
Abstract
Curcuma aromatica is a common Chinese herb for treating diseases with blood stasis and has been regarded as an anticancer herb in modern clinical practice. However, the anticancer effects and related molecular mechanisms of Curcuma aromatica remain unclear. In the present study, human colon carcinoma LS-174-T cell line with wild-type p53 was used as a model cell to evaluate the anticancer effects of aqueous extract of Curcuma aromatica (AECA). AECA inhibits LS-174-T cell proliferation in a dose- and time-dependent manner and colony formation in a dose-dependent manner. AECA treatment induces apoptosis accompanied by caspase-8, -9, and -3 activation in LS-174-T cells. Moreover, blocking the activities of these caspases with a specific inhibitor significantly protected LS-174-T cells from AECA-induced apoptosis. AECA treatment also induces G2/M phase arrest in LS-174-T cells. Expression of p53 was unchanged after AECA treatment; specific silence of p53 did not influence AECA-induced apoptosis and G2/M phase arrest. Further, the expression of cyclin B1 and CDK1 was reduced by AECA. This study suggests that AECA might be effective as an antiproliferative herb for colon carcinoma, the antitumor activity of AECA may involve both extrinsic and intrinsic apoptosis, and AECA induces G2/M phase arrest via downregulation of cyclin B1 and CDK1 and without the participation of p53.
Introduction
The current therapy of colon cancer mainly depends on surgery, chemotherapy, and target therapy, such as Erbitux (cetuximab) and Vectibix (panitumumab), which have been approved by the Food and Drug Administration for metastatic colorectal cancer. Therapeutic progress and dietary and screening programs have facilitated an overall decline in the mortality of colon cancer. Nevertheless, colon cancer remains the third leading cause of cancer death in the United States, and the incidence of colon cancer is increasing in Asian countries. 1,2 Clearly, the development of a novel therapeutic model for colon cancer is highly warranted.
In China, Traditional Chinese Medicine (TCM) has long been used for treating various diseases including cancer. Cancer clinical manifestation and causes have been well described in Chinese medical literature since 400 B.C. 3 The pathogenesis of cancer is related to weakened body resistance, blood stasis, stagnation of phlegm, and poison according to TCM theory, and cancers are recognized as a syndrome of blood stasis, phlegm, and toxicity in TCM. 3 –7 Dissipating blood stasis is one of the most important therapeutic principles for cancer treatment in TCM. Yujing (Curcuma aromatica, Curcuma) is a commonly used Chinese herb for treating diseases with blood stasis and has been regarded as an anticancer herb in modern TCM practice. However, the anticancer effects and related molecular mechanism of Curcuma aromatica remain unclear.
In this study, human colon carcinoma LS-174-T cell with wild-type p53 was used as a model cell to evaluate the anticancer effects of Curcuma aromatica. The results of the present study show that the aqueous extract of Curcuma aromatica (AECA) inhibits cell proliferation and induces caspase-mediated apoptosis as well as G2/M cell cycle arrest in LS-174-T cells in a p53-independent manner.
Methods
Materials
Cell Counting Kit-8 (CCK-8) was purchased from Dojindo. Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum were obtained from Hyclone. Propidium iodide, Z-DEVD-FMK, Z-IETD-FMK, and Z-LEHD-FMK were from Sigma-Aldrich. Colorimetric CaspACE™ Assay System was the product of Promega, Caspase-8, and Caspase-9 Colorimetric Assay kits were from R&D Systems. Antibodies against p53, cyclin B1, CDK1 (CDC2), and β-actin were procured from Cell Signaling Technology or BD Biosciences Pharmingen. Small interfering RNA (siRNA) against p53 and the control siRNA were procured from Santa Cruz Biotechnology (Santa Cruz, CA). Lipofectamine™ 2000 was obtained from Invitrogen.
Extraction of Curcuma aromatica
Authentic Curcuma aromatica herb material was obtained from Longhua hospital herb store and was decocted two times with 8 volumes of boiling distilled water for 2 hours. The decoction was centrifuged twice (12,000 g, 30 minutes) to remove insoluble ingredients, precipitated with 50% alcohol, and lyophilized. The dried extract was weighed, dissolved in DMEM, and sequentially passed through 0.45- and 0.22-μm filters for sterilization.
Cell culture
Human colon carcinoma cell LS-174-T with wild-type p53 was obtained from the cell bank of Chinese Academy of Sciences. Human colon epithelial cell CRL-1790 was obtained from American Type Culture Collection. LS-174-T and CRL-1790 cells were grown in DMEM with 10% fetal bovine serum and 1% Pen–Strep and maintained in a 37°C incubator with a humidified 5% CO2 atmosphere.
Growth inhibition assay
The cells in logarithmic growth phase were seeded into a 96-well plate (5 × 103 cells/well) and allowed to attach for 24 hours before treatment. The cells were exposed to various doses of AECA for 72 hours, and cell viability was evaluated every 24 hours by using the CCK-8 colorimetric assay according to the manufacturer's instructions.
For colony formation assays, the cells were plated in a six-well plate (2 × 103 cells/well) and allowed to adhere for 24 hours before treatment. The cells were exposed to different concentrations of AECA and allowed to grow for 2 weeks (until colonies were visible to the naked eye). The colonies were stained with Giemsa and counted.
Flow cytometric analysis
At the end of treatment, both floating and attached cells were collected and washed with phosphate-buffered saline, fixed in 70% ethanol at 4°C, and treated with 10 mg/mL RNase for 30 minutes at 37°C. Finally, the cells were stained with propidium iodide (50 μg/mL) and analyzed in a FACSCalibur flow cytometer (Becton Dickinson).
Colorimetric caspase activation assay
After treatment for the indicated time with various concentrations of AECA, caspase-3, -8, and -9 activities were measured by the cleavage of the specific chromogenic substrate. Caspase-3 activity was measured by Colorimetric CaspACE Assay System, and caspase-8 and -9 activities were determined by Caspase-8 and Caspase-9 Colorimetric Assay kits according to manufacturer's instructions. For caspases inhibition, the cells were preincubated for 2 hours with 50 μM caspase-3 inhibitor Z-DEVD-FMK, caspase-8 inhibitor Z-IETD-FMK, or caspase-9 inhibitor Z-LEHD-FMK for 2 hours, respectively.
Western blot analysis
Western blots were performed as previously described. 8,9 Briefly, the collected cell sample was lysed, subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel, and transferred onto a polyvinylidene difluoride membrane (Amersham). The transferred or stripped membrane was blocked with 5% nonfat milk, washed, and probed with the indicated antibodies. Blots were then washed, incubated with IRDye 700- and IRDye 800-conjugated secondary antibodies (Rockland Immunochemicals), and visualized by Odyssey Infrared Imaging System (LI-COR Biosciences). For stripping, the blots were incubated in Restore Western Blot Stripping Buffer (Pierce) for 15 minutes at room temperature and washed with phosphate-buffered saline.
siRNA transfection
For siRNA transfection, LS-174-T cells were cultured on a six-well plate to 60% confluence, and 80 pmol specific or nonspecific control siRNA was introduced into the cells using Lipofectamine 2000 according to the manufacturer's recommendations. After 24 hours of transfection, the cells were treated with 100 μg/mL of AECA or same volume of DMEM for 48 hours and harvested for Western blot, flow cytometric analysis, or caspase activity assay.
Statistical analyses
Results were expressed as means ± standard deviation of at least two independent experiments, each conducted in triplicate. Differences between control and Curcuma aromatica treatment were analyzed by one-way analysis of variance. Differences were considered significant at p < 0.05.
Results
AECA inhibits proliferation of LS-174-T cells
First, the effects of AECA on proliferation of LS-174-T cells were examined. LS-174-T cells were cultured in the presence of various concentrations of AECA (0–500 μg/mL) for 24, 48, or 72 hours, and cell viability at each time point was measured by CCK-8 assay. As shown in Figure 1A, proliferation of LS-174-T cells were significantly inhibited in a time- and dose-dependent manner, with maximal inhibition following 72 hours of exposure to 500 μg/mL AECA (p < 0.05). On the other hand, survival of CRL-1790 cells, the normal epithelial cells, was minimally affected by AECA treatment even at concentrations that were highly toxic to LS-174-T cells (Fig. 1B). These data suggest that AECA may inhibit cell proliferation in LS-174-T cells.
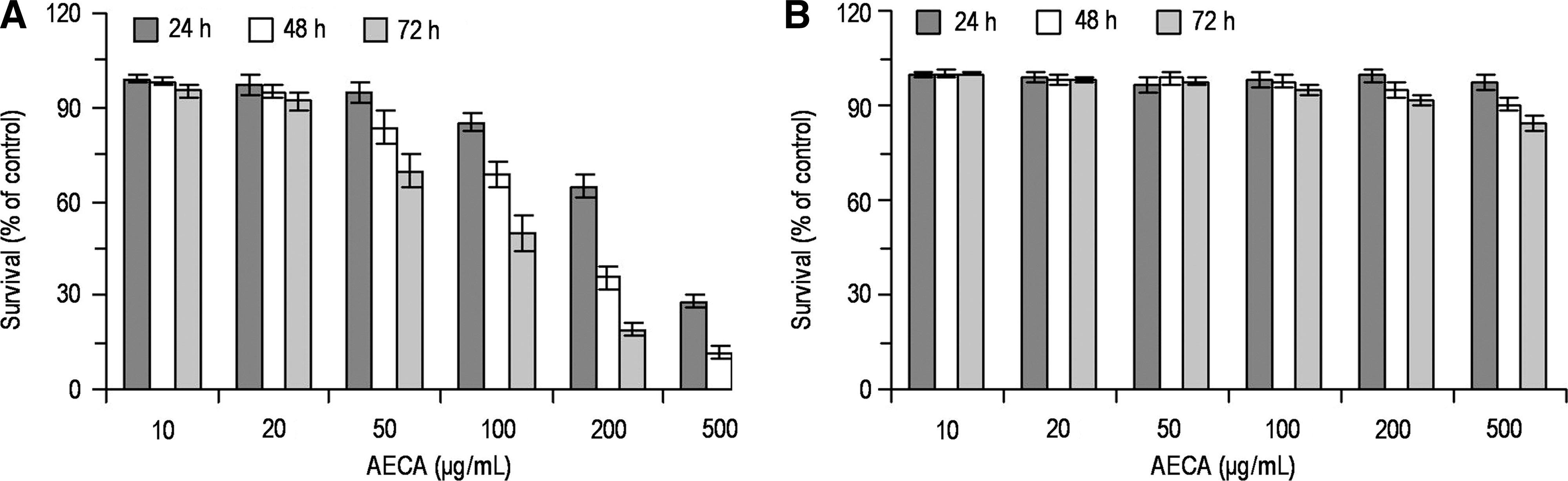
Effects of aqueous extract of Curcuma aromatica (AECA) on the proliferation of LS-174-T and CRL-1790 cells. Human colon carcinoma LS-174-T cells
AECA inhibits colony formation of LS-174-T cells
The long-term effect of AECA was investigated in colony formation assays. Figure 2A and B shows that colony formation of LS-174-T cells was significantly inhibited by AECA treatment even at concentrations as low as 10 μg/mL (p < 0.01). These observations suggest that AECA may inhibit colony formation in LS-174-T cells.
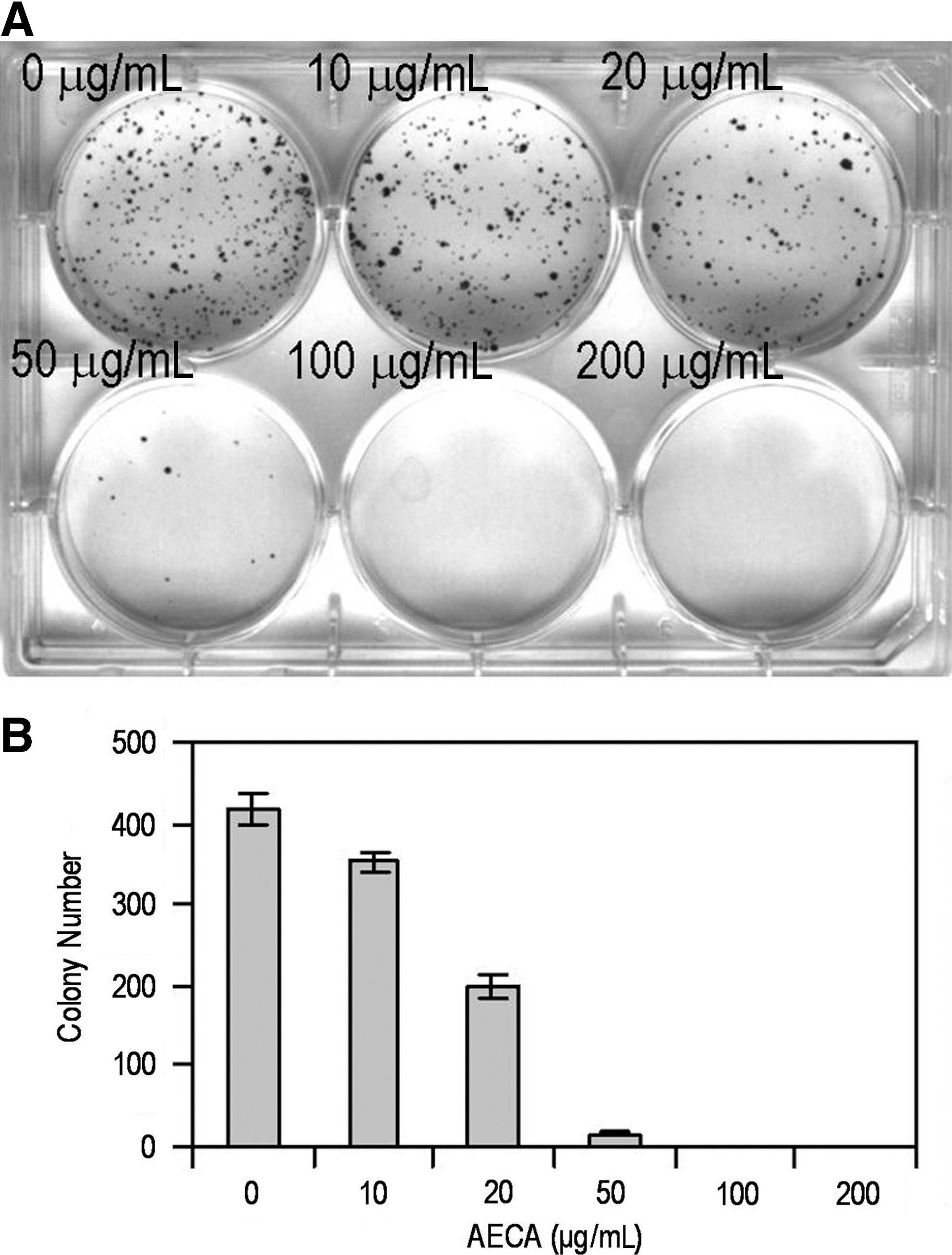
Effects of aqueous extract of Curcuma aromatica (AECA) on LS-174-T cells colony formation. LS-174-T cells were plated on a six-well plate, exposed to various doses of AECA, and allowed for formation of colonies. The colonies were stained, and counted.
AECA induces apoptosis and cell cycle arrest in G2/M
Induction of apoptosis had been recognized as a major anticancer therapeutic response. 10 –12 So, next, the effects of AECA on cell apoptosis and cell cycle distribution in LS-174-T cells were determined by flow cytometry analysis. As shown in Figure 3, apoptosis was detected in LS-174-T cell following 72 hours of exposure to various doses of AECA in a dose-dependent manner and compared with the controls (p < 0.05). In addition, the treatment of LS-174-T cells with AECA resulted in increased number of cells in the G2/M phase, with concomitant reduction of cell number in the S phase (p < 0.05). These results suggest that AECA may induce apoptosis and G2/M arrest in LS-174-T cells.

AECA treatment induces apoptosis and cell cycle arrest in LS-174-T cells. LS-174-T cells were treated with indicated doses of aqueous extract of Curcuma aromatica (AECA) for 72 hours, and DNA content was analyzed by flow cytometry analysis.

AECA activates caspases in LS-174-T cells
Cell apoptosis is executed by a caspase cascade, so whether AECA could induce apoptosis by caspase signaling was also addressed. LS-174-T cells were treated with various doses of AECA for 72 hours and cell lysates were examined for caspase-8, -9, and -3 activities. As shown in Figure 4, AECA activated caspase-8, -9, and -3 in a dose- and time-dependent manner, compared with controls (p < 0.05). To identify whether the activation of caspases is an essential event for the apoptosis induced by AECA, caspase-3, -8, and -9 activities were inhibited by specific inhibitors (Fig. 5A). It was observed that a caspase-3 inhibitor (Z-DEVD-FMK), a caspase-8 inhibitor (Z-IETD-FMK), and a caspase-9 inhibitor (Z-LEHD-FMK) significantly protected LS-174-T cells from AECA-induced growth inhibition and apoptosis (Fig. 5B, C). These data demonstrate that the activation of caspase-3, -8, and -9 is an essential event for AECA-induced apoptosis.

Aqueous extract of Curcuma aromatica (AECA) activates caspases in LS-174-T cells. LS-174-T cells were treated with different doses of AECA; caspase-8

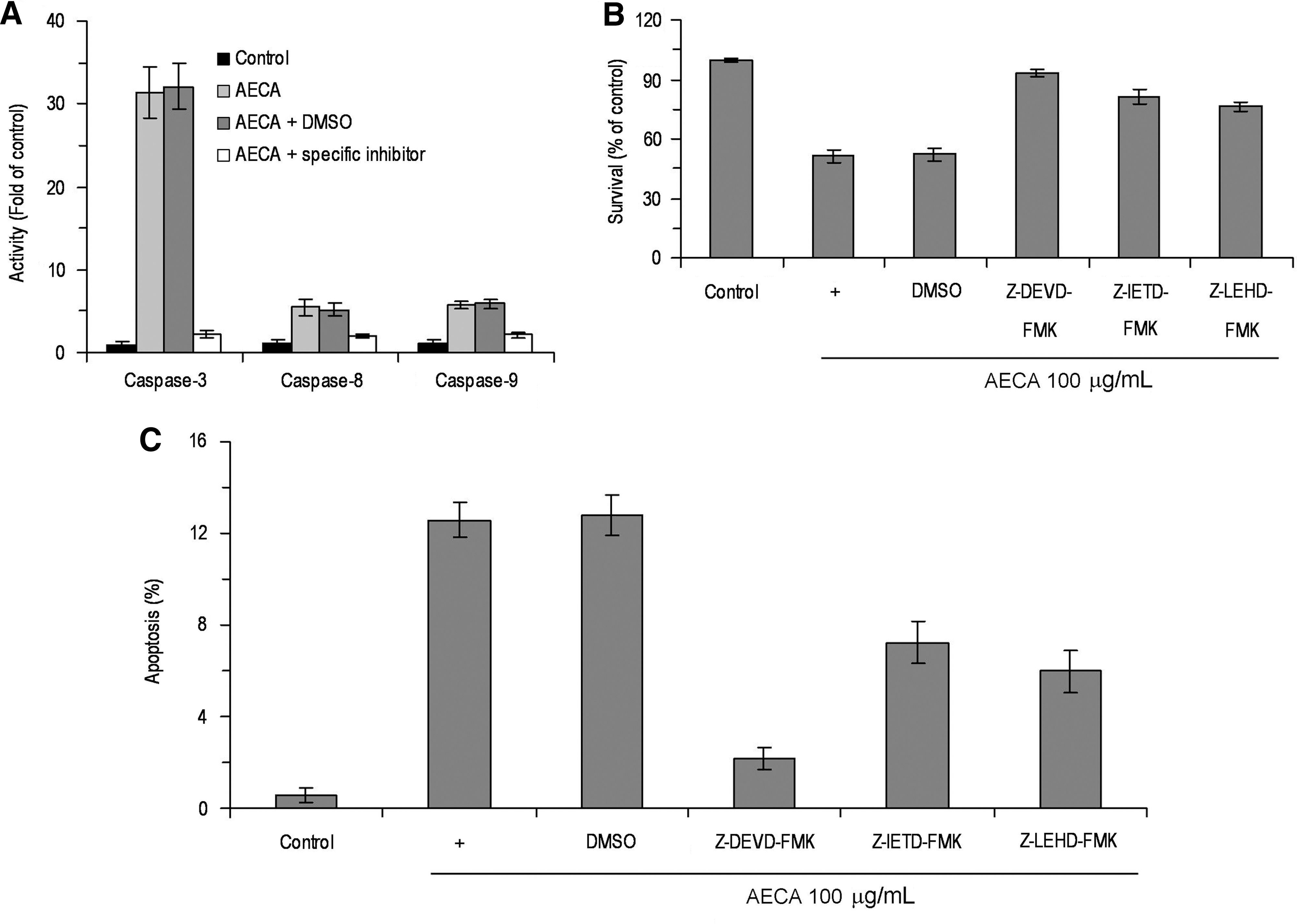
Effect of caspase inhibitors on aqueous extract of Curcuma aromatica (AECA)–induced apoptosis in LS-174-T cells. After pretreatment with 50 μM caspase-3 inhibitor Z-DEVD-FMK, caspase-8 inhibitor Z-IETD-FMK, or caspase-9 inhibitor Z-LEHD-FMK for 2 hours, respectively. LS-174-T cells were exposed to 100 μg/mL AECA for 72 hours and subjected to caspase activation assay
Role of p53 in AECA-induced apoptosis and cell cycle arrest
The tumor suppressor p53 is an evolution conserved “cellular gatekeeper,” and in response to various stress, p53 may efficiently inhibit cell proliferation by both blocking cell cycle progression and promoting apoptosis and play an important role in cancer therapeutic response.
13
In the present study, whether AECA-induced apoptosis in LS-174-T cells was associated with p53 was examined. Surprisingly, the expression of p53 was not changed after AECA treatment (Fig. 6A). So, the expression of p53 by RNA interference was silenced. The transfection efficiency of Lipofectamine 2000 was confirmed to be nearly 90% (Supplementary Fig. S1; Supplementary Data are available online at

The role of p53 in aqueous extract of Curcuma aromatica (AECA)–induced apoptosis and cell cycle arrest. Effects of AECA on p53 expression were identified by Western blot
AECA modulated the expression of G2/M cell cycle regulatory genes
As shown in Figures 3 and 6C, AECA treatment resulted in a significant arrest of cells in the G2/M phase. To further address the molecular basis of AECA-mediated G2/M arrest, its effect on G2/M cell cycle regulatory genes was determined. Exposure of LS-174-T cells to different doses of AECA for 72 hours significantly reduced the expression of cyclin B1 and CDK1 when compared with control cells (Fig. 7). These results suggest the possible involvement of downregulation of cyclin B1 and CDK1 in AECA-induced G2/M cell cycle arrest.
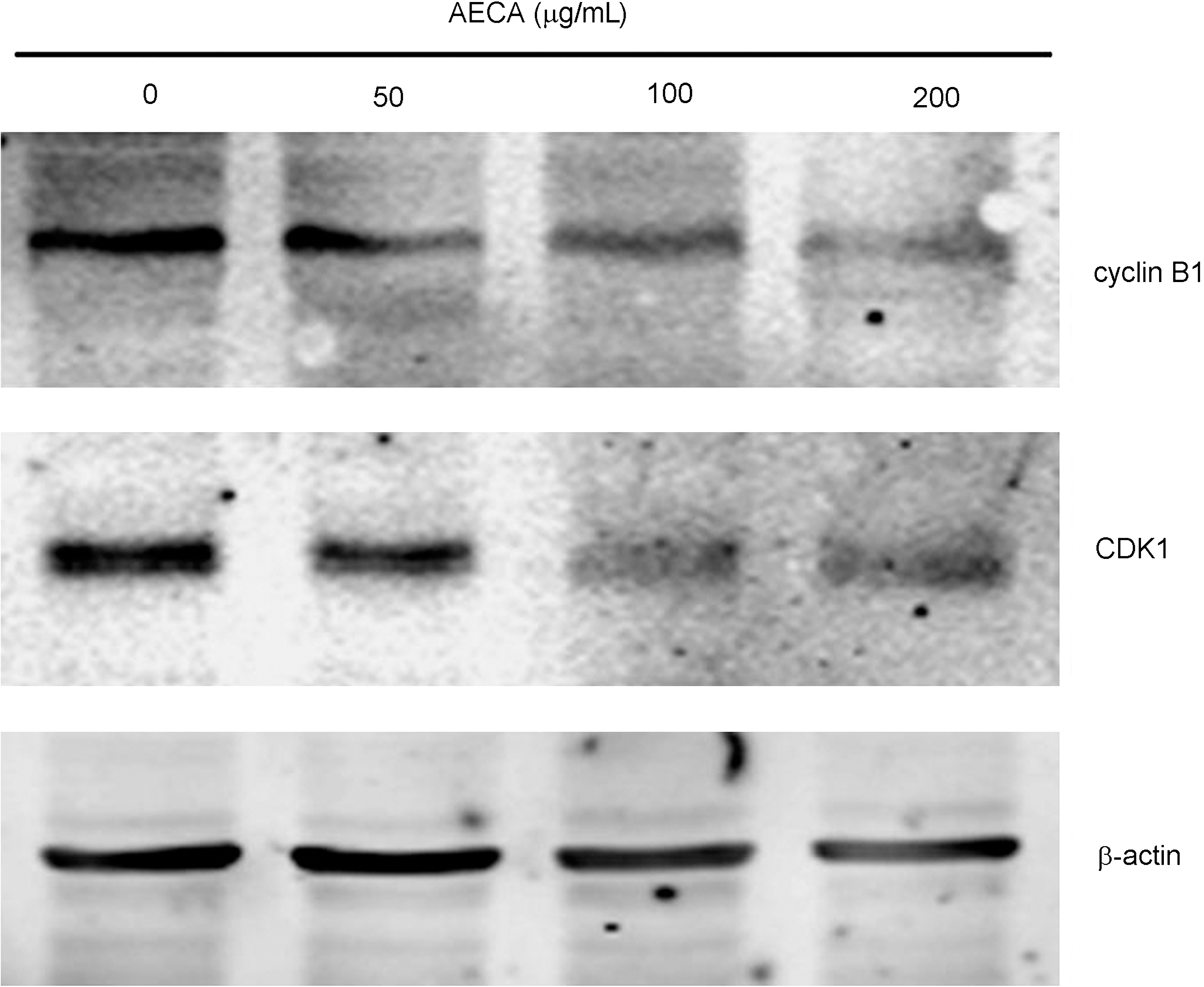
Effects of aqueous extract of Curcuma aromatica (AECA) on expression of G2/M cell cycle regulatory genes. LS-174-T cells were collected after AECA treatment and subjected to Western blots using antibody against cyclin B1, CDK1, or β-actin.
Discussion
Several observations have been made in the present study concerning the anticancer effects of AECA. The data in the present study demonstrated that AECA could inhibit proliferation and colony formation in LS-174-T cells. On the other hand, normal CRL-1790 cells were minimally affected with AECA treatment even at concentrations that were highly toxic to LS-174-T cancer cells. Curcuma aromatica, as a natural Chinese herb, has been used for treating diseases with blood stasis for past 1000 years, and thus, the safety of Curcuma aromatica was well established; this may partially explain why AECA spares normal cell from cytotoxicity induction, making it a relatively safe anticancer herb. However, what makes the different effects of AECA against cancer and normal cells needs more investigation. Further studies indicated that the proliferation inhibitory activity of AECA may be related to the induction of apoptosis and cell cycle arrest.
Apoptosis is an evolutionarily conserved cell suicide process that acts to balance mitosis in the development and maintenance of tissue homeostasis for the removal of superfluous, transformed, or damaged cells and has been recognized as a major anticancer treatment response. 11,12,14 Two major pathways have been identified in the process of apoptosis: the death receptor-induced extrinsic pathway and the mitochondria-apoptosome-mediated apoptotic intrinsic pathway. The intrinsic pathway involves the signals to mitochondria, which lead to the release of cytochrome c and Apaf-1, forming an apoptosome that activates the initiating protease caspase-9, which in turn activates the executioner caspase-3, causing the cell to undergo apoptosis. The extrinsic pathway involves oligomerization of cell-surface death receptors by their ligands, resulting in recruitment and activation of caspase-8 followed by activation of executioner caspases-3. 12,15,16 Caspases-3, -8, and -9 are activated in response to AECA. Blocking the activities of these caspases with a specific inhibitor significantly protected LS-174-T cells from AECA-induced apoptosis, suggesting that both extrinsic and intrinsic apoptotic signals are activated by AECA.
The tumor suppressor p53 is activated in response to a range of stress signals, including DNA damage, oncogene activation, or hypoxia signals. 12,17 Based on the nature of DNA damage, p53 may selectively discriminate between promoters in the induction of target genes 18 and thereby regulate their expression and subsequent cellular outcome including cell cycle arrest, apoptosis, or senescence. 17 Some anticancer drugs have been confirmed to activate p53 pathway and result in cell cycle arrest, apoptosis, or cellular senescence. 19 –23 The present study's data demonstrate that the apoptosis induced by AECA may be independent of p53, as p53 is mutant in ∼50% of human cancers, and in most of the remaining cancers, the functions of p53 pathway are impaired. 24 Induction of apoptosis by AECA independent of p53 is important for cancer therapy. These observations suggest that AECA as a natural herb may be different from classical chemotherapeutic drugs. In fact, there were no typical side-effects often caused by chemotherapy, such as myelosuppression and digestive response, has been reported in Chinese herb clinical trials. 4,25,26 Whether other Chinese anticancer herbs or herb products are similar to Curcuma aromatica is worthy of further study.
Cyclin B1-CDK1 is the key initiator of mitosis. 27 In cycling cells, cyclin B1 expression increases during late S-early G2 phase, and binding to its inactive partner, CDK1, initiates conformational changes allowing CDK1 to alter its phosphorylation status and to become an active kinase. In early M-phase (mitosis), the active cyclin B1-CDK1 complex translocates to the nucleus to phosphorylate nuclear substrates and allow mitotic onset. 27,28 The present study's data show that AECA induces G2/M phase arrest in LS-174-T cells in coincidence with cyclin B1 and CDK1 downregulation, which suggests that AECA-induced G2/M phase arrest may be related to cyclin B1-CDK1.
Taken together, the present study's observations allow that AECA possesses anticancer activity on LS-174-T cells by inducing both extrinsic and intrinsic apoptosis and induces cell cycle arrest by downregulation of cyclin B1 and CDK1 and independent of p53. Nevertheless, further studies are needed to determine the upstream signal transduction of cell apoptosis and cyclin B1 and CDK1 downregulation and to address which chemical(s) are response for the AECA-induced anticancer effects.
Footnotes
Acknowledgments
This work was partially supported by Key Basic Research Program from Science and Technology Commission of Shanghai Municipality (09JC1413600), Shanghai Natural Science Foundation (07ZR14109), E-institutes of Shanghai Municipal Education Commission (E 03008), and Shanghai Shen Kang Platform Grant (SHDC12007206). The authors also thank Shanghai Key Laboratory of Tissue Engineering for technical assistance.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.