
Abstract
Nonviral approaches have been used extensively for intracellular gene transfer and gene therapy. A modified wheat histone H4 protein, H4TL (H4-TAT-LHRH), as a protein-based gene delivery vector that was able to form stable complexes with plasmid DNA and increase gene delivery efficiency has been described previously. In this study, H4TL has been used to deliver apoptin gene into a human ovarian carcinoma cell line HO8910. After transfection, increased expression of apoptin at both mRNA and protein levels was detected in HO8910 cells, accompanied by reduced rate of growth of HO8910 cells in vitro and the loss of mitochondrial membrane potential in these cells. These data demonstrate that H4TL-mediated transfer of apoptin initiates mitochondrial death pathway in ovarian cancer cells and suggest a novel therapeutic strategy for cancer.
Introduction
The current challenge for gene therapy is to develop a vector that effectively delivers a therapeutic gene (transgene) to targeted cells. The major criterion of the vector is that it can bring the transgene across the plasma membrane and nuclear membrane of target cells and protect the transgene from the degradation by nucleases. In addition, it should not affect the transcription and translation of transgene in selective cells and have no or few detrimental effects. Among nonviral gene delivery systems, natural DNA binding proteins, histones, as the vehicles for gene transfer have been reported by different groups. 1 –5 The DNA binding domains and natural nuclear localization signal sequences make histones excellent candidates for effective gene transfer. 6 Wheat histone H4 is a nucleosome core histone whose gene was cloned and characterized by Tabata et al. 7 There are only two amino acid differences at sites 61 and 78 between human and wheat histone H4, suggesting that wheat histone H4 may be a good carrier for gene transfer. Luteinizing-hormone-releasing hormone (LHRH) can potentially be used as the targeting moiety to deliver anticancer genes specifically to some cancer cells and facilitate their cellular uptake. 8 –10 Protein transduction domain TAT is one of the most well-studied and efficient cell-penetrating peptides. 11 –13
The chicken anemia virus-derived protein, apoptin, is a 14-kDa protein that can selectively induce apoptosis in a wide variety of cancer cells, but not in normal cells. 14 –16 The cancer-selective toxicity of apoptin is associated with its cellular localization. Apoptin is localized in the nuclei of cancerous cells, whereas it remains in the cytoplasm of normal cells. 17 It is proposed that nuclear trafficking and Thr-108 phosphorylation of apoptin in tumor cells are essential for the induction of apoptosis. 18,19 The proapoptotic signal of apoptin is transduced from the nucleus to the cytoplasm by Nur77, which activates apoptosis by a p53-independent mitochondrial death pathway. 20,21
In the present study, LHRH and TAT have been genetically incorporated with the wheat histone H4 to enhance the penetration of H4 across the plasma membrane and reach the targeted LHRH receptor-positive cells. H4TL (H4-TAT-LHRH) has been used with naked plasmid DNA that contains an apoptin tumor-selective killer gene to evaluate the transfection efficiency and antitumor activity in human ovarian carcinoma HO8910 cells.
Materials and Methods
Preparation of recombinant protein H4TL
Recombinant protein H4TL was expressed in Escherichia coli. In brief, E. coli BL21 (DE3) containing the H4TL recombinant plasmid (constructed in our lab) was grown at 37°C in LB medium with 50 μg mL−1 kanamycin till OD600 was ∼0.4, and then induced by 1 mM isopropyl-β-D-thiogalactopyranoside at 30°C for 10 hours. The cells were collected by centrifugation at 7000 g at 4°C for 15 minutes, washed twice with phosphate-buffered saline (PBS), and lysed by sonication. H4TL was purified by one-step affinity chromatography with nickel-Sepharose in the presence of 6 M guanidine hydrochloride, and the concentration of purified H4TL was determined by a SmartSpec™ Plus spectrophotometer (Bio-Rad).
Construction of plasmids pcDNA3.1/apoptin (p-apoptin)
The wild-type apoptin gene was amplified by polymerase chain reaction (PCR) with PET-28a-apoptin (a construct made in our lab) as the template, and the following primers were synthesized by Sangon BioTech: 5′-tacggtaccatgggaaacgctctccaag-3′ and 5′-ttgctcgagttacagtcttatacacctt-3′. The restriction site of KpnI meets the requirements for the Kozak translation initiation sequence [(G/A)NNATGG]. The PCR conditions were as follows: 5 minutes at 94°C, 30 cycles at 94°C for 30 seconds, annealing at 56°C for 45 seconds, and extension at 72°C for 1 minute followed by 1 cycle of 72°C for 10 minutes. Purified PCR product was digested using KpnI/XhoI and subcloned into pcDNA3.1 to make pcDNA3.1/apoptin (p-apoptin). Correct constructs were confirmed by sequencing (Sangon BioTech).
Cell culture and transfection
The 23rd generation of the human fetal lung diploid cell line, 2BS, and the human ovarian cancer cell line, HO8910, were seeded on a six-well plate at 2 × 106 cells/well. Cells were grown in Dulbecco's modified Eagle's medium/Ham's F-12 (Gibco BRL), supplemented with 10% (v/v) fetal bovine serum (Gibco BRL), 1% antibiotic–antimycotic (Invitrogen), and 2.5 mM
Reverse transcriptase–PCR
Total RNA was isolated from the transfected cells using Trizol® Reagent (Invitrogen) and first-strand cDNA was synthesized using 1 μL purified RNA, 0.5 μL AMV reverse transcriptase (RT) (TaKaRa), 1 μL 10× RT buffer, 2 μL MgCl2, 1 μL dNTP mixture, 0.5 μL Oligo-dT-Adopter-Primer, and 4 μL RNase-free water. The condition of RT was 42°C for 30 minutes followed by 99°C for 5 minutes. PCR was performed using the primers for apoptin (5′-tacggtaccatgggaaacgctctccaag-3′ and 5′-ttgctcgagttacagtcttatacacctt-3′) and β-actin (5′-ggatcagcaagcaggagtatg-3′ and 5′-caccttcaccgttccagttt-3′). The condition of PCR was as follows: 94°C for 3 minutes (1 cycle), 94°C for 45 seconds, 55°C for 45 seconds, and 72°C for 1 minute (30 cycles).
Western blotting analysis
The transfected cells were washed twice with PBS and lysed in 100 μL protein sample loading buffer (50 mM Tris pH 6.8, 2% sodium dodecyl sulfate [SDS], 0.1% BPB, 10% glycerol, and 1% β-mercaptoethanol). Ten (10) microliters of the prepared sample was loaded onto a 12% SDS–polyacrylamide gel electrophoresis gel and was transferred to a polyvinylidene fluoride membrane (Amersham Pharmacia Biotech). The transferred membrane was blocked with 5% dried milk in TBS plus 0.1% Tween 20 (block buffer) for 1 hour at room temperature and subjected to immunoblot analysis with apoptin and β-actin antibodies (Santa Cruz) diluted 1:1000 in block buffer. The polyvinylidene fluoride membrane was developed using Immobilon Chemiluminescent Western HRP Substrate (Millipore).
MTT assay
Cells were seeded on 96-well plates at 5 × 103 cells/well. After 24 hours, the cells were treated with three different agents (100 μL) as follows: (1) pcDNA3.1 (as the control); (2) H4TL alone; (3) H4TL/p-apoptin. Next, 10 μL of 5 mg/mL MTT dissolved in PBS was added to the cells in 100 μL of medium and incubated at 37°C for 4 hours under 5% CO2, followed by removal of the medium and the addition of 100 μL of lysis buffer (99.4% v/v dimethylsulfoxide, 0.6% v/v acetic acid, and 10% w/v SDS). Absorbance values, measured at 490 nm, were expressed as a percentage of untreated controls.
Apoptosis assay
Apoptosis assay was performed by flow cytometry (FACSCalibur; Becton Dickinson) using an annexin V/propidium iodide double-labeling assay kit (KeyGene). Briefly, HO8910 cells were cultured on a six-well plate and allowed to grow for 24 hours. Next, the cells were transfected with Lipofectamine 2000/pcDNA3.1, H4TL, H4TL/p-apoptin complexes, or Lipofectamine 2000/p-apoptin (L/p-apoptin) complexes, respectively. 2BS cells were used as controls. Forty-eight (48) hours after transfection, both attached and floating cells were collected and washed twice with PBS. The cells were then stained with the annexin V–fluorescein isothiocyanate and propidium iodide. The samples were analyzed using a flow cytometer.
Analysis of genomic DNA from apoptotic cells
More than 1 × 106 cells were collected in 1.5-mL tubes by centrifugation at 1000 g for 5 minutes, resuspended in 100 μL 5× TBE buffer (445 mM Tris–boric acid and 10 mM ethylenediaminetetraacetic acid) and incubated for 30 minutes. Proteinase K (1 mg/mL) was added for incubation at 37°C for an additional hour. Lysates were then extracted once with phenol-chloroform and DNA was precipitated with 2.5 volumes of 100% ethanol overnight at −20°C. DNA was collected by centrifugation at 14,000 rpm for 20 minutes, resuspended in 10 mM Tris buffered with 1 mM ethylenediaminetetraacetic acid plus 0.1 mg/mL RNase A, and incubated at 37°C for 1 hour. DNA was then electrophoresed in 1.0% agarose gels containing ethidium bromide.
Measurement of mitochondrial membrane potential
Mitochondrial permeability transition was determined by staining the cells with a rhodamine123 kit (KeyGene), as described previously. 22,23 The mitochondrial membrane potential was quantified by flow cytometric determination of cells with increased fluorescence in the FL-2 channel. Data were given as percent cells with green fluorescence due to the release of rhodamine 123, which reflects the percentage of cells losing the mitochondrial membrane potential.
Statistical analysis
All data are expressed as the mean ± standard error of the mean. Statistical significance was evaluated by the Student's t-test, and p-values <0.05 were considered significant. Statistical analysis was done using the GraphPad software program.
Results
H4TL-mediated efficient apoptin gene transfer into HO8910 and 2BS cells
H4TL-mediated apoptin gene transfer into the cells was evaluated by RT-PCR and Western blot analysis. As shown in Figure 1A, compared with H4TL or pcDNA3.1-treated 2BS and HO8910 cells, apoptin mRNA expression were only detected in H4TL/p-apoptin or L/p-apoptin-transfected cells, into which 2 μg p-apoptin was delivered by 8–10 μg H4TL or by Lipofectamine. In addition, Western blotting analysis confirmed H4TL-mediated expression of apoptin at a protein level in both 2BS and HO8910 cells (Fig. 1B). The results indicate that H4TL can transport apoptin vector into the nuclei efficiently and facilitate the exogenous expression of apoptin.

Expression of apoptin as detected by reverse transcriptase–polymerase chain reaction and Western blotting analysis.
H4TL-mediated apoptin gene transfer inhibits the proliferation of HO8910 cells
To investigate the cellular effects resulting from H4TL-mediated apoptin gene transfer, MTT assay was performed after incubation of HO8910 and 2BS with H4TL/p-apoptin mixture. Whereas apoptin transported with H4TL had negligible effects on cell proliferation in 2BS cells (Fig. 2A), the proliferation of HO8910 incubated with H4TL/p-apoptin mixture was inhibited significantly in a time-dependent manner, compared with control group of H4TL treated or pcDNA transfected (Fig. 2B). These data demonstrate that H4TL-mediated apoptin gene transfer did not interfere with the specific function of apoptin in cancer cells.

Proliferation of 2BS ) treated with 0.2 μg/mL H4TL; (▪) 5 ng p-apoptin was transfected with 20 ng H4TL per well at 12, 24, or 48 hours. The results were based on three experiments and presented as means ± standard error of the mean (SEM). p < 0.05, n = 8.
H4TL-mediated apoptin gene transfer induces the apoptosis of HO8910 cells
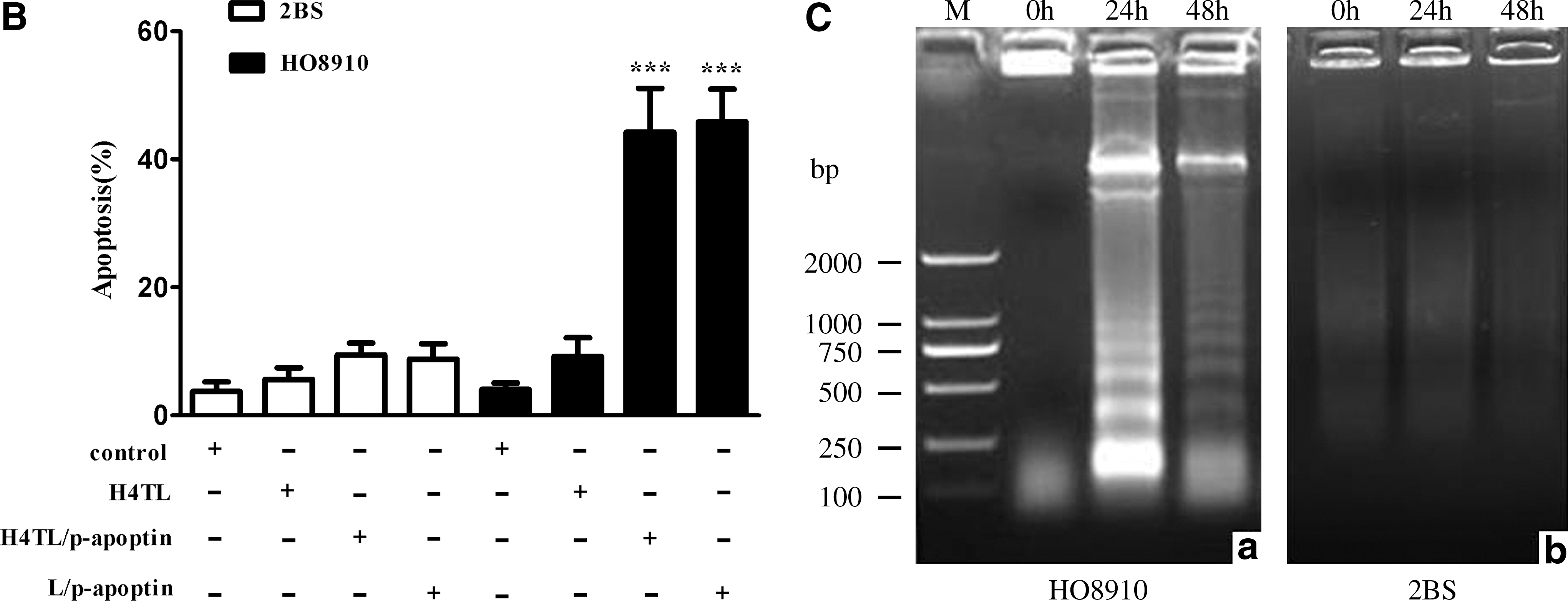
Since apoptin can selectively induce apoptosis in a wide variety of cancer cells, next HO8910 and 2BS cells were exposed to H4TL/p-apoptin mixture and performed apoptosis assay. As shown in Figure 3A, compared with the control groups, H4TL or lipofectamine-mediated transfer of p-apoptin caused cell death in HO8910 cells specifically. The result was further confirmed by annexin V apoptosis assay (Fig. 3B). Noteworthy characteristics of apoptosis are the activation of endonucleases leading to the fragmentation of the genomic DNA and the loss of mitochondrial membrane potential. Thus, these aspects were examined and it was found that H4TL-mediated apoptin gene transfer induced prominent DNA laddering (Fig. 3C) and significant mitochondrial membrane potential loss (Fig. 3D) in HO8910 cells but not in 2BS cells. Collectively, these data demonstrate that apoptin gene transferred by H4TL can effectively and specifically induce apoptosis in HO8910 cancer cells.
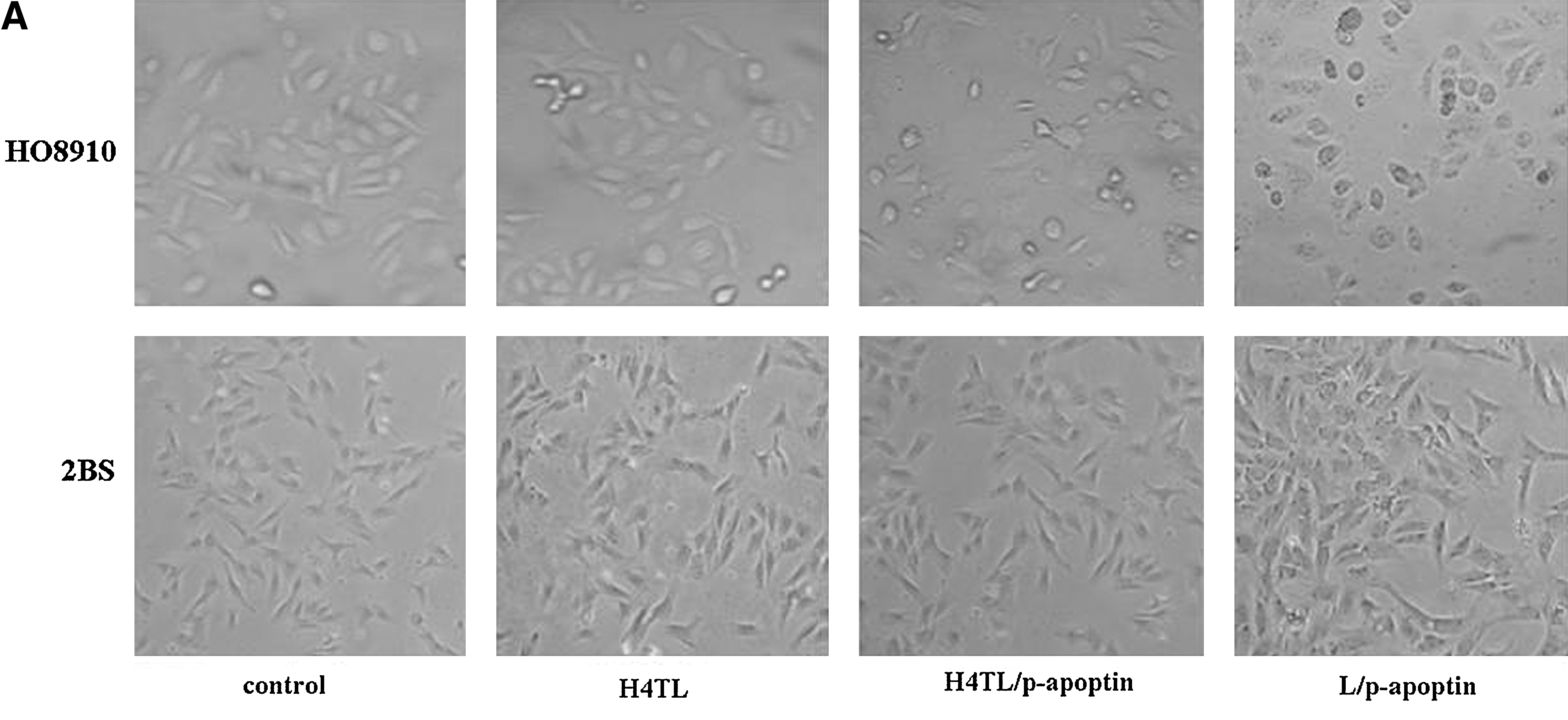
Apoptosis of 2BS and HO8910 cells 48 hours after transfection of apoptin gene via different carriers.

Discussion
Over the past few years, several reports have suggested that histones can efficiently mediate gene transfer due to their ability to bind DNA (histonefection). Several members of the histone family have been demonstrated to be effective in histonefection. H4TL, a recombinant wheat histone H4 protein, shares the properties of eukaryote histones, including small size, basic nature, the toroidal wrapping of DNA, protection of DNA from nuclease digestion, and the formation of nucleosome-like structures. More importantly, H4TL incorporates the peptide of PDT-TAT and LHRH, and thus can directly cross the cell plasma membrane without disrupting the membrane integrity via a receptor and endocytic-independent pathway.
Another important advantage of H4TL-mediated gene delivery is its low cytotoxicity. MTT assay was employed to characterize the cytotoxic effects of fusion protein-based transfection. The fusion protein, H4TL, was nontoxic to normal cells such as 2BS and may be used over a wide range of incubation periods as a highly efficient gene carrier (Fig. 2). Microscopic examination of cells treated with Lipofectamine revealed gross morphological alteration, cell disruption, and a reduction in viability, whereas the H4TL was less toxic and better tolerated than Lipofectamine-based transfection reagents. Combined with the advantages of higher transfection efficiency and lower toxicity, H4TL may serve as a good alternative tool for gene delivery. Puebla et al. 2 reported that H1 was less toxic and better tolerated than the liposome-based transfection reagents. The absence of toxicity of H1 and histone-like proteins has also been described by other groups. 24,25 The recombinant polypeptide that has been developed contains wheat histone H4 that has two amino acids different from human histone H4. When H4TL is repeatedly used for DNA transfection in vivo, this exogenous protein may induce immunity reaction. In addition, LHRH fused to H4TL may have unexpected hormonal activity. Further in vivo study is vital to as the potential side effects of H4TL gene delivery system.
The cancer-selective toxicity of apoptin has attracted great attention because apoptin targets the molecular basis of the transformed phenotype. Targeted cancer therapies interfere with malignant cell growth and division at various points during the initiation, development, and metastasis of cancer. The mitochondrial/apoptosome death pathway is ultimately activated by a number of anticancer drugs and stress stimuli. 26,27 These stimuli frequently cause the loss of mitochondrial membrane potential, and the release of cytochrome c, AIF, and other molecules. 28 –32 Therefore, the loss of mitochondrial membrane potential was detected by flow cytometric analysis and confirmed that apoptin transferred via H4TL vector could trigger mitochondrial death pathway to execute its inhibitory effects on cancer cells. Our next step is to translate these in vitro observations into in vivo intraperitoneal model of ovarian cancer to evaluate the therapeutic effects of H4TL-mediated transfer of apoptin gene.
In conclusion, a protein-based, nonviral DNA delivery vector for cancer-specific gene therapy was developed. The fusion protein, H4TL, shows high transfection efficiency in apoptin gene transfer and is nontoxic to normal cells. The use of peptides to overcome the cellular barriers to DNA entry into the cell and deliver the DNA into the nucleus is a strategy for the nonviral gene vector. Induction of apoptosis and loss of mitochondrial membrane potential in HO8910 cells transfected with H4TL/apoptin vector demonstrate that H4TL-mediated transfer of apoptin initiates mitochondrial death pathway in ovarian cancer cells and suggests a novel therapeutic strategy for cancer.
Footnotes
Acknowledgments
This study was supported by the National Science Foundation (Grant nos. 30471248, 30871850). The authors are grateful to Dr. George Ye for providing HO8910 and 2BS cell lines. The authors gratefully acknowledge Dr. JiMing Wang for critical reading of the article and helpful suggestions.
Disclosure Statement
No competing financial interests exist.