well plates. One day later, the cells were washed once with 1 mL of HBSS 1 day after plating, and then the iodide uptake assay was initiated by the addition of 0.5 mL HBSS containing 3.7 kBq carrier-free 125I and 10 μM sodium iodide per well, to yield a specific activity of 740 MBq/mmol (20 mCi/mmol). After incubation with 125I for 30 minutes at 37 °C, the cells were washed twice with cold HBSS. The radioactivity of cells was quantified with a well γ-counter. The radioactivity of cells was normalized for total protein, and 125I uptake was expressed as cpm/mg protein. For the inhibition study, radioiodine uptake was measured after pretreatment with 50 μM potassium perchlorate for 1 hour, and cellular radioactivity was measured. 12,20
In vitro clonogenic assay
For 131I treatment in vitro, 5 × 106 cells MN61 and HCT-15 cells per T75-culture flask were seeded in a tissue culture flask. Cells were washed once with 5 mL of HBSS and incubated with 5 mL of HBSS (control) or 5 mL of HBSS containing 18.5 MBq of 131I. After incubating with 131I for 7 hours, cells were washed twice with cold HBSS, trypsinized, and counted. One thousand cells were plated in triplicate in 6-well plates to which a doxorubicin dilution (25 nM) was added in triplicate, and incubated for 1 week at 37°C. Cells were then washed once with PBS and stained with a crystal violet solution (for 250 mL, 0.5 g of crystal violet, 25 mL of 40% formaldehyde, 50 mL of ethanol, and 175 mL of water). Colonies of more than 30 cells were counted, and all values were adjusted for plating efficiency. Cell survival was expressed as the percentage of colonies relative to that in the untreated control.
In vivo imaging with 123I and 99mTc-sestamibi
1 × 107 MN61 and HCT-15 were subcutaneously inoculated to left and right thighs of nude Balb/c mice, respectively. When tumors had reached an appropriate volume ≤100 mm3, scintigraphic imaging was obtained. 18.5 MBq 99mTc-sestamibi was intravenously injected into tumor-bearing mice to evaluate the MDR1 shRNA-mediated effects. Images were obtained at 60 minutes after 99mTc-sestamibi injection.
At 24 hours after 99mTc-sestamibi imaging acquisition, 18.5 MBq 123I was intravenously administered for hNIS-mediated imaging, and images were obtained at 90 minutes after 18.5 MBq 123I injection. Images were acquired using a γ camera equipped with a low-energy, pinhole collimator (Infinia, GE).
In vivo therapy with 131I and doxorubicin
When tumors reached an appropriate volume ≤100 mm3, MN61 and HCT-15 tumor-bearing mice were sub-divided into four groups (saline as a control group, doxorubicin or 131I as a single therapy group, and both doxorubicin and 131I as a combination therapy group, 10–12 mice per group). For 131I therapy, mice received a low-iodine diet with T4 supplementation in drinking water for 6 days to maximize radioiodine uptake in tumors and to reduce iodide uptake by the thyroid gland. Control group received intravenous injection of saline. Doxorubicin therapy group (0.5 mg/kg) was intraperitoneally administered at the interval of 3 days for 30 days, and 131I therapy group (55.5 MBq per mouse) received an intravenous injection of 55.5 MBq 131I. Combination therapy group received both doxorubicin (at the interval of 3 days for 42 days) and 131I (55.5 MBq per mouse, single injection). Tumor volume was estimated using the formula (L × W × H × 0.52) for 6 weeks. To monitor the therapeutic effects, 18.5 MBq 99mTc-pertechneate was intravenously administered to tumor-bearing mice, and scintigraphic imaging was acquired at 6 weeks after tumor challenge using a γ camera.
Statistical analysis
Data are presented as the mean ± standard deviation. Intergroup differences were assayed using the two-tailed Student's t-test. p-Values of <0.05 were considered statistically significant.
Results
RT-PCR analysis for MDR1 and hNIS gene expression
RT-PCR analysis revealed that the MDR1 gene expression was markedly reduced in the MN61 cells as compared with the parental cells, and the hNIS gene expression was observed in the MN61 cells but not in the parental HCT-15 cells (Fig. 1).
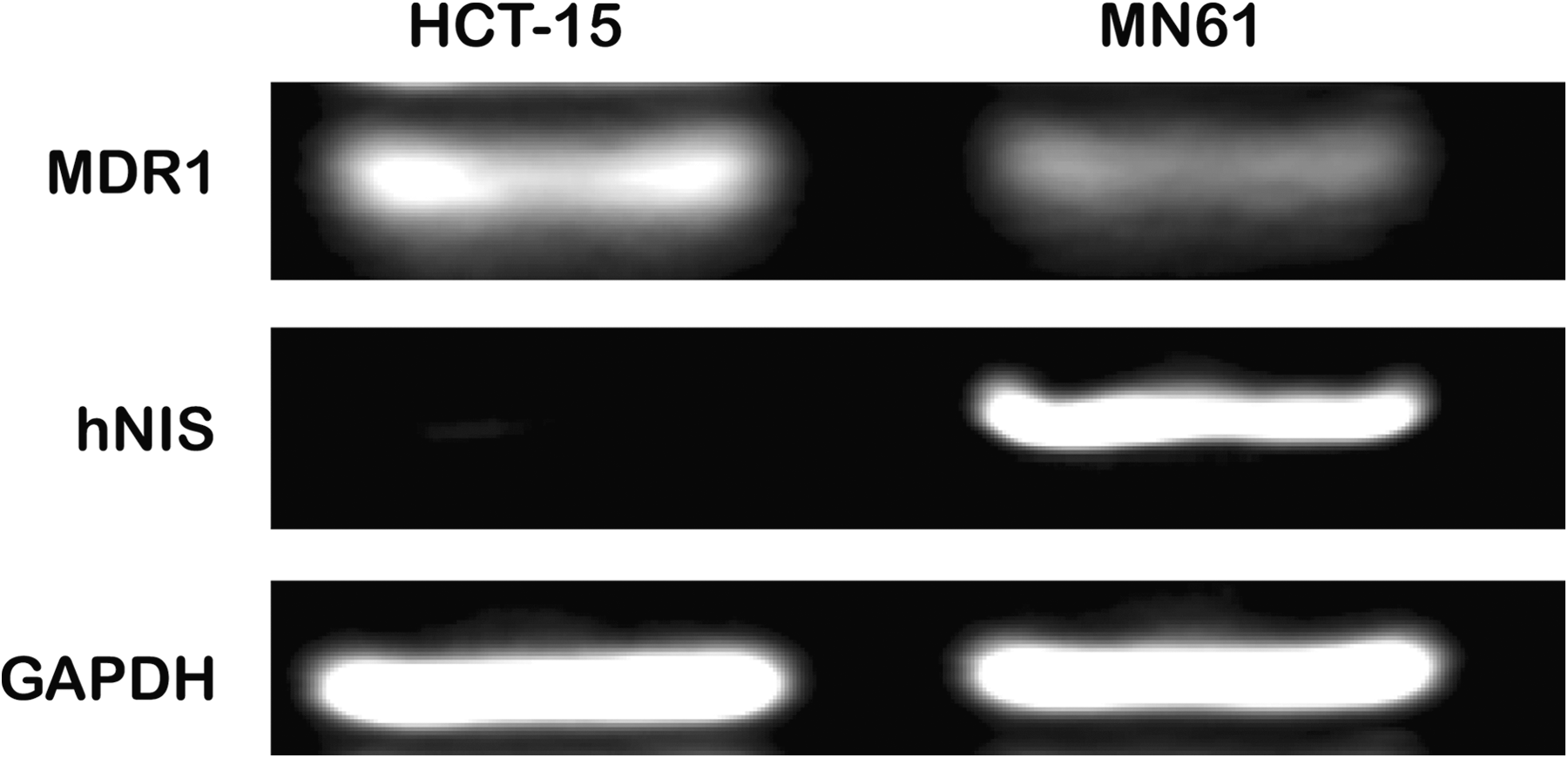
Suppression of MDR1 mRNA and hNIS gene expression in MN61 cells. Total RNA was isolated from HCT-15 and MN61 cells, and the MDR1 and hNIS gene was amplified and determined by RT-PCR analysis. GAPDH was used as an internal control. PCR products were electrophoresed on agarose gels and then visualized by ethidium bromide staining. hNIS, human sodium iodide symporter; MDR, multidrug resistance.
99mTc-sestamibi and 125I uptake in the MN61 cells coexpressing both MDR1 shRNA and hNIS gene
99mTc sestamibi uptake was significantly higher in MN61 cells than in parental cells (Fig. 2A, MN61 and parental HCT-15 cells, 1500 ± 8.1 and 805 ± 100.0 cpm/mg protein, respectively, p < 0.05). 125I uptake of MN61 cells was 1100 ± 80.1cpm/mg protein, which is 157-fold higher than that of parental HCT-15 cells (7 ± 0.2 cpm/mg protein) (Fig. 2B, p < 0.05). 125I uptake in hNIS expressing cells was almost completely inhibited by treatment with perchlorate, a competitive NIS inhibitor.
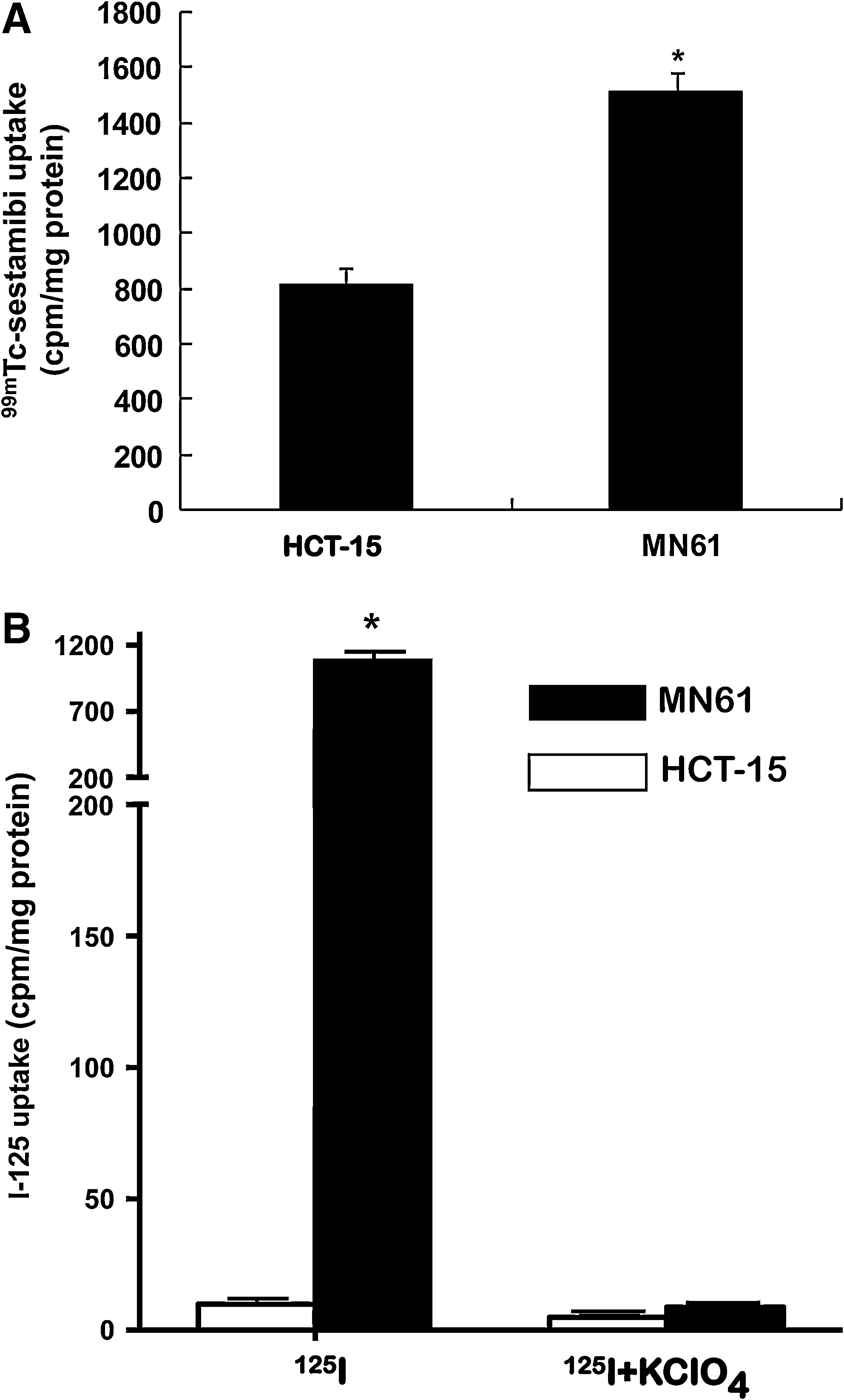
Functional uptake of 99mTc-sestamibi and 125I in MN61.
Combination therapy with 131I and doxorubicin in vitro
In the presence of 25 nM doxorubicin, the cell survival was comparatively lower in MN61 cells than in parental HCT-15 cells (Fig. 3). When 18.5 MBq 131I was treated, cell survival of MN61 cells or parental HCT-15 cells was 35 ± 4.3% and 93 ± 4.8%, respectively (p < 0.05). The combination of 25 nM doxorubicin and 18.5 MBq 131I resulted in much more decrease in cell survival of MN61 cells than parental HCT-15 cells (MN61 and HCT-15, 12 ± 5% and 73 ± 7.3%, p < 0.01)
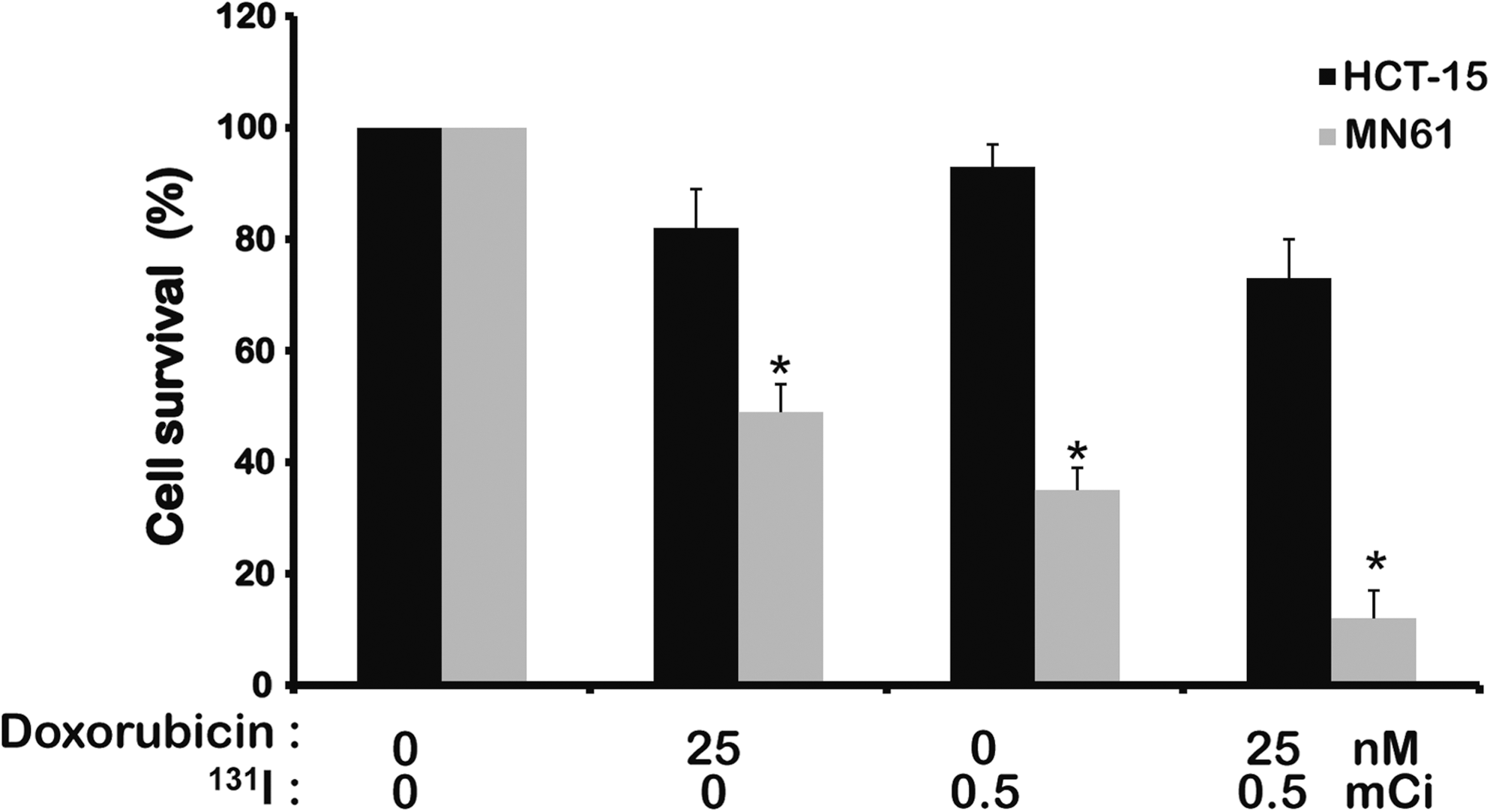
Enhanced cell growth inhibition by combination therapy with 131I and doxorubicin in vitro. For 131I treatment in vitro, 5 × 106 cells MN61 and HCT-15 cells per T75-culture flask were seeded. Cells were washed once with 5 mL of HBSS and incubated with 5 mL of HBSS (control) or 5 mL of HBSS containing 18.5 MBq of 131I. After incubating with 131I for 7 hours, cells were washed twice with cold HBSS, trypsinized, and counted. One thousand cells were plated in triplicate in six-well plates to which a doxorubicin dilution (25 nM) was added in triplicate and incubated for 1 week at 37°C. Cells were then washed once with PBS and stained with a crystal violet solution. Colonies of more than 30 cells were counted, and cell survival was expressed as the percentage of colonies relative to that in the untreated control. Results were expressed as mean ± SD of three experiments. *p < 0.05, statistical significance value.
In vivo 99mTc- sestamibi and 123I imaging of tumor-bearing mice
As shown in Figure 4, scintigraphic imaging demonstrated the higher uptake of 99mTc- sestamibi in MN61 tumor than parental HCT-15 tumor, which is 2.2-fold higher by quantitative analysis (p < 0.05). γ Camera imaging with 123I demonstrated about a 15-fold higher uptake in MN61 tumor than parental HCT-15 tumor (p < 0.01).
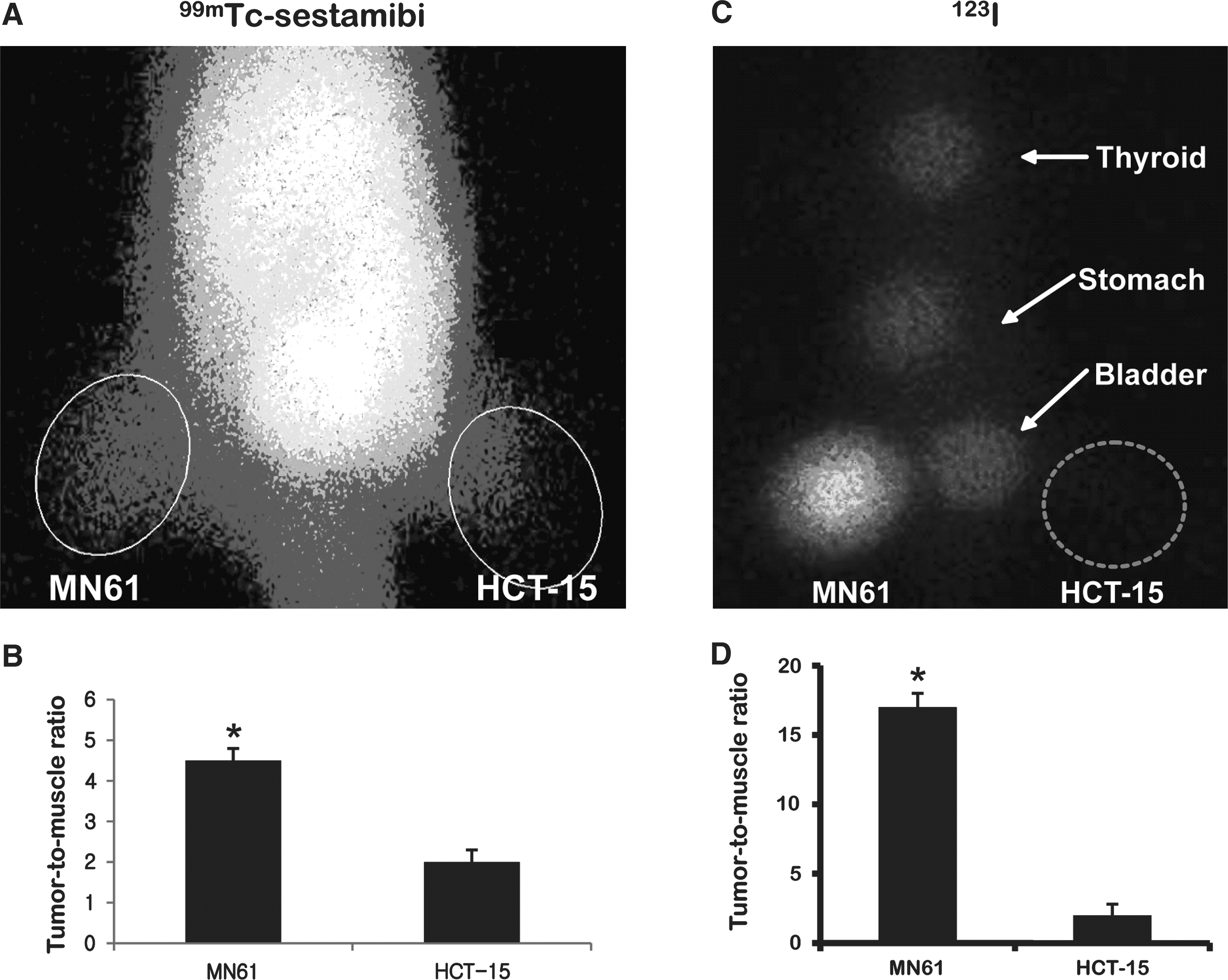
Scintigraphic imaging with 99mTc-sestamibi and 123I in vivo. 1 × 107 MN61 and HCT-15 were subcutaneously inoculated to left thighs and right thighs of nude mice, respectively. When tumors had reached an appropriate volume ≤100 mm3, scintigraphic imaging was acquired.
In vivo therapeutic effects of combination treatment and monitoring of tumor regression
HCT-15 tumor (right thighs) in four groups showed similar growth rate, and aggressive tumoral growth continued for 6 weeks (Fig. 5B). MN61 tumor (left thighs) of doxorubicin therapy group showed slight retardation of growth, but nontreated MN61 tumors (control) grew aggressively (nontreated MN61 tumor and doxorubicin treated MN61 tumor, 200 ± 45% and 95 ± 12% of initial tumor volume, p < 0.05). MN61 tumors in combination therapy group (doxorubicin + 131I) as well as 131I therapy group showed a rapid decrease of tumor volume from 0.5 weeks after therapy initiation, and then tumors in the two groups were rarely detected at 6 weeks with visual inspection and palpation (Fig. 5C, D). Particularly, the combination therapy resulted in faster tumor regression than 131I therapy alone (Fig. 5D, p < 0.05, at 1 week after combination therapy).

In vivo enhanced antitumor effects of combination with doxorubicin and 131I.
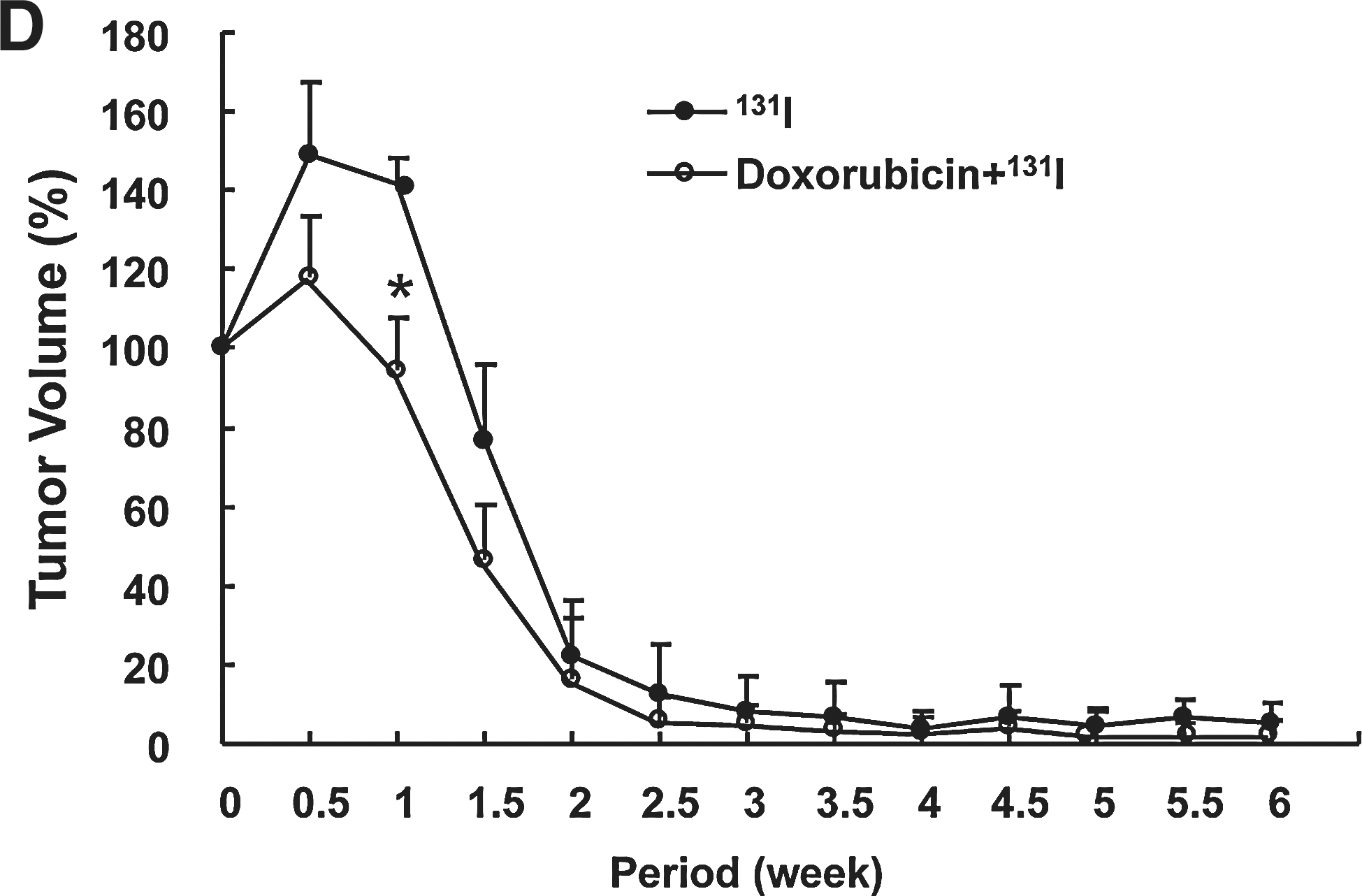
Scintigraphic imaging to monitor the therapeutic response of combination therapy was acquired at 6 weeks after in vivo therapy using 99mTc-pertechnetate. Scintigraphic imaging revealed an intensive accumulation of 99mTc-pertechneate in MN61 tumor of both control group and doxorubicin treatment group (Fig. 6). The accumulation of radiotracer was comparatively lower in MN61 tumor of doxorubicin group than control group. 131I therapy revealed slightly decreased tracer accumulation in tumor region, but radioactivity was rarely shown in MN61 tumor of combination therapy group.

In vivo monitoring of antitumor effects. To monitor the therapeutic effects, scintigraphic imaging was acquired at 6 weeks after tumor challenge using 99mTc-pertechneate. 18.5 MBq 99mTc-pertechneate was intravenously administered to tumor-bearing mice. Images were acquired using a gamma camera equipped with a pinhole collimator. The physiologic iodide uptake was shown in thyroid, stomach, and urinary bladder. Experiments were performed in duplicate.
Discussion
The successful modulation of MDR in cancer overexpressing MDR1 gene is an essential concept to maximize therapeutic effects of anticancer drug therapy. Recently, several researchers have adopted the RNAi technology to modulate the MDR1 gene in MDR cancer, particularly emphasizing that siRNA for MDR1 gene increased drug sensitivity of MDR cancer cells to anticancer drugs such as vinblastine and doxorubicin. 16,17 RNAi has been found to be a powerful tool for post-transcriptional gene silencing. 14,15 The degradation of long double-stranded RNA to siRNA is mediated by double-stranded RNA-specific enzymes, RNase III dicer. siRNA is incorporated into a nuclease complex known as the RNA-induced silencing complex, in which unwinding of the duplex siRNA takes place. The antisense strand binds to targeting mRNA in a highly sequence-specific manner, which is then endonucleolytically cleaved and degraded. Application of siRNA for MDR1 gene to MDR cancer can provide an opportunity to use lesser dosage of anticancer drug and subsequently maximize the therapeutic effects of chemotherapy. However, it has been noted that single therapy using RNAi-based technology for cancer has frequently resulted in unsuccessful therapeutic outcomes for aggressive tumor. As seen in our in vivo treatment trial, although single therapy with MDR1 shRNA for 6 weeks resulted in slight retardation of tumor growth, it could not induce complete inhibition of tumor growth. These findings also suggest that RNAi-based gene therapy alone is not sufficient to achieve the control of aggressive or bulky tumors necessitating combination therapy with other therapeutic strategies that are required to overcome this weak therapeutic response of RNAi-based gene therapy.
hNIS radioiodine gene therapy has shown strong inhibitory effects of tumor growth in various cancer models. It has the potential to become an attractive therapeutic gene due to the possibility of delivery of hNIS gene to cancer cells under the control of tissue-specific promoter. Several reports have already revealed the antitumor effects by hNIS radioiodine gene therapy in several cancer models (prostate cancer, hepatocelluar carcinoma, and breast cancer) using adenovirus system expressing hNIS gene under the tissue-specific promoter. 21 –23 These advantages of hNIS gene would satisfy some critical hazards of radiation therapy in the management of patients with cancer.
Based on several interesting sequential reports, the following mechanisms were postulated: (1) Delivery of hNIS gene into cancer cells can induce the accumulation of various therapeutic radionuclides in hNIS expressing cancer cells, and therapeutic radionuclide accumulation causes a strong apoptosis of cancer cells and it subsequently inhibits tumor growth without damaging the normal cells; (2) Combined MDR1 shRNA and anticancer drug results in retardation of tumor growth at lower doses of anticancer agents; (3) Combination hNIS radioiodine gene therapy and MDR1 shRNA-mediated chemotherapy could provide an opportunity to treat aggressive cancer model (do not respond to single therapy), additionally with potentially minimal adverse effects on normal tissue.
In this study, to investigate the therapeutic potential of combined hNIS radioiodine gene therapy and shRNA for MDR1 (MDR1 shRNA), an attempt was made at establishing stable human colon cancer cells that express both a short hairpin RNA for MDR1 gene and hNIS gene with a single construct (MN61 cell), and the therapeutic effects of combination therapy in animal MDR tumor model were compared with single therapy. Since HCT-15 colon cancer cell line expressed MDR1/P-gp, this was observed by FACS analysis using a primary P-gp antibody (data not shown); hence, this cell line was selected. A specific sequence of MDR1 shRNA was adopted that had proved to be effective in downregulating the MDR1 gene in other groups. 16,24,25 MDR1 shRNA can successfully downregulate the gene expression of MDR1 gene and the ablation of MDR1 gene in MN61 that renders an increased sensitivity of doxorubicin (Figs. 1 and 3). Also, the modulation of MDR1 gene by MDR1 shRNA was successfully assessed with in vivo scintigraphic imaging, showing that 99mTc-sestamibi is accumulated more in MN61 tumor than parental tumor (Fig. 4). 99mTc-sestamibi is a lipophilic cationic radiopharmaceutical for myocardial and cancer imaging performed with a gamma camera, and it is believed to be transported as a common transport substrate by P-gp. 26 The decreased intracellular level of substrate is due to enhanced efflux by membrane transporters, and reduced MIBI uptake in an MDR-expressing tumor is related to P-gp. hNIS gene expression and 131I treatment in HCT-15 in vitro induce the cellular uptake of radiotracer and result in strong cytotoxic effects (Figs. 2B and 3). Scintigraphic imaging revealed a higher uptake of radiotracer in MN61 tumors but not in parental tumors (Fig. 4C, D), and 131I or doxorubicin treatment alone induces stronger tumor growth inhibition than control (Figs. 5C and 6, nontreatment group in MN61), suggesting that MDR1 shRNA enhances the sensitivity of doxorubicin and that intensive accumulation of 131I by hNIS induces strong antitumor effects. Doxorubicin is known to be an activator of p53 pathway that induces the gene expression of various proteins related to cell cycle and apoptosis; it was expected that doxorubicin could accelerate the strong apoptosis induced by hNIS radioiodine therapy. Interestingly, a more rapid reduction of tumor volume was observed in combination therapy group than 131I therapy group (Fig. 5D).
Although combined hNIS radioiodine gene therapy and MDR1 shRNA showed strong antitumor effects, two important problems to be overcome are anticipated for expanding this current combination therapy to clinical trial. (1) Since a higher dose of 131I (1.5mCi per mouse) is used to potentiate therapeutic effects in animal cancer model for hNIS radioiodine gene therapy, proper dosage of 131I that can be used for the application in a clinical setting should be investigated. (2) Stable colon cancer cell line was used that coexpressed hNIS and MDR1 shRNA, and this is an artificial setting for clinical cancer therapy. Thus, improved therapeutic gene delivery to cancer cells in vivo should be investigated, such as using adenovirus system under the control of tissue-specific promoter.
Conclusions
This study demonstrated the therapeutic potential of combined 131I and doxorubicin therapy in animal colon cancer model and monitored not only the reversal of MDR1 by MDR1 shRNA but also tumor growth inhibition using NIS gene and scintigarphic imaging. Although many hurdles still remain to be resolved for further application procedures, current study suggests that hNIS radioiodine gene therapy and RNAi-based gene therapy in a single gene construct can potentially be used as new therapeutic strategies for treating a patient with cancer who does not respond to conventional therapy.
Footnotes
Acknowledgments
This work was supported by Ministry of Education, Science, and Technology and National Research Foundation of Korea through Nuclear R&D Program, 2010 (No. 001-7540); the Korean Ministry of Education, Science, and Technology (The Regional Core Research Program/Medical Convergence Technology Development Consortium for Antiaging and Well-being); and the Basic Atomic Energy Research Institute (BAERI, 2010-0017515) and BK21 (2009). The authors declare no conflict of interest.
Disclosure Statement
No competing financial interests exist.