
Abstract
Cyclophosphamide (CTX) is a commonly used chemotherapeutic agent for breast cancer. However, chemoresistance remains a common clinical problem. Osteopontin (OPN) has been shown to induce chemoresistance by inhibiting apoptosis; p38 MAPK pathway has been reported to be involved in chemotherapy-induced apoptosis. Thus, this study investigated whether OPN knockdown would chemosensitize MDA-MB-231 cells to CTX by enhancing apoptosis through activating p38 MAPK pathway. MDA-MB-231 cells were transfected with OPN stable siRNA plasmid, and it was found that OPN knockdown chemosensitized MDA-MB-231 cells to CTX, which is dependent on p38 MAPK pathway activation. Moreover, results showed that each of OPN knockdown and CTX could induce apoptosis through activating p38 MAPK pathway and that OPN knockdown and CTX could induce enhanced apoptosis through activating p38 MAPK pathway synergistically. Therefore, this study concludes that OPN knockdown chemosensitizes MDA-MB-231 cells to CTX by enhancing apoptosis through activating p38 MAPK pathway.
Introduction
Breast cancer, the most common malignant tumor worldwide, is one of the leading causes of cancer-related deaths in women. 1 The last two decades has witnessed a decline in breast cancer mortality rate, which is partly attributed to adjuvant therapy. 2 Among various agents, cyclophosphamide (CTX) exerts its cytotoxicity by damaging DNA. However, chemoresistance, primarily existed or acquired, remains a common clinical problem in the treatment of breast cancer.
Osteopontin (OPN), highly expressed in several types of cancers, 3 –5 has been recently shown to induce chemoresistance to DNA-damaging chemotherapeutic drugs by inhibiting apoptosis. 6,7 Moreover, knockdown of OPN has been reported to chemosensitize hepatocellular carcinoma cells to chemotherapeutic agents through apoptosis. However, whether knockdown of OPN would chemosensitize MDA-MB-231 cells to CTX by enhancing apoptosis has not been investigated. In addition, a growing body of evidence revealed that p38 MAPK pathway has proapoptotic activity in cells exposed to various agents 8,9 and that several chemotherapeutic agents induce apoptosis via activating p38 MAPK pathway, 10,11 justifying a critical role of p38 MAPK pathway in apoptosis-mediated chemosensitivity. Thus, the present study hypothesizes that knockdown of OPN induces chemosensitivity to CTX by enhancing apoptosis through activating p38 MAPK pathway.
In the present study, in vitro experiments were conducted to determine whether knockdown of OPN by RNA interference would chemosensitize MDA-MB-231 cells to CTX by enhancing apoptosis through activating p38 MAPK pathway.
Materials and Methods
Cell culture
MDA-MB-231 cells, obtained from Heilongjiang Cancer Institute, Harbin, China, were cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% fetal bovine serum (FBS; Invitrogen), 100 U/mL penicillin, and 100 μg/mL streptomycin at 37°C in a humidified atmosphere of 5% CO2 and 95% air.
Transfection of stable siRNA plasmids
For stable RNA interference, hairpin siRNA sequences that contain both sense and antisense siRNA inserts were employed. Three pairs of hairpin siRNA inserts targeting human OPN (NM_000582) were employed: one (OPN#1) that was previously described 12 was sense 5′-ACGAGUCAGCUGGAUGACC-3′, antisense 5′-GGUCAUCCAGCUGACUCGU-3′; one (OPN#2) that was designed was sense 5′-GUUUCGCAGACCUGACAUC-3′, antisense 5′-GAUGUCAGGUCUGCGAAAC-3′; and another (OPN#3) that was also designed was sense 5′-AGCGAGGAGUUGAAUGGUG-3′, antisense 5′-CACCAUUCAACUCCUCGCU-3′. In addition, a pair of hairpin siRNA inserts (sense, 5′–3′ AUUGCGUUCGCAGUAAUCU; antisense, 5′–3′ AGAUUACUGCGAACGCAAU) that had no significant homology to any human mRNA was used as a control. The structure of a hairpin siRNA consists of two inverted sense and antisense inserts separated by a short spacer sequence (UUCAAGAGA) as the loop and ends with a string of U's. These sequences were chemically synthesized by Shanghai Gene Chem Co., Ltd. Plasmids were constructed by inserting these hairpin siRNA sequences into pSilencer 2.1-U6 neo vectors (Ambion). Hence, pSilencer vectors contained either OPN siRNA sequences or negative control siRNA sequences. Plasmids were thereafter amplified and purified using QIAfilter Plasmid Maxi Kit (Qiagen) and verified by sequence analysis.
MDA-MB-231 cells were divided into three groups: OPN RNAi (pS-OPN), control RNAi (pS-CO), and wild type (WT). Afterward, pS-OPN, pS-CO, and WT were incubated with OPN siRNA plasmid-Lipofectamine 2000 complex, control siRNA plasmid-Lipofectamine 2000 complex, or Lipofectamine 2000 alone in the absence of plasmid, respectively. MDA-MB-231 cell transfection was performed with Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. Forty-eight (48) hours after transfection, the cells were incubated in DMEM supplemented with G418 (400 μg/mL). After 2 weeks of growth in G418-containing DMEM, G418-resistant cells that stably expressed OPN siRNA (pS-OPN) or control siRNA (pS-CO) were selected and used to perform later experiments.
Protein preparation and western blot analysis
Total cell cytoplasmic extracts were prepared as previously described. 13 Then, total protein concentration was determined by BCA Protein Assay kit (Pierce). Twenty (20) micrograms of each soluble protein sample was separated by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to presoaked nitrocellulose membranes that were blocked with 3% bovine serum albumin for 2 hours. Afterward, the membranes were incubated overnight with primary mouse antibodies against human OPN (Santa Cruz), β-actin (Abcam), p38 (Santa Cruz), or anti-phospho-p38 (Sigma-Aldrich) at 4°C. Membranes incubated with primary antibodies were then exposed to horseradish peroxidase-conjugated anti-mouse IgG for 2 hours at room temperature. Proteins were detected by enhanced chemiluminescence. Bands were then scanned into Adobe Photoshop 7.0 and relative absorbance was calculated in Tina 2.0 in reference to β-actin. All experiments were run in triplicate.
Quantitative real-time polymerase chain reaction
Cells were harvested and total RNA was extracted using RNeasy Mini Kit (Qiagen) according to the manufacturer's instructions. First-strand cDNA was synthesized for each RNA sample using Superscript II RT Kit (Gibco-BRL). Real-time polymerase chain reaction (PCR) using SYBR Green I (Invitrogen) was performed on an ABI Prism 7000 Sequence Detection System (Applied Biosystem). Primers for OPN are as follows: forward, 5′–3′ CTGACATCCAGTACCCTGATGC; reverse, 5′–3′ GGCCTTGTATGCACCATTCA. The primer sequences for GAPDH are as follows: forward, 5′–3′ AAGATCATCAGCAATGCCTCC; reverse, 5′–3′ TGGACTGTGGTCATGCCTT. Real-time PCR was performed as follows: 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 60 seconds. All samples were run in triplicate. The quantitative real-time PCR results for OPN that were normalized to the amount of GAPDH mRNA from the same sample were expressed as fold increase or decrease.
MTT assay
After G418-resistant cell selection, cells were harvested and inoculated at a density of 5000 cells/well onto 96-well plates in 100 μL of DMEM supplemented with 10% FBS and were then cultured for 24, 48, 72, and 96 hours. Older medium was replaced by 100 μL of fresh serum-free DMEM every 24 hours. Then, 10 μL of MTT reagent (5 mg/mL; Sigma) was added into each well and the cells were incubated for additional 4 hours in a CO2 incubator (NAPCO). Afterward, the medium was removed and 100 μL of dimethyl sulfoxide was added to dissolve formazan. Results were assessed on a microplate reader by measuring the absorbance at a wavelength of 540 nm and a reference wavelength of 630 nm. Relative cell numbers were calculated as the ratio of optical density (OD) between test samples and control samples (WT). All experiments were performed in triplicate.
Cell cycle distribution analysis
Cells were plated in 60-mm dishes and allowed to grow for 48 hours. The cells were collected by trypsinization. Then, the cells were washed in PBS and fixed in 70% ethanol. After treatment with RNAase (200 μg/mL) for 30 minutes at 37°C, 50 μg/mL propidium iodide was added. Afterward, DNA content was measured through a FACScan flow cytometry (Becton Dickinson). The experiments were carried out in triplicate.
TUNEL assay
Cells were harvested and inoculated at a density of 5000 cells/well onto 96-well plates and incubated in 100 μL DMEM supplemented with 10% FBS for 48 hours. Afterward, the cells were mounted on eight-well chamber slides of 1000 cells per 100 μL and fixed by 4% paraformaldehyde in PBS for 1 hour. Then, apoptosis was analyzed by TUNEL assay (Roche) according to the manufacturer's protocols. Apoptotic cells were identified as those that were stained yellow-brown with pyknotic nuclei under a microscope. Apoptotic cells were counted and results were calculated as a mean percentage of apoptotic cells counted in five fields per chamber.
Chemosensitivity assay
After cell selection, cells were harvested and inoculated at a density of 5000 cells/well onto 96-well plates and incubated in 100 μL fresh serum-free DMEM containing various concentrations of CTX (0.01, 0.1, 1, 10, and 50 mM) for 48 hours. MTT assay was performed as mentioned earlier to evaluate chemosensitivity to CTX.
Statistical analysis
Data were expressed as means ± SD of triplicate samples in single experiments. Student's t-test was used to compare between two groups. All the statistical analyses were performed on SPSS 16.0 and a value of p < 0.05 was considered statistically significant.
Results
RNAi decreased OPN protein and mRNA expressions
Detection of OPN protein in cytoplasmic extracts by western blot analysis showed that OPN#1 was successful in suppressing OPN protein expression, whereas OPN#2 and OPN#3 were unsuccessful (Fig. 1A). Statistically, with reference to WT, the expression of OPN protein was significantly lower in pS-OPN (OPN#1) than in WT (p = 1.1 × 10−5 < 0.05), but pS-OPN (OPN#2), pS-OPN (OPN#3), and pS-CO were similar to WT (Fig. 1B). Moreover, similar results were found for OPN mRNA expression (Fig. 1C). These findings indicate that OPN knockdown by OPN#1 was successful and effective. Thus, in later experiments, only pS-OPN (OPN#1) was used.
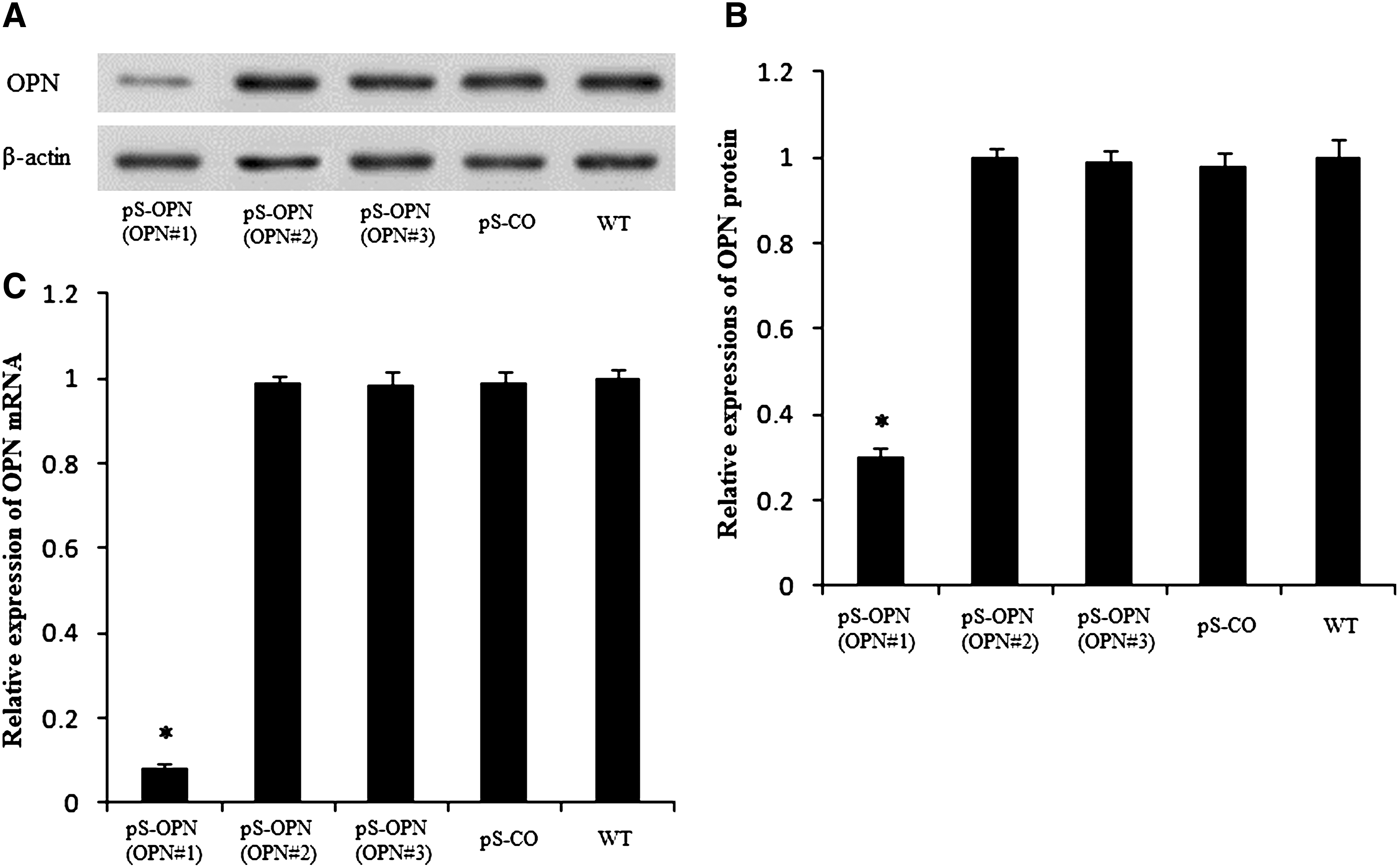
RNAi decreased OPN protein and mRNA expressions.
Effects of CTX on MDA-MB-231 cells
The effects of CTX on MDA-MB-231 cells were studied. To exclude the effects of OPN knockdown, the WT group was specifically studied. First, MDA-MB-231 cells were exposed to various concentrations of CTX (0.01, 0.1, 1, 10, and 50 mM) and it was found that MDA-MB-231 cells were sensitive to CTX (Fig. 2A). Then, whether this effect was due to cell apoptosis was investigated. TUNEL assay was performed and it was found that CTX was able to induce cell apoptosis (Fig. 2B). In search of a target responsible for apoptosis induced by CTX, western blot analysis of p38 MAPK (p38) and the phosphorylation state of p38 MAPK (p-p38) was performed, and it was found that CTX was able to activate p38 MAPK pathway (Fig. 2C). Then, it was curious to know whether the activation of p38 MAPK pathway is essential for CTX-induced apoptosis. So, the activity of p38 MAPK pathway was inhibited using SB202190, a p38-specific inhibitor, and it was found that SB202190 was able to inhibit p38 MAPK activity (Fig. 2C) and that the inhibition of p38 MAPK activity was able to attenuate the CTX-induced apoptosis (Fig. 2B), indicating that CTX induces apoptosis through activating p38 MAPK. Moreover, results showed that attenuation of CTX-induced apoptosis by SB202190 resulted in attenuation of CTX-induced toxicity (Fig. 2A). Therefore, these findings suggested that CTX exerts its toxic effects on MDA-MB-231 cells by inducing cell apoptosis through activating p38 MAPK pathway.

Effects of CTX on MDA-MB-231 cells.
Chemosensitivity assay
To study whether OPN knockdown would chemosensitize MDA-MB-231 cells to CTX, pS-OPN, pS-CO, and WT were exposed to various concentrations of CTX for 48 hours (0.01, 0.1, 1, 10, and 50 mM). As shown in Figure 3A, pS-OPN was significantly more sensitive to CTX than pS-CO and WT, indicating that OPN knockdown was able to chemosensitize MDA-MB-231 cells to CTX.
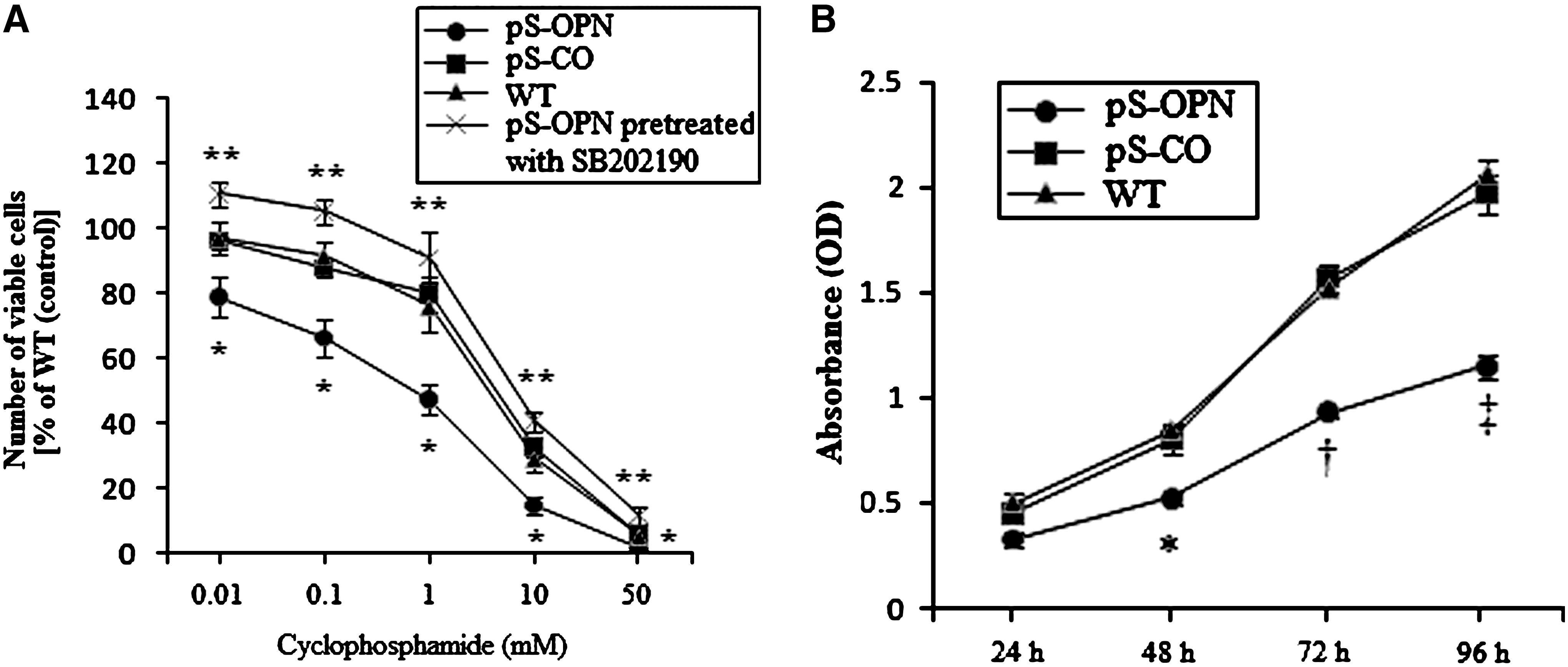
Effects of OPN knockdown on MDA-MB-231 cells.
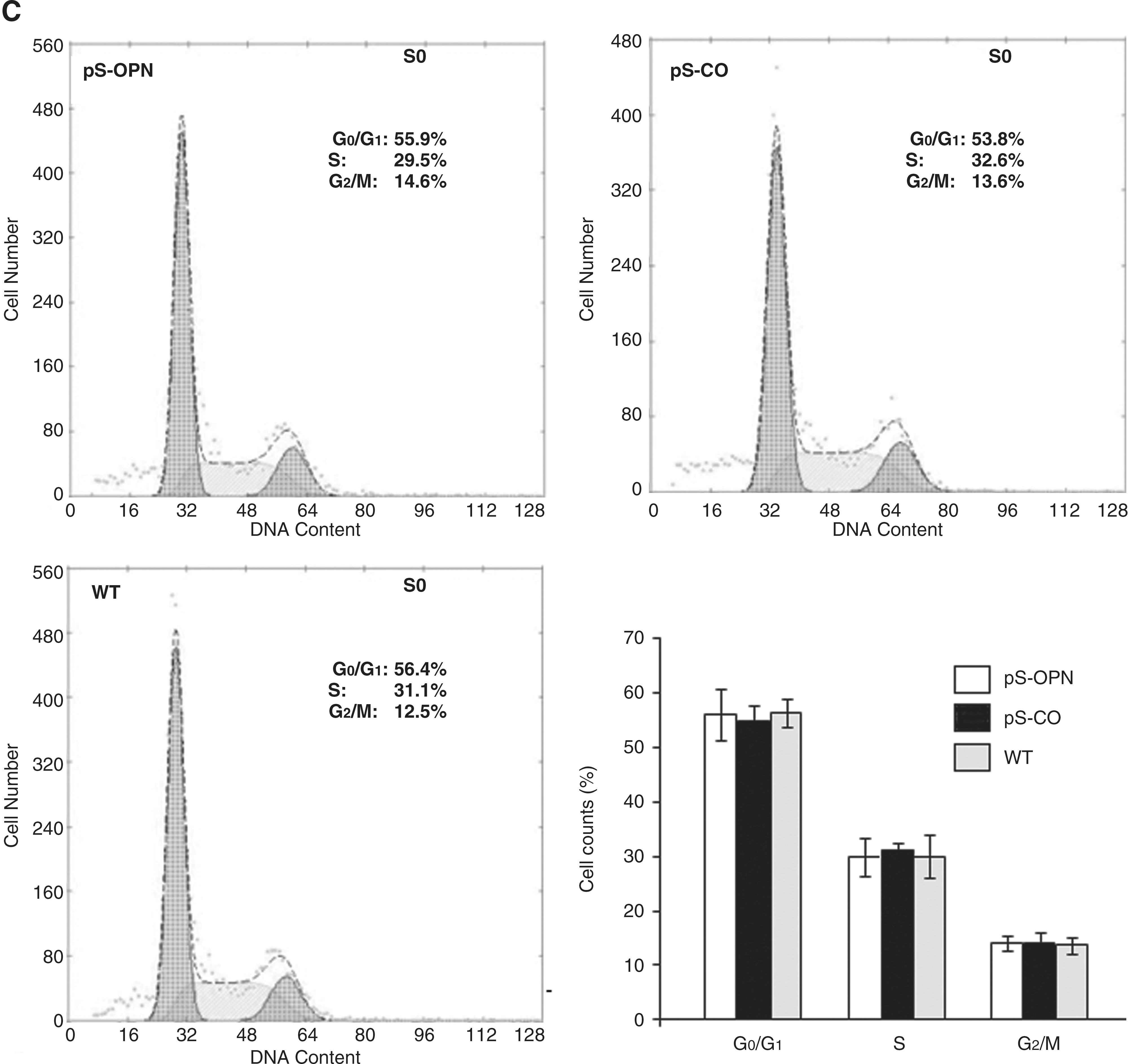
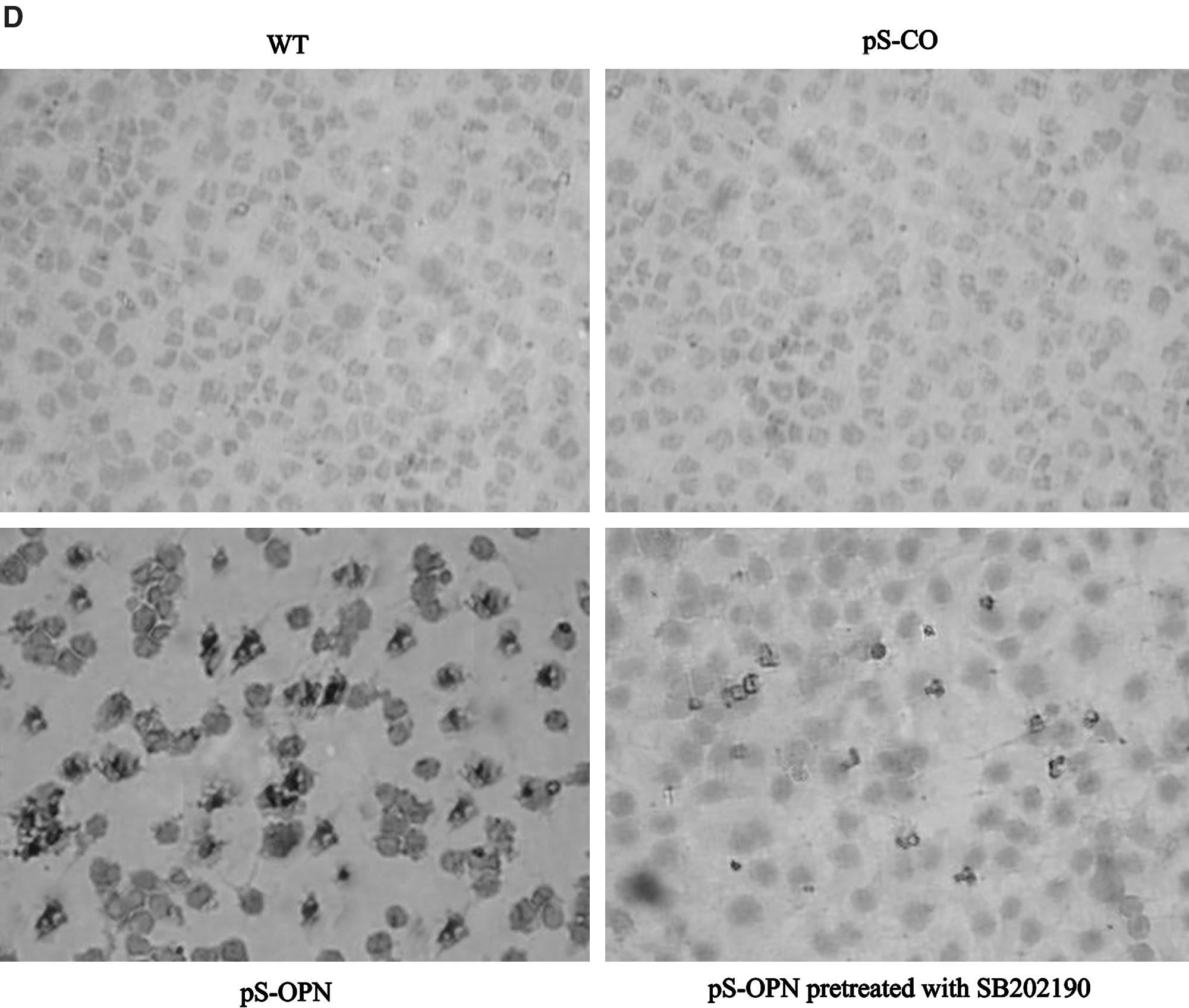

To explore the mechanisms wherein OPN knockdown chemosensitizes MDA-MB-231 cells to CTX, the effects of OPN knockdown on MDA-MB-231 cells were investigated. Results showed that the cell proliferation rate of pS-OPN was significantly lower than those of pS-CO and WT (Fig. 3B), indicating that knockdown of OPN inhibits cell proliferation. In addition, cell cycle distribution analysis and TUNEL assay were performed to investigate whether the inhibition of cell proliferation was due to cell cycle arrest or apoptosis. It was found that OPN knockdown did not influence cell cycle distribution (Fig. 3C) but was able to induce cell apoptosis (Fig. 3D. E), indicating that OPN knockdown inhibited cell proliferation through apoptosis rather than through cell cycle arrest. Results showed that both p38 and p-p38 expressions were higher in pS-OPN than in pS-CO and WT (Fig. 3F), indicating that OPN knockdown was able to activate p38 MAPK pathway. Moreover, it was found that SB202190 was able to inhibit p38 MAPK activity (Fig. 3F) and that inhibition of p38 MAPK activation by SB202190 was able to attenuate OPN knockdown-induced apoptosis (Fig. 3D. E), indicating that p38 MAPK activation was essential for OPN knockdown-induced apoptosis. These results suggest that OPN knockdown was able to induce cell apoptosis through activating p38 MAPK pathway.
As mentioned earlier, both CTX and OPN knockdowns were able to induce cell apoptosis through activating p38 MAPK pathway. It was curious to know whether this chemosensitive effect was due to their synergism to induce cell apoptosis. It was found that there were significantly more apoptotic cells in pS-OPN that was exposed to CTX than in pS-CO and WT that were also exposed to CTX, or in pS-OPN that was not exposed to CTX (Fig. 4A), indicating that OPN knockdown and CTX were able to induce enhanced apoptosis synergistically. Moreover, results showed that OPN knockdown and CTX induced enhanced activation of p38 MAPK pathway synergistically (Fig. 4B). To investigate whether the activation of p38 MAPK pathway was essential for enhanced apoptosis and chemosensitivity, pS-OPN cells were pretreated with SB202190. Results showed that SB202190 can inhibit p38 MAPK activity (Fig. 4B). Moreover, it was found that the inhibition of p38 MAPK activity was able to attenuate enhanced apoptosis (Fig. 4A) and that the resulting attenuated apoptosis was able to abrogate chemosensitivity (Fig. 3A).
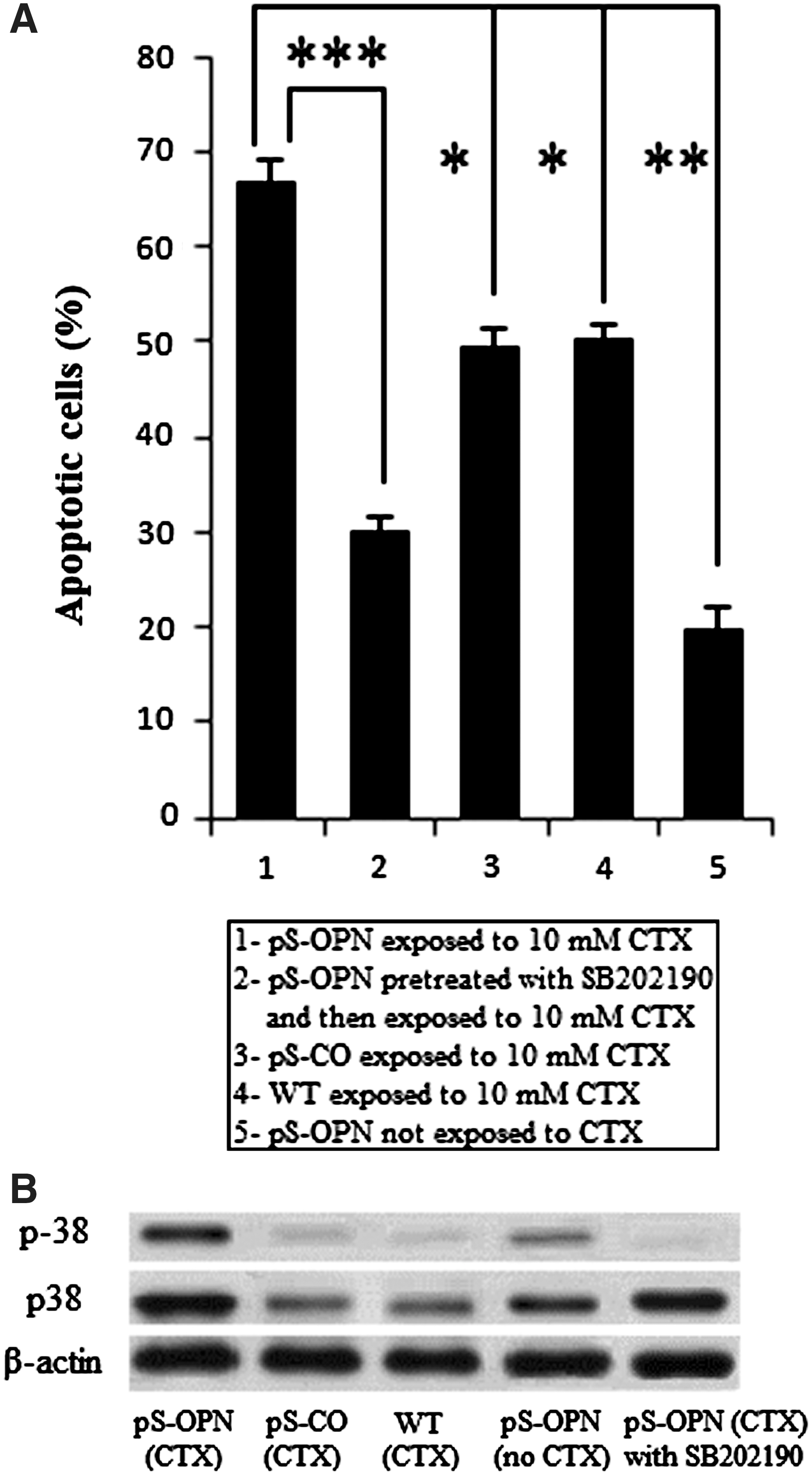
Discussion
CTX, an alkylating agent, exerts its cytotoxic effects via transferring alkyl groups to DNA. The resulting damaged DNA would lead to DNA double-strand breakage (DDSB), which in turn interferes with normal cellular functions and leads to cell death. Apoptosis is the final common pathway of cell death induced by many chemotherapeutic drugs. 14,15 It has been reported that several chemotherapeutic agents induce cancer cell apoptosis through activating p38 MAPK pathway 10,11,16 and that p38 MAPK pathway activation is essential in cell apoptosis in both cancer and noncancer cells. 17,18 Similarly, in the present study, it was found that CTX exerts its toxicity through cell apoptosis by activating p38 MAPK pathway. However, the detailed mechanisms underlying how CTX activates p38 pathway are unknown. Given that DDSB was able to activate p38 MAPK pathway 19 –21 and that CTX could induce DDSB, in the present study, it was speculated that CTX activates p38 MAPK pathway via DDSB.
Results showed that OPN knockdown was able to chemosensitize MDA-MB-231 cells to CTX. This is consistent with studies previously reported in which OPN induces chemoresistance to therapeutic drugs 6,7 and OPN knockdown chemosensitizes hepatocellular carcinoma cells to chemotherapeutic agents through apoptosis. 22 To find out the mechanism wherein OPN knockdown chemosensitized MDA-MB-231 cells to CTX, the effects of OPN knockdown on MDA-MB-231 cells were studied.
It was found that knockdown of OPN was able to inhibit cell proliferation through inducing cell apoptosis by activating p38 MAPK pathway. This is in line with studies reported elsewhere in which suppression of OPN inhibits cell proliferation of other histological types of cancer cells, including liver cancer cells, 22 brain cancer cells, 23 and prostatic cancer cells, 24 but disagrees with a study by Du et al., 25 in which suppression of OPN does not influence cell proliferation of human endometrial cancer cells. This disagreement can be attributed to different histological origins of the experimental cell lines. However, the detailed mechanisms underlying the disagreement are poorly understood.
As mentioned earlier, both CTX and OPN knockdowns were able to induce cell apoptosis through activating p38 MAPK pathway. However, whether the mechanisms underlying chemosensitivity are governed by their synergistic effects on cell apoptosis is unknown. Therefore, the combined effects of CTX and OPN knockdown on MDA-MB-231 cells were investigated. It was found in the present study that OPN knockdown and CTX could induce enhanced apoptosis and enhanced activation of p38 MAPK pathway synergistically. In addition, inhibition of enhanced activation of p38 MAPK pathway can attenuate enhanced cell apoptosis, indicating that OPN knockdown enhances CTX-induced cell apoptosis by enhancing CTX-induced p38 MAPK activation. Moreover, the resulting attenuated enhanced apoptosis in turn abrogated chemosensitivity induced by OPN knockdown. Thus, these findings suggest that OPN knockdown chemosensitizes MDA-MB-231 cells to CTX through enhancing apoptosis by enhancing p38 MAPK activation.
Conclusions
In conclusion, the present work provides evidence for the first time that OPN knockdown is able to chemosensitize MDA-MB-231 cells to CTX. Specifically, the present study shows for the first time that each of OPN knockdown and CTX is able to induce cell apoptosis through activating p38 MAPK pathway and that OPN knockdown and CTX are able to induce enhanced apoptosis through activating p38 MAPK pathway synergistically. Inhibition of p38 MAPK pathway could attenuate enhanced apoptosis and the resulting attenuated enhanced apoptosis in turn abrogated chemosensitivity. Therefore, the present study concludes that knockdown of OPN chemosensitizes MDA-MB-231 cells to CTX by enhancing apoptosis through activating p38 MAPK pathway.
Footnotes
Disclosure Statement
No financial conflicts exist.