
Abstract
Preclinical and early clinical trials indicate that synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN) have potent immunostimulatory effects and can enhance the anticancer activity of a variety of cancer treatments. In this study, CpG ODN1826 was used to increase the radiosensitivity of human pulmonary adenocarcinoma cell line A549, and some underlying mechanisms were also detected. With the treatment of CpG ODN on A549 cells, there was an upregulation of CD40, CD80, CD86, and MHC-II on A549 cells in association with Th1 cytokine production. Further, the protein level of toll-like receptor (TLR)-4 was decreased, whereas TLR-9 was enhanced. Flow cytometry results showed that CpG ODN could increase mitochondrial membrane potential and subsequently promote the apoptosis of A549 cells, resulting in increased radiosensitivity. These results suggest a regulatory role of CpG ODN1826 in enhancing the radiosensitivity of A549 cells by activating the adaptive and innate immune responses, thereby providing a new potential strategy to the treatment of malignant tumors.
Introduction
Lung cancer, for which the incidence, metastasis, and mortality rates are very high, has become more dangerous to aged men and women. Besides surgery, radiotherapy and chemotherapy are effective and widely used. However, the organs and tissues that are adjacent to the cancer-affected lung, such as heart, spinal cord, and normal lung tissue, restrict therapeutical radiation dose. A lower radiodosage to cancer-affected lungs may result in a recurrence, whereas a higher radiodosage harms the peripheral normal tissues around tumors, thereby depressing the quality of life. Thus, it is more stringent to search for a new strategy to enhance the radiosensitivity of tumor cells while lowering the side-effects of radiations. Many investigations suggested that there was a significant relationship between apoptosis rate and radiosensitivity of tumor cells. 1
Synthetic oligodeoxynucleotides (ODN) containing unmethylated CpG motifs (CpG ODN) are immune stimulators. 2 DNA extracted from Mycobacterium bovis-activated natural killer cells to produce IFN and to induce antitumor activities and particular sequences with 50-CpG-30 motif(s) were essential for these effects. 3 Direct conjugation between CpG ODN and Ag improved the therapeutic efficacy of CpG ODN in the treatment of allergies and tumors. 4 Later, synthetic CpG ODN were reported to have similar immunologic effects by activating innate immunity through toll-like receptor (TLR)-9. 5 Among several immunomodulatory effects of CpG ODN, particular interest is in the induction and activation of type 1 T helper (Th1) cells and CD8+ cytotoxic T lymphocytes through the activation and maturation of DC, leading to the secretion of interleukin-2 (IL-12) and the expression of costimulatory molecules. 6
Experiments in murine models suggest CpG ODN can enhance the response to radiation treatment for both immunogenic and nonimmunogenic tumors, 7,8 indicating that the combination of CpG ODN and radiation can be effective. Recent studies demonstrated that the treatment with a combination of CpG ODN and radiation enhances the response and improves the cure rate of murine tumors. 9 The treatment with either CpG ODN or radiotherapy induced complete tumor remissions in one-third of rats inoculated with glioma. 10 When both treatments were combined, complete tumor remission was achieved in two-thirds of the animals. The treatment with CpG ODN and radiotherapy to improve the radiosensitivity by inducing tumor cell apoptosis is intriguing. Thus, the aim of the present study was to investigate the effects of the combined treatment on A549 cells and to elucidate the underlying mechanisms of CpG ODN.
Materials and Methods
Cell culture
Human pulmonary adenocarcinoma cell line A549 cells were kindly provided by Shanghai Cancer Institute and cultured in RPMI 1640 medium (Biowest) with 10% fetal bovine serum and 1% penicillin–streptomycin. Cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2. The pH of the cultures was 7.35 under these conditions.
CpG-ODN1826
The CpG-ODN1826, comprising a nucleotide sequence of 5′-TCCATGACGTTCCTGACGTT-3′, was selected according to a published report 11 and was provided by Coley Pharmaceutical Group, Inc. It was dissolved in water and stored at −20°C. CpG-ODN 1826 (10 μg/mL) was added to a 24-well plate and left for 24 hours. A549 cells without CpG-ODN 1826 acted as control.
Experimental design
The cultured cells were first divided into two groups: control group and CpG ODN-treated group. Based on whether X-irradiation was performed to cultured cells or not, two subgroups emerged: irradiation (IR)-treated group and CpG ODN combined IR-treated group.
Analysis of CD40, CD80, CD86, and MHC-II expression
To analyze CD40, CD80, CD86, and MHC-II on A549 cells, Fc receptors were blocked by normal rat serum for 15 minutes at 4°C. PerCP-conjugated anti-CD11c, FITC-conjugated anti-Gr-1 in combination with PE-conjugated antibodies recognizing CD40, CD80, CD86, and MHC-II, and their respective isotope controls were then added at a concentration of 1 μg per 1 × 106 cells per 100 μL, and the cells were incubated for a further 30 minutes at 4°C. The cells were washed once with ice-cold phosphate-buffered saline (PBS), pH 7.2, containing 0.1% NaN3 and 0.5% BSA and were resuspended in 300 μL PBS. Flow cytometry (FCM) analysis was gated on Gr-1- and CD11c+ cells, and data were analyzed with CellQuest software (both from Becton Dickinson).
Western blot analysis of TLR-4 and TLR-9 expression on A549 cells
A549 cells were washed twice with ice-cold PBS and lysed in HEPES buffer (20 mM, pH 7.4) containing 150 mM NaCl, 100 mM NaF, 10 mM EDTA, 10 mM Na4P2O7, 2 mM sodium orthovanadate, 1 mM PMSF, 2 mg/mL aprotinin, 20 mM leupeptin, and 1% Triton X-100 for 10 minutes on ice. After clarification of the cell lysates by centrifugation at 13,000 g for 15 minutes, the supernatants were collected and their protein concentrations were determined using a bicinchoninic acid protein assay kit (Hyclone Pierce).
The lysates were loaded on 12% SDS-PAGE gels, run for 1.5 hours at constant voltage (150 V), and electrotransferred (2 hours, 720 mA, 4°C) onto PVDF membranes. The PVDF membrane was blocked in 5% nonfat dried milk in PBS-Tween for 1 hour at room temperature, and then it was probed with primary antibodies to the following proteins for 1 hour: TLR-4 (1:1000) and TLR-9 (1:1000). The membrane was then incubated with horseradish peroxidase-conjugated goat antibodies for 1 hour, and immunoreactive bands were detected by a supersignal chemiluminescent system (Pierce Chemical).
Cytokine measurements
The concentrations of TNF-α and IL-12 in the supernatants of A549 cells with or without CpG ODN were measured by ELISA Development kits (Genzyme-Techne).
Radiation exposure
A549 cells were exposed to 8 Gy of 6 megavoltage (MV) X-rays with 5 × 10 cm2 field size at room temperature using a linear accelerator (ELEKTA). The dose rate was 2.0 Gy/min, which was determined by Fricke's chemical dosimeter.
Apoptotis assay
The experiments were performed using the Annexin V Apoptosis Detection Kit (BD Biosciences Pharmingen). According to the manual, cell pellets were resuspended in 100 μL binding buffer and stained with 5 μL Annexin V and 5 μL propidium iodide staining solution in the dark at room temperature for 15 minutes. FCM acquisition was performed on FACScan (Becton Dickinson) using the CELL Quest program.
Mitochondrial membrane potential
Mitochondrial membrane potential (Δψm) was assessed using JC-1, a cationic dye that exhibits potential-dependent accumulation and formation of red fluorescent J-aggregates in mitochondria. In contrast, the changes in plasma membrane potential do not affect the JC-1 status. The JC-1 monomer accumulates in the cytoplasm, where it produces green fluorescence. The formation of J-aggregates in the mitochondria is indicated by a fluorescence emission shift from green (525 nm) to red (590 nm). 12 Mitochondrial depolarization is indicated by a decrease in the red/green fluorescence intensity ratio. At different time points of cisplatin exposure, A549 monolayers were loaded with 10 μM JC-1 for 30 minutes at 37°C. After loading, media were aspirated, and the monolayers were kept on ice, washed twice with ice-cold PBS, scraped off culture dishes, washed, and resuspended in PBS. Fluorescence was analyzed by FCM (FACSCalibur; BD Biosciences) using excitation by a 488-nm argon-ion laser. The JC-1 monomer (green) and the J-aggregate (red) were detected separately in FL1 (emission, 525 nm) and FL2 (emission, 590 nm) channels, respectively.
DNA ladder
Cells were incubated for 12 hours at 37°C in lysis buffer (10 mM Tris-HCl [pH 8.0]/10 mM NaCl/10 mM EDTA/0.5% SDS/100 μg/mL proteinase K). DNA was isolated by chloroform extraction and treated with 50 μg/mL RNaseA (Sigma) for 1 hour at room temperature. DNA samples (20 μg) were separated by electrophoresis in a 1.5% agarose gel. The gel was stained with SYBR Green I nucleic acid gel stain (Molecular Probes) after electrophoresis, and the DNA was visualized under UV light.
Statistical analysis
All results are presented as mean value ± SD. Student's t-test was used to test the differences between groups. Statistical significance was considered at p < 0.05.
Results
CpG ODN upregulated the expression of CD40, CD80, CD86, and MHC-II
The expressions of CD40, CD80, CD86, and MHC-II on control A549 cells were all kept at low levels, whereas they were significantly upregulated by the treatment with 10 μg/mL CpG ODN for 24 hours (Fig. 1 and Table 1) (p < 0.05, <0.001, <0.05, <0.001, respectively).
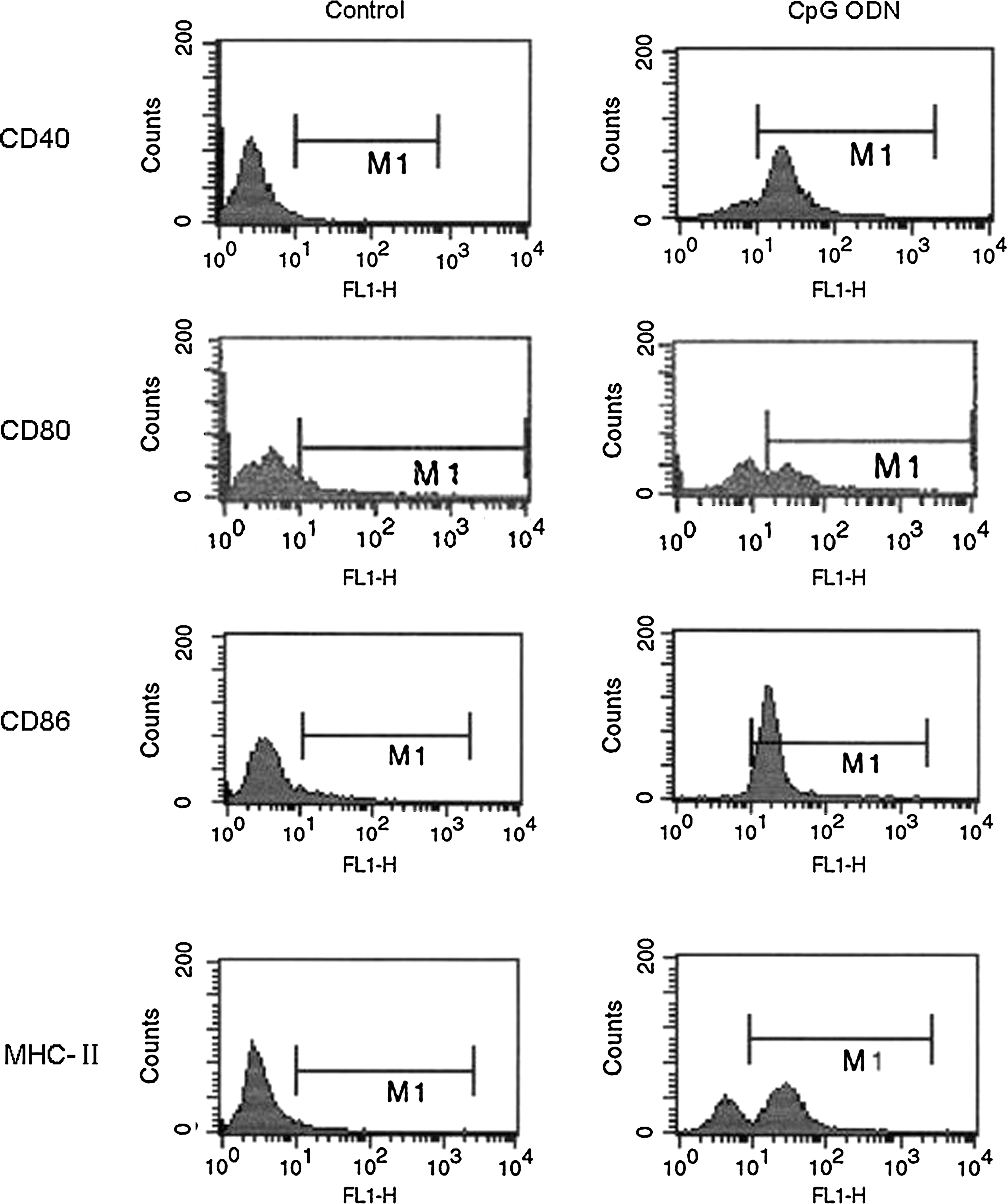
Flow cytometry (FCM) analysis of CD40, CD80, CD86, and MHC-II on A549 cells. TNF-α, tumor necrosis factor–α; CpG ODN, synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides.
Differences are shown between control and synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN)–treated cells. Values are means for five repeated experiments.
p < 0.01 versus the control.
p < 0.01 versus the control.
CpG ODN increased the release of Th1 cytokines
The production of Th1 cytokines, such as TNF-α and IL-12, was also analyzed. With the treatment of CpG ODN for 24 hours, A549 cells produced significantly more TNF-α than the control cells (Fig. 2). The production of IL-12 was also comparable between the CpG ODN-treated cells and the control cells. When CpG ODN was administered to the cells, the amount of TNF-α produced by A549 was approximately tripled, whereas the amount of IL-12 was approximately doubled (Fig. 3).
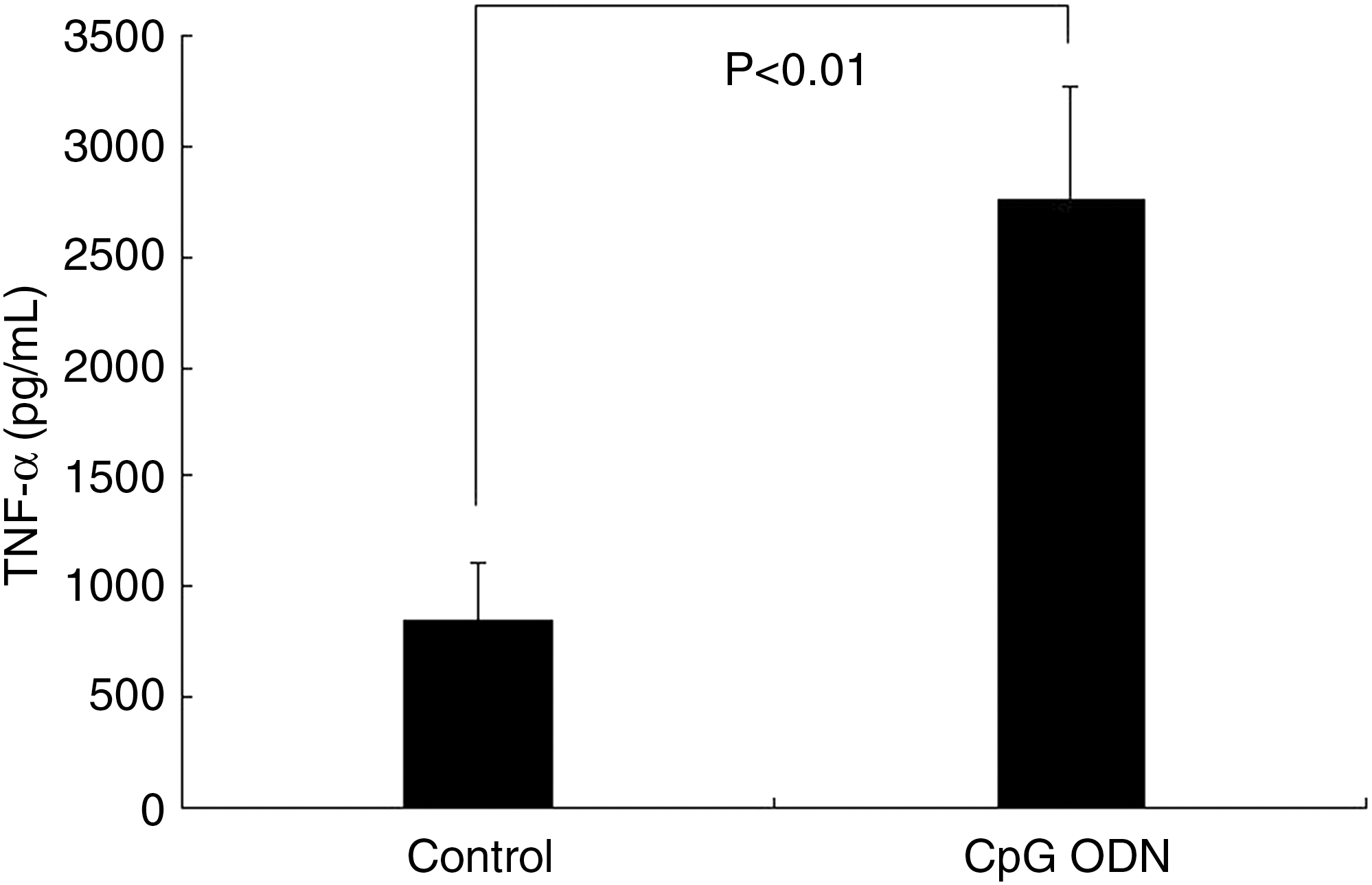
Tumor necrosis factor–α (TNF-α) production of A549 cells. Values are means for five repeated experiments.

Interleukin-12 (IL-12) production of A549 cells. Values are means for five repeated experiments.
Effects of CpG ODN on TLR-4 and TLR-9 protein levels
The protein level of TLR-9 was kept at a low level on control A549 cells. With the treatment of 10 μg/mL CpG ODN for 24 hours, TLR-9 protein level was much higher than the control (p < 0.01), whereas the TLR-4 protein level was much lower than the control cells (p < 0.05) (Figs. 4 and 5).

Western blot analysis of toll-like receptor (TLR)–4 and TLR-9 proteins on A549 cells. CpG ODN, synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides.
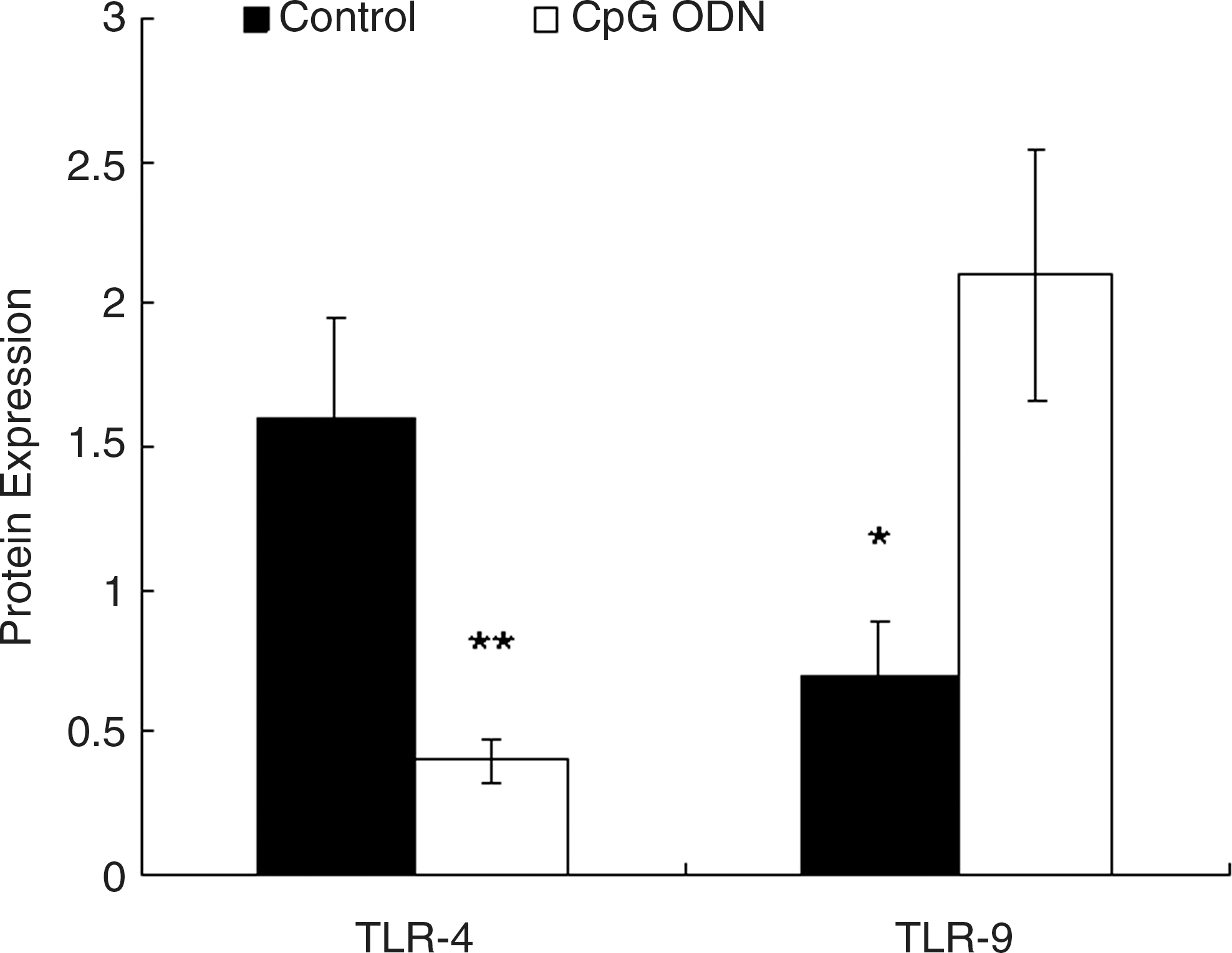
Quantification of blots in Figure 4, normalized to β-actin. Differences are shown between the control cells and the synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN)-treated cells. Values are means for five repeated experiments. *p < 0.05 versus the CpG ODN-treated cells. **p < 0.01 versus the control. TLR, toll-like receptor.
CpG ODN accelerated A549 cell apoptosis
DNA ladder showed isometric strips, which were ruptured DNA fragments on the gel (Fig. 6). FCM analyses suggested that the apoptosis index of the CpG ODN + IR-treated group, the IR group, and the CpG ODN-treated group are 16.85% ± 1.39%, 11.21% ± 0.74%, and 5.24% ± 0.65%, respectively, which were all higher than that of the control group (p < 0.001, p < 0.001, and p < 0.05, respectively) (Fig. 7 and Table 2).

DNA ladder. Lane 1, synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN) + IR-treated cells; lane 2, irradiation (IR)–-treated cells; lane 3, control cells; lane 4, marker; lane 5, CpG ODN-treated cells.

FCM analysis of apoptosis index of A549 cells.
Differences are shown between the control and synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN)–treated cells, the control and irradiated (IR)–treated cells, and the control and CpG ODN + IR-treated cells. Values are means for five repeated experiments.
p < 0.05 versus the control.
p < 0.01 versus the control.
AI, apoptosis index.
CpG ODN damages the mitochondrial membrane function
Mean absolute fluorescence values and ratio were measured in A549 cells (Fig. 8 and Table 3). The orange/green (shown in black and white in Fig. 8) fluorescence ratios of the CpG ODN-treated cell, the IR-treated cells, and the CpG ODN + IR-treated cells were much lower than that of the control cells (p < 0.05, <0.001, and <0.001, respectively).
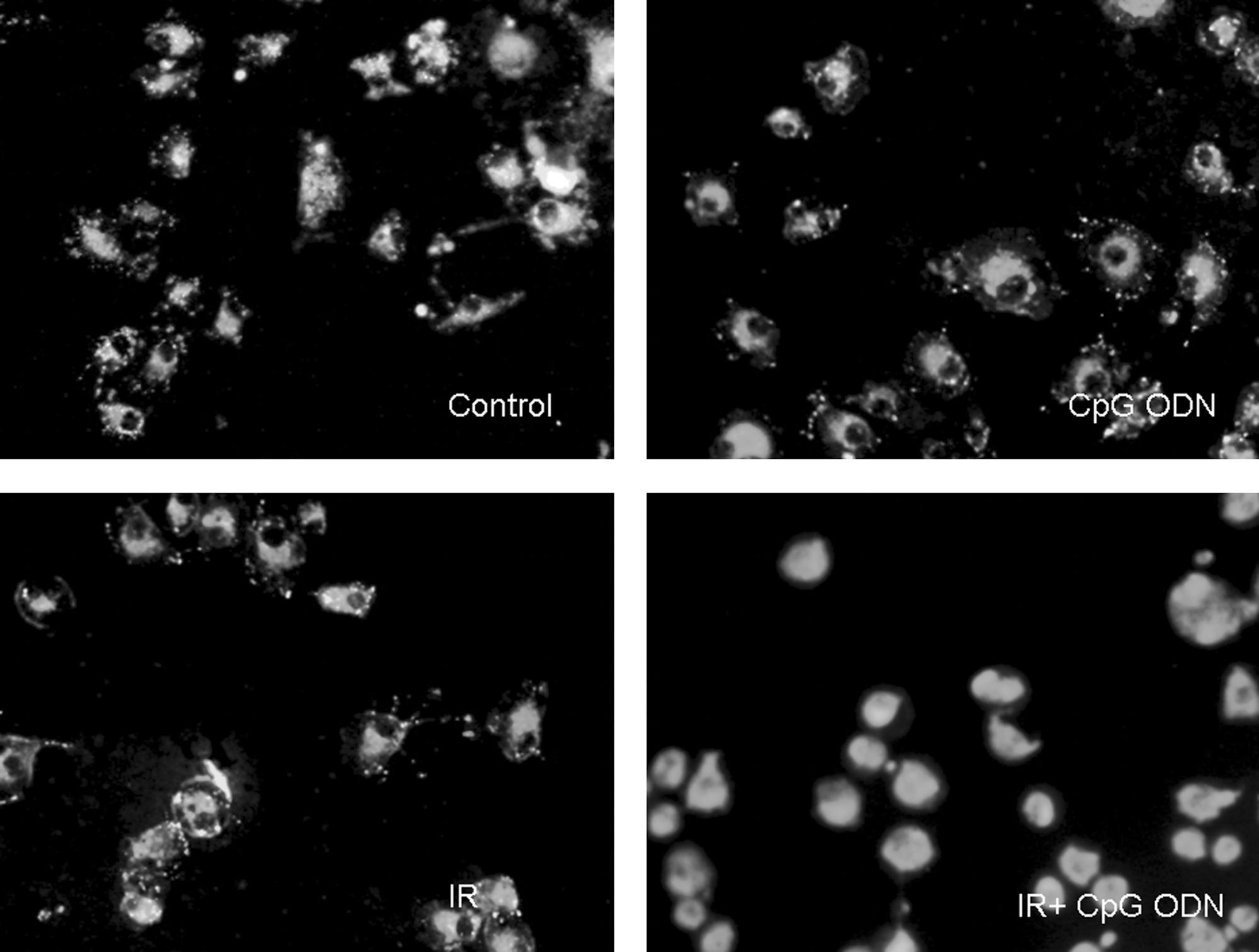
Fluorescent image showing A549 cells loaded with the mitochondrial membrane potential indicator JC-1.
Differences are shown between the control and synthetic oligodeoxynucleotides containing unmethylated CpG dinucleotides (CpG ODN)–treated cells, the control and irradiated (IR)–treated cells, and the control and CpG ODN + IR-treated cells. Values are means for five repeated experiments.
p < 0.05 versus the control.
p < 0.01 versus the control.
Discussion
Although surgery, chemotherapy, and radiotherapy are effective cancer treatments, some cancers are refractory to these treatments. Effective treatment of advanced and metastatic cancers and the prevention of recurrence are especially difficult. 13 Immunotherapy combined with radiotherapy are promising approaches to cancer treatment and prevention, which may someday overcome the shortcomings of traditional cancer managements. In the present study, the promising effects of CpG ODN1826 in increasing the radiosensitivity of A549 cells were demonstrated, and some immunologic mechanisms of CpG ODN were also explored.
Two different signals are required to prime and activate naive CD4+ and CD8+ T cells. 14 First, antigenic peptides must be presented on the surface of activated antigen-presenting cells (APCs) by MHC class I or II molecules to CD8+ or CD4+ T cells, respectively. The binding of peptide/MHC complexes to TCRs mediates a signal into the T cells. A second signal must be mediated from costimulatory molecules on activated APCs to T cells. The absence of MHC or costimulatory signals on tumors induces the anergy of host immunosurveillance. 15 Advances in the understanding of the first signal and costimulatory signals have provided a vast array of novel approaches to prevent cancer. The effects of CpG ODN as an immunoactivator were elucidated by downregulating the expressions of MHC-II, CD80, CD86, and CD40.
Because of the pivotal role of TLR in immune responses to pathogens, most research aimed at understanding the TLR biology has been focused on immune cells. However, TLRs are also expressed on tumor cells. 16 In the patients with cancer, who are often immunosuppressed, TLR-mediated activation of innate or adaptive immunity could be of considerable benefit. 17 It has been known for a long time that microbial compounds can be used as efficient adjuvants in antitumor vaccine formulations, and numerous animal tumor models clearly indicate the potency of different TLR agonists in enhancing antitumor immune responses. 18 TLR-9 has been described as the only receptor of CpG ODN. In the present study, it was found that TLR-9 protein was expressed on A549 cells, and the protein level could even be significantly enhanced with the treatment of CpG ODN. Further, it was found that CpG ODN could decrease the protein level of TLR-4. The precise mechanisms of CpG ODN involved in the regulation of TLR-4 and TLR-9 remain unclear. However, other studies demonstrated that TLR-9 stimulation on lung cancer cells sensitized tumor cells to apoptosis, leading to the arrest of tumor growth. 19,20 Ligation of TLR-4 expressed on tumor cells can induce chronic inflammation, which promotes tumor growth. 21,22 Signaling via the TLR-4 expressed on human tumor cells induces tumor growth and facilitates tumor escape from immune surveillance. 23
In the present study, it was found that TNF-α and IL-12 were elevated in cell culture supernatants after CpG ODN treatment, and the increased secretion of TNF-α and IL-12 contributed to enhanced radiosensitivity in a subsequent research. TNF-α and IL-12 were reported to be upregulated in the human glioma cell line after combined treatment with CpG ODN and β-rays and were apparently associated with the inhibition of clonogenic cell survival (24). The present study's results therefore suggest that enhanced secretion of TNF-α contributes to the radiosensitization induced by CpG-ODN.
X-irradiation induces apoptosis through DNA damage and generation of free radicals in many types of cells. So radiation therapy is used extensively in the management of patients with malignant tumor, including lung cancer. Despite the widespread use of radiation therapy for the treatment of lung cancer, the uniform side-effects of radiations remain a critical problem in the management of these patients and so enhancing the radiosensitivity of this malignant tumor while lowering the side-effects of radiations is more stringent. The above results manifest the apoptosis-inducing effects of CpG ODN on A549 cells, and so CpG ODN was further utilized as a radiosensitizer to detect the synergy between CpG ODN and X-rays on A549 cells. The satisfying FCM results indicated that CpG ODN in combination with X-rays could more significantly accelerate the apoptosis of A549 cells in comparison with CpG ODN or X-rays alone.
In the present study, Δψm in A549 cells was tested using the fluorescent dye, JC-1, which aggregates into healthy mitochondria and fluoresces red. Upon mitochondrial collapse in apoptotic cells, JC-1 dye no longer accumulates and is instead distributed throughout the cell, resulting in a decrease of red fluorescence. In accordance with this, it was found that CpG ODN in combination with X-rays indeed disrupted the mitochondrial membrane potential much more than the control cells, X-ray–treated cells, and CpG ODN-treated cells, as observed by a decrease in the intensity of red fluorescence.
Conclusions
In summary, the present study shows that CpG ODN sensitizes A549 cells to radiation in vitro. The radiosensitization induced by CpG ODN provides a rationale for future investigation of the therapeutic efficacy of CpG ODN in combination with radiotherapy.
Footnotes
Acknowledgments
This study was funded by the Science Foundation of Shanghai Health Bureau (2006-8).
Disclosure Statement
No financial conflicts of interest exist.