
Abstract
Background:
Silymarin, the main flavonoid constituent element extracted from Silybum marianum possessing antioxidant activity, is already known to be able to block the NF-κB activation process and result in cell apoptosis, implicating silymarin's potential to control cancer cell growth.
Materials and Methods:
In this study, based upon the above assumption, silymarin was administered to a highly metastatic lung cancer cell line Anip973 to test silymarin's role in cancer cell proliferation.
Results:
Silymarin had significant inhibitory effects on the proliferation of Anip973 cells in a dose-dependent and time–response manner within 48 hours. Silymarin can induce Anip973 apoptosis.
Conclusions:
Silymarin may in vitro inhibit the proliferation of the human lung adenocarcinoma cell line Anip973 and induce apoptosis via the mitochondria-dependent caspase cascade pathway.
Introduction
L
Based upon the above knowledge about pulmonary carcinogenesis, there is certainly a plausible need to employ it in the compelling treatment of lung cancer. New chemotherapy drugs, particularly molecular agents with novel mechanisms of action, are now in clinical testing and may improve therapeutic options. 17 Rapid developments in biotechnology and increased knowledge about important signaling pathways leading to cell proliferation, apoptosis, angiogenesis, and metastases, new treatment possibilities with specific targeted therapies have emerged for chemoprevention and treatment of lung cancer and had been reviewed in details. 18 Several studies have evaluated the prognostic and therapeutic implications of abnormalities in proliferation and apoptosis in lung cancer, and the results showed that markers of increased proliferation and decreased cell death were associated with poor prognosis. 19 Among the few known apoptotic markers are the Fas receptor, Apo2ligand/tumor necrosis factor-related apoptosis-inducing ligand, peroxisome proliferator-activated receptor, Bax, and Survivin. Though there are different pathways to apoptosis, many of these share common elements of activating cytochrome c in mitochondria with subsequent activation of caspase-3 leading to apoptosis through protein degradation. The prognostic implication of expression of caspases had been also evaluated, and some studies have demonstrated better prognosis for patients having caspase-positive tumors. 20
Silymarin, a crude seed extract complex of silibinin isolated from the fruits of milk thistle, Silybum marianum, has high human acceptance and thus been initially used as an antihepatoxic agent as well as dietary supplement around the world. 21,22 It is a polyphenolic flavonoid and flavone composed mainly of silibinin with small amounts of other silibinin stereoisomers, namely, isosilybin, dihydrosilybin, silydianin, and silychristin. 23 Silymarin had displayed a fairly strong antioxidant activity as well as many other pharmacological properties, such as antagonizing hepatic toxicity arising from many chemicals such as carbon tetrachloride, galactosamine, and ethanol. 24,25 During the last decade, a wide range of studies have shown a strong cancer chemopreventive and anticancer efficacy of silymarin (or silibinin) in many kinds of tumor. 26 –29 But until now, to the authors' best knowledge, little is known about the effects of silymarin on human lung cancer cells. Here, silymarin's effects on proliferation, cell cycle distribution, and apoptosis and its possible mechanism were assessed using a highly metastatic human lung cancer cell line Anip973, hoping to preview the perspective of silymarin application for lung cancer therapy.
Materials and Methods
Cell lines and culture
A highly metastatic lung cancer cell line Anip973 was purchased from Cell Preservation Center, Shanghai Institute of Biochemistry and Cell Biology, Chinese Academy of Science. The cells were cultured at 37°C in a 5% CO2 humidity-saturated incubator in 25-mL glass flasks containing RPMI 1640 culture medium supplemented with fetal bovine serum (FBS 10%, v/v), 100 U/mL penicillin, and 100 μg/mL streptomycin.
Reagents and kits
Silymarin was purchased from National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China), dissolved with dimethyl sulfoxide (DMSO) under sterile conditions, and diluted to desired concentrations with RPMI 1640 culture medium. Ethidium bromide, RNase A, Protease K, and 3-[4,5-diamethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT) were from Sigma-Aldrich Chemical Co. (St. Louis, MO). NAD was purchased from Shanghai Chemical Reagents Company (Shanghai, China). Caspase-9 and caspase-3 activity detection kits were purchased from Chemicon International (Temecula, CA). Polyclonal antibodies against Bcl-2 and Bax were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Purified mouse anti-human poly ADP-ribose polymerase (PARP) monoclonal antibody was obtained from BD Biosciences Pharmingen (San Jose, CA). RPMI 1640 culture medium, FBS, and trypsin were purchased from GIBCO®, Invitrogen Corporation (Carlsbad, CA). DMSO was product distributed by Sigma-Aldrich Chemical Co. Horseradish peroxidase (HRP)-labeled goat anti-rabbit and goat anti-mouse IgG were purchased from Boster Bioengineering, Inc. (Wuhan, China). ECL detection system kit and BCA protein quantification kit were from Pierce Biotechnology, Inc. (Rockford, IL). Phenylmethylsulfonyl fluoride was from Shanghai Chemical Reagents Company (Shanghai, China), and nitrocellulose was from Amersham Plc. (Buckinghamshire, UK).
Effects of silymarin on Anip973 cell proliferation by MTT assay
Cells in log-phase growth were digested by 0.25% trypsin, suspended in 10% FBS, and cultured in 96-well culture plates starting from 1.5 × 104 cells/mL per well (100 μL/well) at 37°C in 5% CO2. After 24-hour culture, silymarin in 100 μL RPMI 1640 culture medium was added to every culture well, reaching a final silymarin concentration of 5, 10, 15, 20, 40, and 60 μg/mL. Also, RPMI 1640 culture medium containing only DMSO was set as negative control, and only culture medium without any cells was set as blank control. All testing and control groups were repeated in triplicate. At 24, 48, and 72 hours of culture, 20 μL MTT (5 mg/mL) was added to every well, and culturing was continued for 4 hours. Then, culture supernatant was discarded and replaced by 150 μL DMSO, and the cell plates were shaken for 10 minutes. Finally, the absorbance of every culture well was read on an ELISA Reader at 490 nm.
Morphological changes observed under an inverted microscope
Anip973 cancer cells in log-phase growth were subject to 0.25% trypsin digestion, resuspended into RPMI 1640 culture medium containing 10% FBS adjusting cell concentration at 1.5 × 104/mL, and distributed into 96-well culture plates for culture (100 μL per well). After 24 hours, to the cell culture was added 100 μL RPMI 1640 culture medium per well containing silymarin at varying concentrations (0, 10, 20, and 40 μg/mL). After 48 hours, the cancer cell culture plates were observed under an inverted microscope and photographed.
Cellular ultrastructural observation under a scanning microscope
After 48 hours of culture under 20 μg/mL silymarin treatment, Anip973 cells were digested by 0.25% trypsin, suspended in RPMI 1640 culture medium supplemented with 10% FBS, transferred to a 10-mL glass centrifuge tube, and centrifugated at 1000 rpm for 5 minutes. The supernatants were discarded, and the cell pellets were resuspended in 1 mL fresh culture medium, transferred to a 1.5-mL Eppendorf plastic tube, and centrifugated at 1500 rpm for 15 minutes. After the supernatants were pipetted out, 4°C precold 0.25% glutaradehyde fixative was slowly added into cell pellets along the tube wall and finally observed under an electron microscope after the cell specimen embedding and slicing.
Cell cycle detection by FCM
Anip973 cells were treated with 20 μg/mL silymarin for 24 and 48 hours, digested with 0.25% trypsin, and centrifugated at 500 rpm for 5 minutes, and the pellets were washed with phosphate-buffered saline (PBS) twice. The pellets were resuspended with 0.9 mL PBS, and 2.1 mL pure ethanol (final concentration at 70%) was slowly added for fixation at 4°C overnight. Then, fixed cells were centrifugated, washed twice with PBS, resuspended in 100 μL PBS, added with 300 μL propidium iodine for 15 minutes, and finally assayed by flow cytometry (FCM) for cell cycle distribution (G1, G2, and S phases).
Apoptosis detection by FCM
Anip973 cells were treated with desired doses of silymarin at 10, 20, and 40 μg/mL concentration in complete medium for 48 hours. At the end of treatment time, the cells were collected after a brief incubation with 0.25% trypsin and centrifugation at 500 rpm for 5 minutes. After washing with PBS twice, cell cycle distribution was analyzed by FCM using an Annexin V–FITC staining kit according to manufacturer's instruction.
Apoptosis detection by DNA ladder
Anip973 cells were plated for varying times (0, 6, 24, and 48 hours) under 40 μg/mL silymarin, harvested, lysed by lysis buffer (10 mM Tris-Cl [pH 7.6], 10 mM EDTA [pH 8.0], and 0.5% Triton X-100), and centrifugated at 13,000 rpm for 5 minutes. The supernatants were collected and successively incubated with 1% SDS and RNase A (5 mg/mL) at 56°C for 2 hours and protease K (2.5 mg/mL) at 37°C for 2 hours. DNA sediments were then precipitated by 1/10 volume 3 M sodium acetate and 2.5 volume cold ethanol, incubated overnight at 4°C, and centrifugated at 14,000 rpm for 15 minutes. Finally, the precipitates were dissolved in 50 μL TE buffer, electrophoresed in 1.8% agarose gel with DNA loading buffer, and photographed with ethidium bromide staining.
Apoptosis molecule PARP detected by Western blot
Anip973 cells were cultured with or without varying silymarin concentration (0, 5, 10, 15, 20, 40, and 60 μg/mL) at varying times (0, 6, 12, 18, 24, and 48 hours). After PBS washing, the cells were lysed in ice with addition of appropriate RIPA lysis buffer and subjected to BCA assay for protein quantitative analysis. Cell protein samples with equal amount of total proteins were subjected to 9% SDS-PAGE and transferred to a nitrocellulose membrane. Finally, the membrane was incubated with mouse anti-human PARP monoclonal antibodies as primary antibody and HRP-labeled goat anti-mouse IgG as secondary antibody and then exposed to films.
Bax and Bcl-2 expression detected by Western blot after silymarin administration
Anip973 cells were plated for 24 hours with varying silymarin concentrations (0, 5, 10, 15, 20, 40, and 60 μg/mL), harvested, and subjected to intracellular Bax and Bcl-2 detection by Western blotting. Except for rabbit anti-human Bax polyclonal antibodies and mouse anti-human Bcl-2 monoclonal antibody as primary antibodies and HRP-labeled goat anti-rabbit and goat anti-mouse IgG as secondary antibodies, the remaining procedures were the same as described earlier.
Activity detection of caspase-3 and caspase-9
A total of 2 × 105 Anip973 cells were plated in RPMI l640 medium (containing 10% FBS, penicillin 100 U/mL, and streptomycin 100 μg/mL) at 37°C in 5% CO2 with full humidity, treated with silymarin at 40 μg/mL, and harvested at times 6, 12, 18, 24, and 48 hours. The collected cells were lysed in 500 μL cell lysis buffer on ice for 10 minutes and centrifugated at 10,000 g for 1 minute at 4°C. The supernatants were pipetted out and mixed with 2 volume reaction buffer (containing 10 mM DTF) followed by addition of 5 mL of 4 mM LEHT-ONA as the substrate, incubated at 37°C for 2 hours, and read on EL312 (BioTek Instruments, Inc., Winooski, VT) at wavelength 400–405 nm.
Results
Silymarin shows inhibitory effects on proliferation of a human highly metastatic lung cancer cell Anip973
To examine the effects of silymarin on lung cancer, Anip973 cells were treated with various concentrations of (5, 10, 15, 20, and 40 μg/mL) silymarin for indicated times (24, 48, and 72 hours). MTT assay was used to stain viable cancer cells for assessing silymarin inhibition on Anip973 cells. The inhibitory effects of silymarin (demonstrated as cancer cell viability rates) were clearly seen at all times of 24-, 48-, and 72-hour treatments in a dose-dependent manner (Fig. 1). Because the inhibitory effect of silymarin on Anip973 cells was significant at the 48 hours time point, the IC50 was calculated as 18.6 μg/mL. So, the 20 μg/mL concentration was used for following experiments.

Inhibitory effect of silymarin on Anip973 cell proliferation by MTT assay. In a time–response study, the inhibitory effect of silymarin on Anip973 cells was evident at 24, 48, and 72 hours post-treatment at 5, 10, 15, 20, 40, and 60 μg/mL. MTT, 3-[4,5-diamethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide.
Morphological changes of Anip973 cells after silymarin treatment under microscopes
On treatment with 0, 10, 20, and 40 μg/mL silymarin for 48 hours, the count number of viable Anip973 cells was found to be decreased under an inverted light microscope, with some cells being much shrunk and blebbed. When silymarin concentration was increased, however, it was found that some cancer cells appeared larger, whereas more cells were dead, especially at 48 hours after 40 μg/mL silymarin treatment. In the negative controls with only commensurate solvent in RPMI 1640, Anip973 cells grew fairly well in cell number and density (Fig. 2A). Under a scanning electron microscope, it was observed that nonsilymarin-treated Anip973 cancer cells sticked to plate walls tightly and grew well with numerous micropili stretched out at cell surface. After 20 μg/mL silymarin treatment for 48 hours, Anip973 cells were seen to be swelled with micropili loss, looking much smooth, and apoptosis bodies were formed. At a later time, apoptosis bodies were found to be released and scattered around cells in the form of clumps (Fig. 2B).
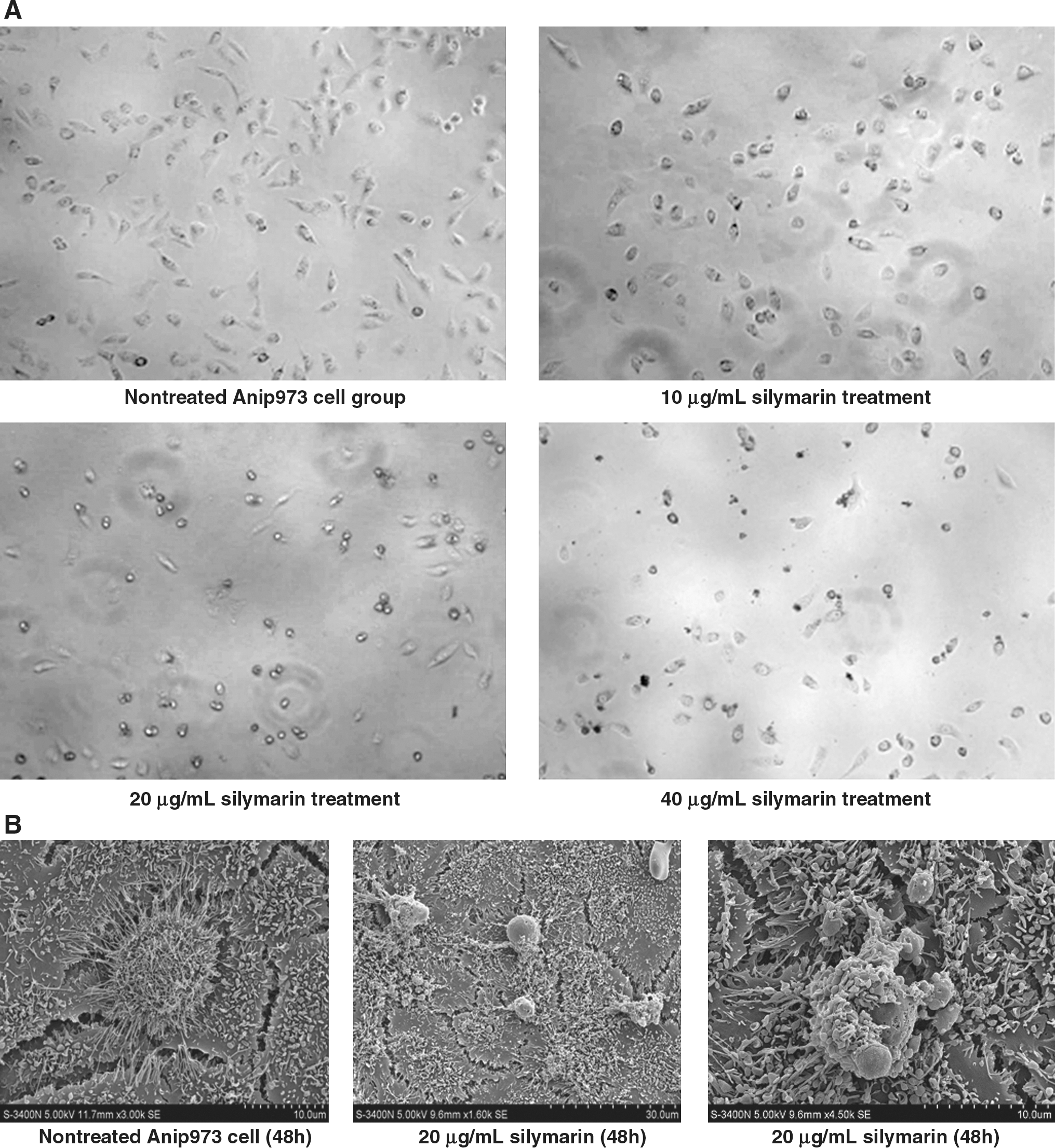
Morphological changes of Anip973 cells after silymarin treatment.
Silymarin induces a strong G1 arrest of Anip973 cell cycle distribution
Further, having witnessed Anip973 cell death after administration of silymarin, to know whether the growth inhibitory effect of silymarin on Anip973 cells is accompanied by cell cycle arrest, silymarin-specific effects on cell cycle distribution were examined by FCM. After treatment with 20 μg/mL silymarin for 24 and 48 hours, silymarin caused a strong time-dependent increase in G1 phase together with a concomitant remarkable decrease in S phase and a slight G2 decrease in population when compared with negative control, indicating a strong G1 arrest. Silymarin treatment at 20 μg/mL dose for 24 hours caused 67.0% (p < 0.05) cells in G1 phase when compared with negative control showing 40.8%. Conversely, S-phase cell population was decreased to 27.2% (p < 0.05) when compared with control having 50.2%. A similar increase in G1-phase cell population and a decrease in S phase were also observed with 20 μg/mL silymarin dose for 48 hours (Table 1). Overall, these results suggested the role of cell cycle arrest in silymarin-induced growth inhibition of human lung cancer cells.
A, negative control; B, 20 μg/mL silymarin treatment after 24 hours; C, 20 μg/mL silymarin treatment after 48 hours.
p < 0.05.
Silymarin induces Anip973 cell apoptosis
Furthermore, as silymarin exhibited an inhibitory effect on Anip973 cell proliferation, whether silymarin induced Anip973 apoptosis was examined. At 24 hours post-treatment of silymarin, the percentage of apoptic cells in total cell population was 9.0%, 17.5%, and 21.5%, respectively, of dosage compared with the negative control showing 2.8% (p < 0.01) in a dose-dependent manner (Fig. 3A). The apoptosis was also verified by DNA ladder display. On treatment with 20 μg/mL silymarin, Anip973 cells were harvested at 0, 6, 24, and 48 hours and then subjected to agarose electrophoresis to demonstrate the distinct status of the cellular genomic DNA. Notable apoptosis was observed ever after 24 hours of silymarin treatment, especially the strong ladder bands between 180 and 200 bp (Fig. 3B).

Silymarin induces Anip973 cell apoptosis.
To verify the cancer cell apoptosis induced by silymarin further, western immunoblotting was used to manifest definite silymarin effects on Bcl-2/Bax expression. The results showed that at 24 hours postadministration of the drug, silymarin rendered a prominent enhancement of Bax expression and downregulation of Bcl-2 expression in a dose-dependent manner, indicating a proapoptosis effect triggered by silymarin (Fig. 3C).
Silymarin causes caspase-dependent apoptosis
The levels of caspase-3 and caspase-9 activity in Anip973 cell lysates after various times of 40 μg/mL silymarin treatment were investigated. The test results demonstrated that the activities of caspase-3 and caspase-9 at 48 hours after silymarin application had prominently increased by 7.6- and 10.2-fold, respectively, compared with 0 hour of silymarin treatment (Fig. 4A). Next, the effect of silymarin-caused caspase activation on PARP cleavage was examined, which is one of the downstream substrates of caspase cascade and a reliable marker of apoptosis. By western immunoblotting, it was found that at 12 hours postsilymarin treatment, the specific 89-kDa fragment emerged under 10 μg/mL and higher silymarin concentration in a dose-dependent manner (Fig. 4B). When Anip973 cells were treated with 10 μg/mL silymarin, the 89-kDa fragment appeared as early as at 6 hours post-treatment (Fig. 4B). Overall, these results suggest the involvement of caspase-dependent mechanism in silymarin-induced apoptosis of lung cancer cells.
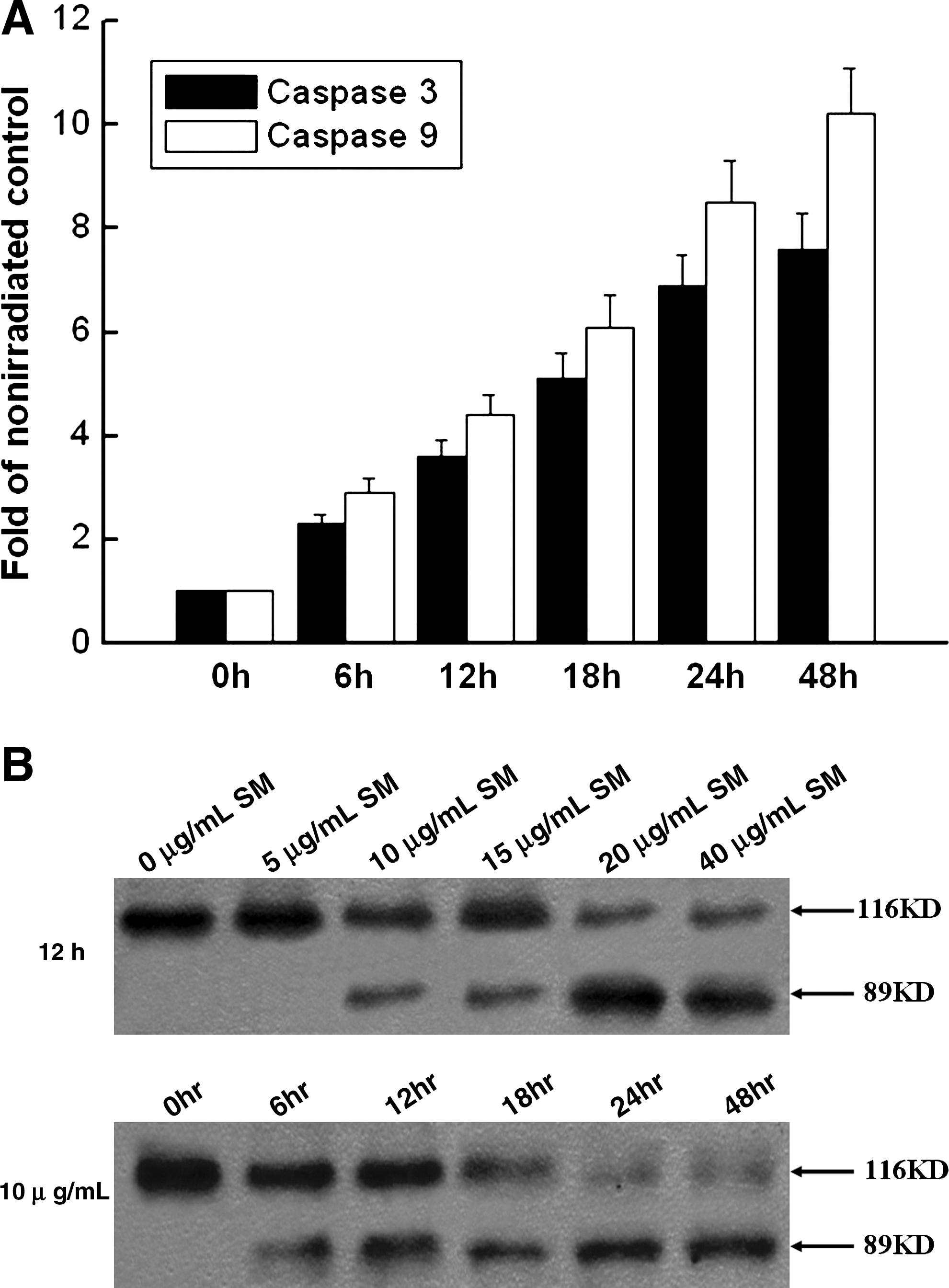
Silymarin causes caspase-dependent apoptosis.
Discussion
Silymarin is a polyphenolic flavonoid compound isolated mainly from the fruits or seeds of milk thistle (Silybum marianum). It has been used as dietary supplement and clinical drug with high acceptability for few decades in Europe, Asia, and the United States. Silymarin is well known for its hepatoprotective activity as well as other pharmaceutical properties, such as antioxidant activity, free radical elimination, 5-lipoxygenase inhibition, maintenance of cellular membrane stability, hepatocyte regeneration, and hypocholesterolemia. 30,31 In the last decade, several studies have suggested that silymarin (also including silibinin) possesses anticancer potential against many cancers. 32 –34 However, its mechanisms of action are not known completely, although silymarin may act upon through promoting tumor cell apoptosis and differentiation, suppressing antiapoptotic molecule survivin expression, phosphorylation of kinases ERK1/2 and JNK1/2, and tumor angiogenesis, and promoting tumor necrosis factor-α-induced apoptosis through suppressing NF-κB activation, caspase activation, and PARP cleavage. 35 To the authors' best knowledge, there are few reports about silymarin's effects on lung cancer; these research showed that silibinin can inhibit the growth of lung cancer through cell-cycle arrest 36 or downregulate inducible nitric oxide synthase. 37 Metastasis is the basic biological feature of malignant tumor cells, especially lung adenocarcinoma, and the major cause of deaths from lung tumor. Anip973 manifested much higher metastatic potential. In the present study, the anticancer efficacy targeting apoptosis and cell cycle regulation and the associated molecular mechanism in a highly metastatic human lung cancer cell line Anip973 were assessed.
As unchecked proliferative potential involving deregulation in cell cycle progression is generally described as a central process in cancer development, one of the focusing strategies in cancer management is to target the limitless replication characteristics and deregulate cell cycle progression in cancer cells. 38,39 In this regard, the findings of the present study suggest that silymarin exerts strong antiproliferative effects against human lung cancer Anip973 cells in a dose-dependent manner and inverse proportion in a time response and that this effect involves alterations in cell cycle regulators, causing a strong G1 arrest as well as cell death (apoptosis), all confirmed by corresponding morphological changes under light and electron microscopes. These observations again unsurprisingly validate the definite anticancer efficacy of silymarin, consistent with numerous previous studies mentioned earlier.
In the present study, silymarin induced Anip973 apoptosis, and western immunoblotting results showed a great suppression of Bcl-2 (antiapoptotic) expression and remarkable elevation of Bax (pro-apoptotic) expression, suggesting that an intrinsic pathway was activated, which may involve cytochrome c release from mitochondria.
One important thing to be noted here is the concentration of silymarin. An ongoing phase I–II clinical trial with silibinin (main constituent of silymarin with similar biological activities) in prostate cancer patients, aiming to analyze serum silibinin levels and any associated biological response, has shown that up to 74 μg/mL total silibinin levels can be achieved in the plasma without any sign of gross toxicity in silibinin-treated groups of animals versus controls. 40 The maximum recommended human dose of silymarin is 7 mg/kg body weight according to FDA criteria. Another pilot study of oral silibinin in colorectal cancer patients has shown that silibinin level in plasma achieved 0.3–4 μM with repeated oral administration at dosages of 360, 720, or 1440 mg silibinin daily for a week. 41 Taken together, these results clearly indicate that the silymarin concentrations used in the present study (the highest at 60 μg/mL) showing strong efficacy in human lung cancer cells Anip973 are pharmacologically achievable without any toxicity at least in the rodent studies completed.
Conclusions
To summarize, the present study shows that silymarin, a naturally occurring polyphenolic agent from milk thistle, could strongly inhibit the proliferation of human lung cancer cells Anip973. It was also shown that silymarin caused caspases activation and PARP cleavage identified by western blotting as well as apoptosis induction confirmed by AV/PI staining followed by FACS analysis. As evidenced from the results, silymarin may be a powerful candidate in the development of therapeutic agents for lung cancer therapy.
Disclosure Statement
There were no financial conflicts as required by law.