
Abstract
To investigate the effects of thalidomide on the radiosensitivity of human esophageal cancer cells (TE1 cells) and the potential mechanism underlying these effects. The effects of thalidomide on proliferation of TE1 cells were determined by Methyl thiazolyl tetrazolium assay. The multitarget click model was used to delineate the survival curve using a colony-forming assay, and the radiosensitivity was determined after TE1 cells were treated by thalidomide and/or X-ray radiation. The cell cycle was detected using flow cytometry. Our results are as follows: thalidomide alone suppressed the proliferation of TE1 cells in a dose- and time-dependent manner. The suppressive effects were enhanced by prolonged duration or elevated concentration of thalidomide. However, thalidomide did not affect the cell cycle of TE1 cells. The expression of vascular endothelial growth factor (VEGF) mRNA and protein was suppressed after treatment with thalidomide alone in a dose-dependent manner. Synergistic suppressive effects on VEGF expression were observed after administration of thalidomide and X-ray exposure. In conclusion, thalidomide was able to enhance the radiosensitivity of TE1 cells in vitro, which could be closely related to its suppressive effects on the expression of VEGF in TE1 cells, but had no obvious effects on the cell cycle.
Introduction
E
Currently, increasing evidence has indicated that the vascular endothelial growth factor (VEGF) plays a crucial role in the recurrence and metastasis of cancers. VEGF is a glucoprotein carrying signal peptide and is highly expressed in a majority of cancers. It has been shown that VEGF binds to specific receptors (VEGFR) on endothelial cells to form the VEGF/VEGFR complex, which constitutes an angiogenic signaling pathway in cancers. This pathway may initiate a series of signal transduction and activate a complex intracellular cascade to induce angiogenesis and lymphangiogenesis, promote cancer growth, and mediate radiotherapy resistance. 11 VEGF is the most potent and specific angiogenic factor and is closely associated with the growth, metastasis, and prognosis of cancers. 12 –15
VEGF is widely expressed in numerous solid tumors in humans and has been considered to be a predictor of poor prognosis. 16 Overexpression of VEGF in tumors or serum is closely associated with local recurrence, lymph node metastasis, and prognosis. 17 –22 Multivariate analysis reveals that VEGF is an independent predictor of esophageal cancer. 23 A majority of studies have shown a negative relationship between VEGF and the radiosensitivity of cancer cells. In addition, it is suggested that the expression of VEGF in tumors is up-regulated after radiotherapy, 24,25 which is thought to be a defense mechanism against radiation. Consistently, cancers with an overexpression of VEGF are insensitive to radiotherapy, indicating a poor treatment efficacy. Therefore, the sensitivity of cancers to radiotherapy can be assessed via the expression of VEGF. Based on this assessment, cancers can be classified and personalized regimens can be developed. For cancers with an overexpression of VEGF, comprehensive treatments that inhibit VEGF expression should be used. 26 There is a great need for approaches that enhance the radiosensitivity of cancer cells by suppressing the expression of VEGF.
Thalidomide (peptide amine piperidone) was introduced as a nonprescription sedative drug in the late 1950s in Europe. In 1961, it was withdrawn due to its teratogenicity. Since the 1990s, evidence has shown that thalidomide can be used to treat complications associated with AIDS and tumors, such as multiple myeloma. These therapeutic effects were related to the suppression of angiogenesis. In 1998, thalidomide was approved in the United States by the FDA for the treatment of malignancies. 27 Subsequent studies revealed that thalidomide can suppress the expression of VEGF and the secretion of basic fibroblast growth factor 28 as well as inhibiting the synthesis of TNF-α, thus exerting immunoregulatory, antineoplastic, and antimetastatic effects. 29 The suppression of angiogenesis by thalidomide through the regulation of VEGF expression has been one of the important strategies in the treatment of solid tumors. 30 A large number of studies have been conducted to investigate the efficacy of thalidomide in combination with chemotherapy in the treatment of multiple myeloma, malignant melanoma, renal cell carcinoma, small cell lung cancer, glioma, prostate cancer, liver cancer, and other solid malignancies. 31 –35 These studies found that the addition of thalidomide resulted in superior efficacy compared with chemotherapy alone.
Based on the findings just mentioned, it was hypothesized that thalidomide could enhance the radiosensitivity and, thus, improve the radiotherapeutic efficacy of esophageal cancer cells by suppressing the expression of VEGF. In the present in vitro study, the change in radiosensitivity of esophageal cancer cells after thalidomide treatment was investigated, and the potential mechanism was also explored, to provide theoretical basis for the radiotherapy of esophageal cancer.
Materials and Methods
Reagents and instruments
Esophageal cancer cells (TE1 cells) were purchased from the Institute of Biochemistry and Cell Biology and maintained in RPMI-1640 medium (Sigma) containing 10% fetal bovine serum at 37°C in 5% CO2. Monolayer cell culture was performed, and cells were passaged every 2–3 days for a maximum of eight passages. TE1 cells in the logarithmic growth phase (<90% confluence) were harvested for use. Thalidomide (Sigma) was dissolved in dimethyl sulfoxide (DMSO; Sinopharm Chemical Reagent Co., Ltd.) at an initial concentration of 50 mg/mL followed by storage at −20°C. The thalidomide solution was diluted into designed concentrations with RPMI-1640 medium maintaining a constant concentration of DMSO in each group (≤0.4%). The present study also used TRIzol (Invitrogen), a reverse transcription–polymerase chain reaction (RT-PCR) kit (Shanghai Sangon), a Western blot kit (Beyoutime), anti-VEGF monoclonal antibody, anti-β-actin monoclonal antibody, horseradish peroxidase-conjugated goat anti-mouse IgG(H+L) (Abcam), a flow cytometer (FC-500; Beckman), a medical linear accelerator (Primus-H, Siemens; dose rate: 1 Gy/min, source-to-sample distance: 100 cm), UV spectrophotometry (Eppendorf), an electrophoresis chamber, a gel image analysis system, and a transfer device (Bio-rad).
Detection of cell survival rate by methyl thiazolyl tetrazolium assays
The cells in the logarithmic growth phase were digested with 0.25% trypsin, and a single cell suspension of 4 × 104/mL was obtained. These cells were seeded into 96-well plates (100 μL/well) and grown in RPMI-1640 medium supplemented with 10% fetal bovine serum for 24 hours. These cells were then treated with different concentrations of thalidomide (0, 25, 50, 100, 150, and 200 μg/mL) for 20, 44, or 68 hours followed by incubation with methyl thiazolyl tetrazolium (MTT; final concentration: 0.5 mg/mL) for 4 hours. The supernatant was removed, and cells were treated with 100 μL of DMSO. The absorbance (A) was determined at 570/630 nm. The survival rate was calculated as follows: survival rate = (A treated cells/A untreated cells) × 100%.
Determination of cell cycle by flow cytometry
The single-cell suspension was obtained from the cells in the logarithmic growth phase. TE1 cells were seeded into six-well plates and maintained for 24 hours to allow for adherence. Cell cycle synchronization was achieved by incubation with serum-free RPMI-1640 medium for 12 hours. The cells were then treated with thalidomide (150 μg/mL) for 24 hours. After digestion, the suspended cells were harvested and incubated with cold 70% ethanol at −20°C overnight. Thereafter, staining with propidium iodide (10 μg/mL) and RNase A (5 μg/mL) was performed in the dark for 1 hour. Cell cycle was determined using the FACSCalibur system.
Cell irradiation
Cell irradiation was performed with a medical linear accelerator. The radiation field, dose rate, and source-to-cell distance were 30 × 30 cm, 1 Gy/min, and 100 cm, respectively.
Colony-forming assay
Cells in the logarithmic growth phase were digested with trypsin, and a single-cell suspension was prepared. The cells were seeded in dishes at different densities (5 × 102, 1 × 103, 2 × 103, 5 × 103, 1 × 104, and 2 × 104 cells/mL) and maintained in the medium for 24 hours. The cells were then divided into the radiation group and the thalidomide plus radiation group. Cells in the radiation group were treated with different doses of X-rays (0, 3, 6, 9, and 12 Gy), whereas those in the thalidomide plus radiation group were treated with different doses of thalidomide (0, 50, 100, and 150 μg/mL) 30 minutes before radiation. Experiments were performed in triplicate. After radiation, the medium was immediately replaced with drug-free medium. Twelve days later, cells were fixed in methanol and then stained. The number of cell colonies containing at least 50 cells was counted under a microscope, and the colony-forming efficiency was determined. The survival rate of cells in the thalidomide plus radiation group was normalized by the survival rate of cells in the radiation group. Analysis was performed with Graphpad prism 5 Demo software, and a multitarget click model [S = 1 − (1 − e−D/D0)N ] was applied to delineate the survival curve. Radiobiological parameters including D 0 (mean lethal dose), D q (quasi-threshold dose), and SF2 (survival fraction under X ray of 2 Gy) were then calculated.
Detection of VEGF mRNA expression by RT-PCR
The primers for VEGF and β-actin (GenBank) were designed. For VEGF, forward: 5′-AGAAAGACAGATCACAGG-3′, reverse: 5′-CGGTACAAATAAGAGAGC-3′. For β-actin, forward: 5′-CTACAATGAGCTGCGTGTGG-3′, reverse: 5′-AAGGAAGGCTGGAAGAGTGC-3′. The anticipated sizes of the amplified products were 660 bp (VEGF) and 518 bp (β-actin).
Cells in the logarithmic growth phase were digested with trypsin and seeded into dishes. Cells were treated with different concentrations of thalidomide (50, 100, and 150 μg/mL) after cell adherence. Half an hour later, radiation (12 Gy) was performed, and the total RNA was extracted 24 hours later. The optical density was detected at 260 nm with an ultraviolet spectrophotometer, and the RNA concentration was calculated.
Synthesis of cDNA was performed as follows: 6 μL of the PCR mixture, including 300 ng of RNA, 0.5 μL of Random primers, and RNase-free ddH2O were used. The reaction was initiated at 70°C for 10 minutes, and the mixture was allowed to stay on ice. Then, 2 μL of 5 × M-MLV buffer, 2 μL of 2.5 mM dNTP, and 0.5 μL of 200 U/μL M-MLV Rtase were added in sequence, and the reaction was continued at 30°C for 10 minutes, 42°C for 1 hours, and 70°C for 15 minutes. RT-PCR: The synthesized cDNA was used as a template, and VEGF and β-actin were amplified with specific primers. The 25 μL of reaction mixture consisted of 0.125 mL of 5 U/μL EX Taq HS, 2.5 μL of 10 × EX Taq buffer, 1 μL of 2.5 mM dNTP mixture, 0.5 μL of each primer (25 μM) for VEGF or β-actin, 1 μL of cDNA, and RNase-free ddH2O.
For VEGF, the cycling conditions for PCR were 30 cycles of denaturation (94°C for 30 seconds), annealing (59°C for 30 seconds), and extension (72°C for 20 seconds). A preheating step at 94°C for 5 minutes and a final extension step of 7 minutes at 72°C were also carried out. For β-actin, the cycling conditions for PCR were 30 cycles of denaturation (94°C for 30 seconds), annealing (69°C for 30 seconds), and extension (72°C for 20 seconds). A preheating step at 94°C for 5 minutes and a final extension step of 7 minutes at 72°C were also carried out. The amplified products were separated by electrophoresis on a 1% agarose gel followed by staining with 0.5 μg/mL ethidium bromide. The bands were observed with a gel analyzer. The images were captured, and semi-quantitative analysis was carried out using Bandscan5.0 gel image analysis software.
Detection of VEGF protein expression by Western blot
Cells in the logarithmic growth phase were digested with trypsin and maintained in dishes. After adherence, cells were treated with 150 μg/mL thalidomide, and radiation with 12 Gy was performed 30 minutes later. Total protein was extracted 24 hours later, and the protein concentration was determined by the BCA method. The extracted protein was stored at −80°C for use. The protein samples were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred onto a nitrocellulose membrane, and then blocked with blocking buffer overnight. Then, the membrane was treated with primary antibodies against β-actin and VEGF (1
Statistical analysis
Statistical analysis was performed using SPSS 13.0 statistic analysis software. Data are presented as the mean ± standard deviation (
Results
MTT assay
As shown in Figure 1, when compared with the control group, no significant difference in the survival rate was observed after treatment with the different concentrations of thalidomide for 24 hours (p > 0.05). However, after treatment with 150 or 200 μg/mL of thalidomide for 48 or 72 hours, the survival rate was significantly decreased when compared with the control group (*p and **p < 0.01).
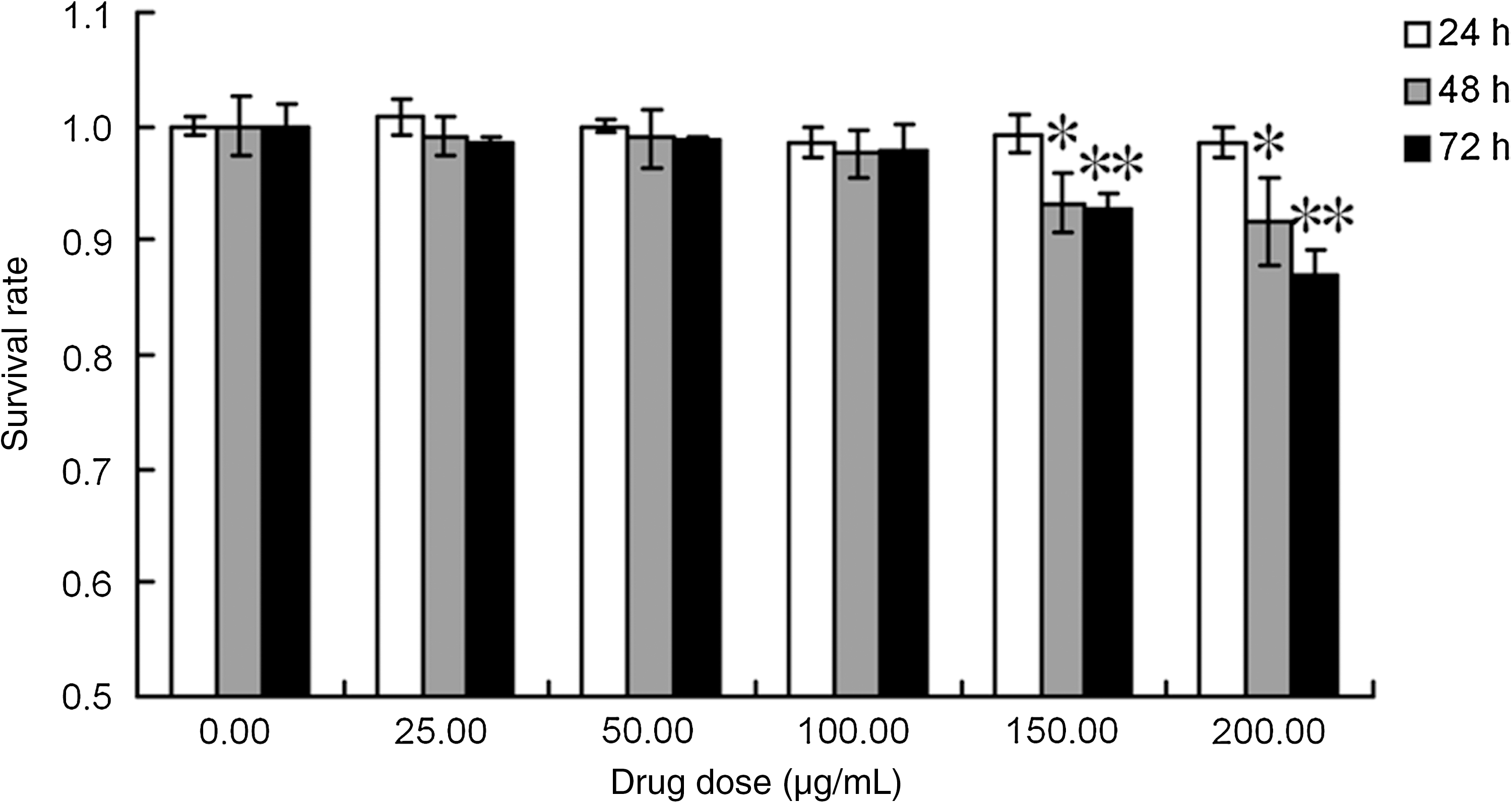
Effects of thalidomide on the survival rate of TE1 cells. Survival rate of TE1 cells was determined by methyl thiazolyl tetrazolium assays after continuous exposure to thalidomide for 24, 48, or 72 hours. As the drug concentration increased, the growth inhibition ratio of TE1 cells increased. After treatment with 150 or 200 μg/mL of thalidomide for 48 or 72 hours, the survival rate was significantly decreased when compared with the control group (*p and **p < 0.01).The image represents results from three independent experiments.
Effects of thalidomide on the cell cycle of TE1 cells
As shown in Table 1 and Figure 2, no significant difference in the proportion of cells in the G1, G2 and S phase was observed between 0 and 150 μg/mL thalidomide group (p > 0.05).
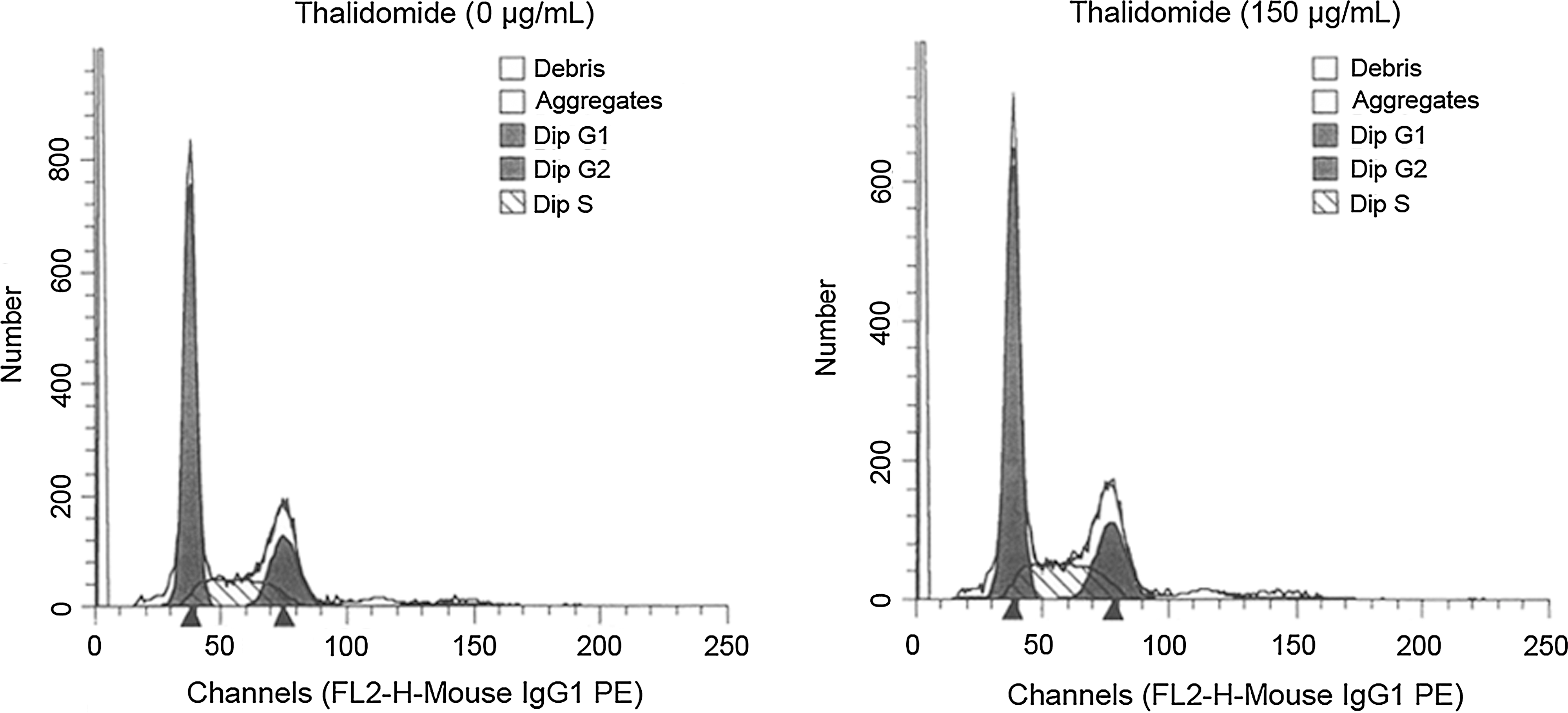
Effects of thalidomide on the cell cycle of TE1 cells. The cell cycle distribution of TE1 cells was determined by flow cytometry. Cell cycle synchronization was achieved by incubation with serum-free RPMI-1640 medium for 12 hours. The cells were then treated with thalidomide (150 μg/mL) for 24 hours.
The cell cycle distribution of TE1 cells at 24 hours after treatment with 0 and 150 μg/mL thalidomide were compared. No significant difference in the proportion of cells in the G1, G2, and S phase was observed between 0 and 150 μg/mL thalidomide group. Results are shown as the mean ± SD of three independent experiments conducted in triplicate.
Effects of thalidomide on the radiosensitivity of TE1 cells
Figure 3 shows the survival curves of cells after treatment with thalidomide and different doses of radiation. The survival rate of TE1 cells was decreased by the dose increase of irradiation. The dose-response curves of TE1 cells were characterized by the shoulder area at a low dose and linear area at a high dose, which represented the multitarget click model.
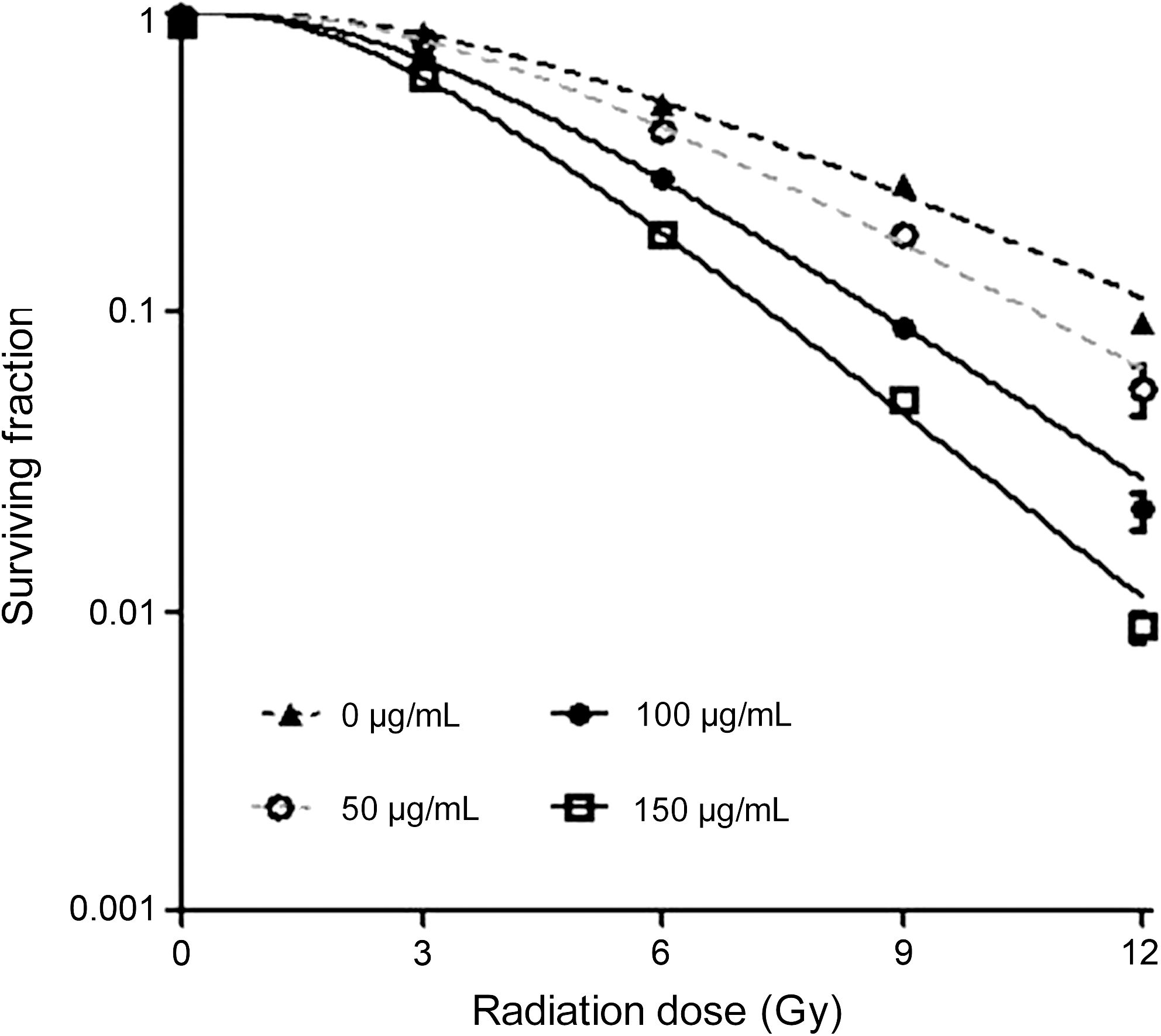
Effect of thalidomide plus radiation on colony-forming efficiency. Synergistic effects of combined thalidomide and radiation on the colony forming of TE1 cells were observed. After treating by thalidomide (0, 50, 100, 150 μg/mL) and radiation (0, 3, 6, 9, 12 Gy), the survival rate of TE1 cells was decreased by the dose increase of irradiation.
Table 2 shows the values of several radiobiological parameters determined by the multitarget click model. D 0 is the mean lethal dose at which the survival rate was decreased by 63%. The lower the D 0, the higher the radiosensitivity of the TE1 cells. D q, the quasi-threshold dose, represents the capability of TE1 cells to accumulate sublethal damage. This is illustrated by the shoulder area of the dose-response curve, in which cells counteract the toxicity of thalidomide. A lower D q purports a narrower shoulder area, indicating the enhanced radiosensitivity. SF2 represents the survival rate under an X-ray of 2 Gy and is an important indicator of radiosensitivity. A greater SF2 purports more resistance to radiation in the TE1 cells. The sensitizing enhancement ratio (SER) is calculated as follows: SERD0(SERDq) = [D 0(D q)]control/[D 0(D q)]treatment. As shown in Table 2, the values of D 0, D q and SF2 decreased gradually with the increase in the thalidomide concentrations, indicating a dose-dependent effect. When the irradiation dose was D 0 or D q, the SER of TE1 cells was 1.38 or 1.44, respectively, after treatment with 100 μg/mL thalidomide. The SER increased to 1.47 or 1.78, respectively, after treatment with 150 μg/mL thalidomide.
Biological parameters of radiation determined by multitarget click model: D 0, mean lethal dose; D q, quasi-threshold dose; SF2, survival fraction under X-ray of 2 Gy. The values of D 0, D q, and SF2 were significantly decreased when compared with the 0 μg/mL thalidomide group (* p < 0.01). Sensitizing enhancement ratio SERD0(SERDq) = [D 0(D q)]control/[D 0(D q)]treatment. When the irradiation dose was D 0 or D q, the SER of TE1 cells was 1.38 or 1.44, respectively, after treatment with 100 μg/mL thalidomide. The SER increased to 1.47 or 1.78, respectively, after treatment with 150 μg/mL thalidomide.
SER, sensitizing enhancement ratio.
Effects of thalidomide plus X-ray on the mRNA expression of VEGF
As shown in Figures 4 and 5, the band of β-actin was found at 518 bp and had an even density, whereas the band of VEGF mRNA was observed at 660 bp with an uneven density. The expression of VEGF mRNA decreased with the increase in the thalidomide concentrations. After treatment with 100 or 150 μg/mL thalidomide, the expression of VEGF mRNA was significantly decreased compared with the control group (*p < 0.01). After radiation, the VEGF mRNA expression was not significantly different than the control group (p > 0.05). When compared with the control group, the mRNA expression was significantly decreased after treatment with thalidomide alone or thalidomide plus radiation (*p < 0.01). When compared with the thalidomide group, the VEGF mRNA expression in the thalidomide plus radiation group was significantly decreased (**p < 0.01).
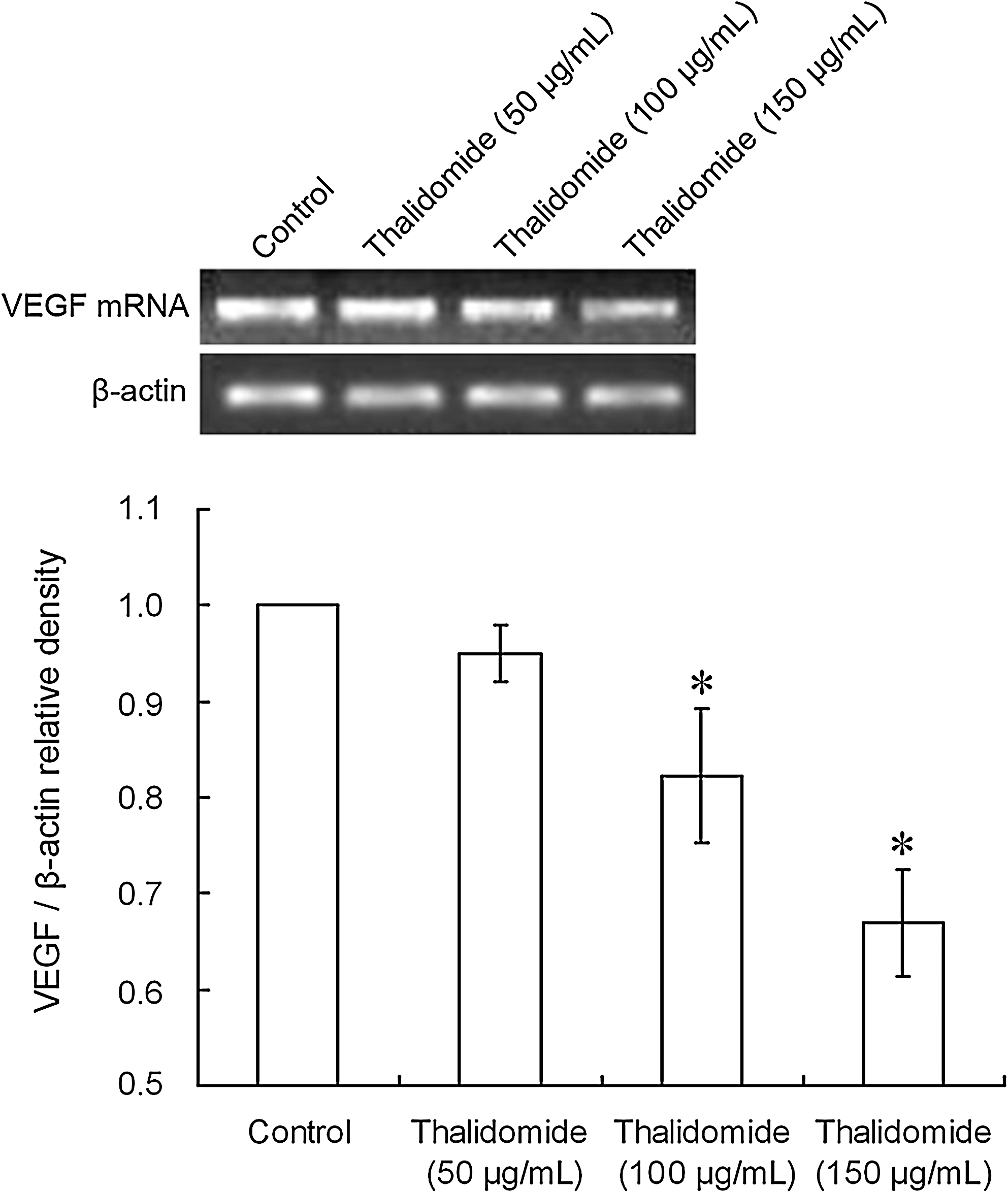
Effect of thalidomide of different concentrations on the mRNA expression of vascular endothelial growth factor (VEGF): (control = 0 μg/mL; 50 μg/mL thalidomide; 100 μg/mL thalidomide; 150 μg/mL thalidomide) reverse transcription–polymerase chain reaction assay indicated the ability of thalidomide to downregulate the VEGF mRNA expression. After treatment with 100 or 150 μg/mL thalidomide, the expression of VEGF mRNA was significantly decreased compared with the control group (*p < 0.01). Band intensity was analyzed by densitometry using the program Bandscan5.0.
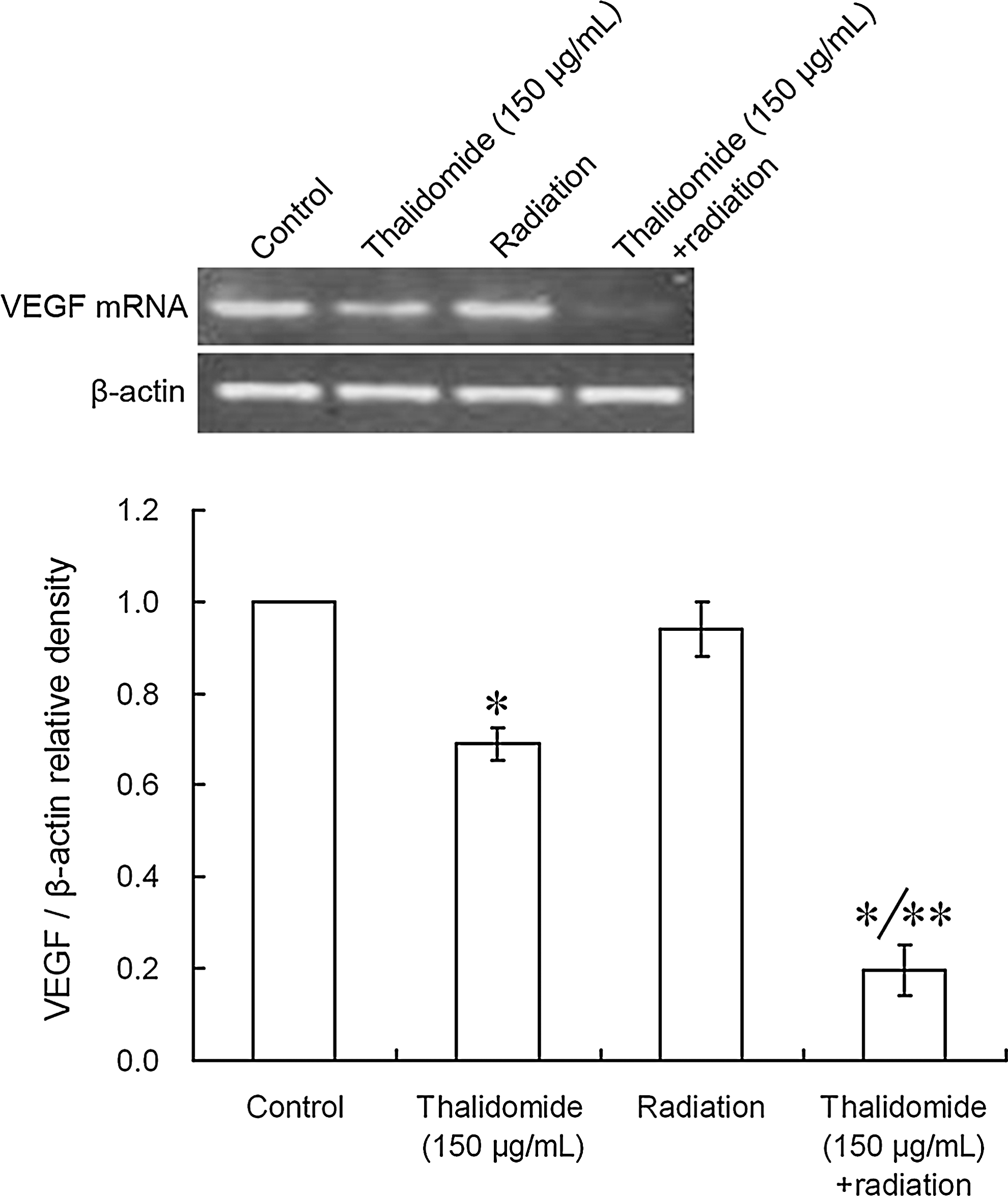
Effect of thalidomide plus X-ray radiation on the mRNA expression of VEGF: (control = 0 μg/mL; 150 μg/mL thalidomide; radiation; 150 μg/mL thalidomide + radiation). When compared with the control group, the VEGF mRNA expression was significantly decreased after treatment with thalidomide alone or thalidomide plus radiation (*p < 0.01). When compared with the thalidomide group, the VEGF mRNA expression in the thalidomide plus radiation group was significantly decreased (**p < 0.01). Band intensity was analyzed by densitometry using the program Bandscan5.0.
Effects of thalidomide plus X-ray radiation on VEGF protein expression
As shown in Figure 6, no significant difference in the expression of VEGF was observed between the radiation group and the control group (p > 0.05). After treatment with thalidomide alone or thalidomide plus radiation, the VEGF expression was significantly decreased (*p < 0.01) when compared with the control group. Further, when compared with the thalidomide group, the VEGF expression in the thalidomide plus radiation group was significantly decreased (**p < 0.01).
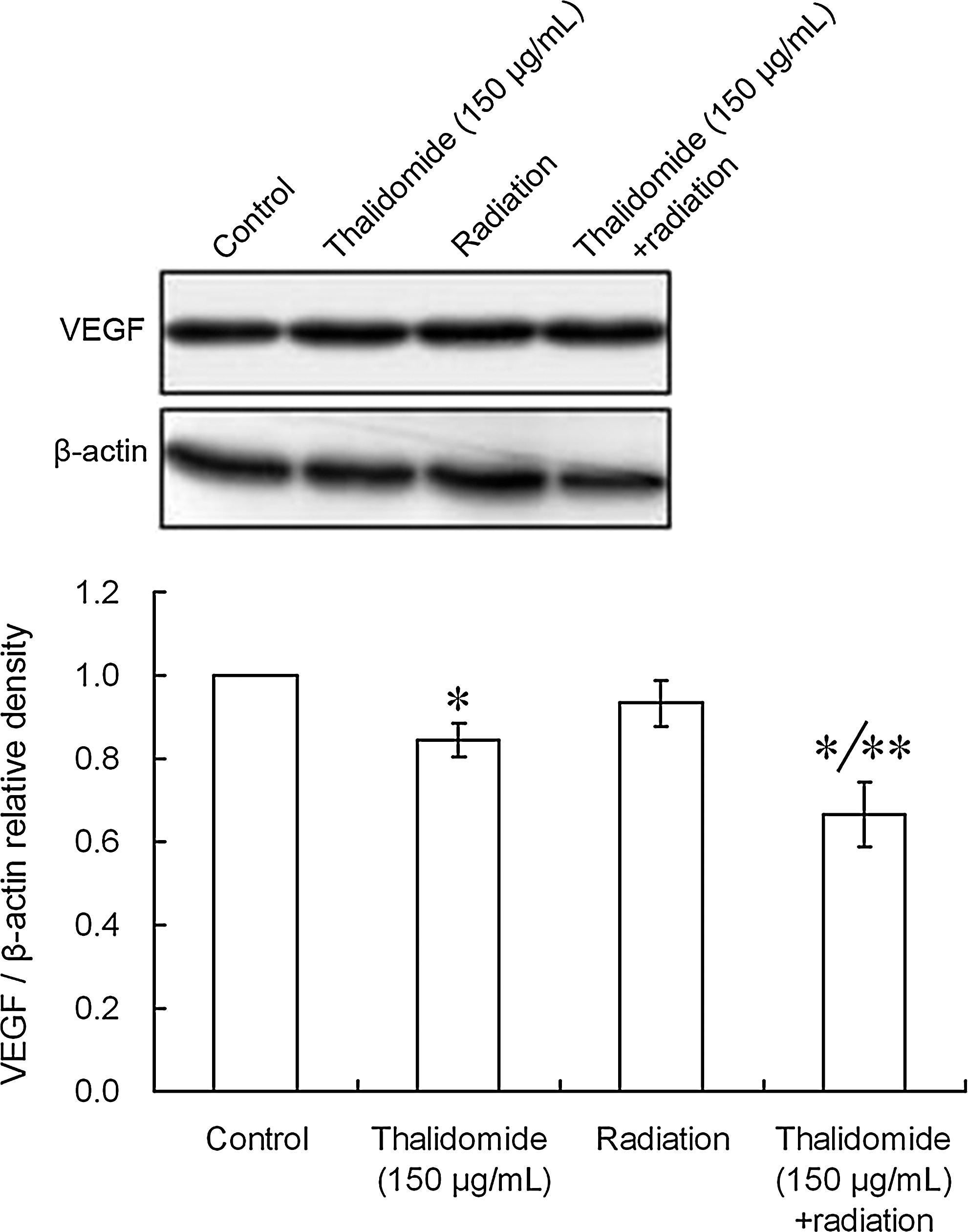
VEGF expression after treatment with thalidomide plus X-ray radiation. (1, control = 0 μg/mL; 2, 150 μg/mL thalidomide; 3 radiation; 4, 150 μg/mL thalidomide + radiation). When compared with the control group, the VEGF expression was significantly decreased after treatment with thalidomide alone or thalidomide plus radiation (*p < 0.01). Further, when compared with the thalidomide group, the VEGF expression in the thalidomide plus radiation group was significantly decreased (**p < 0.01). Band intensity was analyzed by densitometry using the program Bandscan5.0.
Discussion
Angiogenesis inhibitors in combination with chemotherapeutics have been used as a routine regimen in the treatment of cancers. The mechanisms underlying the therapeutic effects of these inhibitors plus radiotherapy and/or chemotherapy have been investigated. Currently, more than 20 angiogenesis inhibitors have been applied in pre-clinical studies. Kozin et al. 36 studied the efficacy of the anti-VEGFR2 antibody plus 5 dose fractions of local radiation in mice bearing two different human tumor xenografts. Their results showed that the favorable local control of cancers could be achieved at 59% and 76% of the usual radiation dose when the radiation was performed in combination with treatment with the VEGFR2 antibody. These results indicated that treatment against VEGF could suppress the clonal growth of cancer cells and also confirmed the hypothesis that the suppression of VEGF could enhance the effectiveness of radiotherapy. In an animal model, Zips et al. 36 –39 compared the efficacy of a tyrosine kinase inhibitor of VEGFR plus radiation in the treatment of cancers, and they found that the suppression of VEGF expression could enhance the radiosensitivity of cancer cells. Greco et al. 40 found that, after hypoxia or radiation, the VEGF expression in bladder T24 cells and breast cancer MC cells was markedly increased. In addition, the radiosensitivity of these cell lines was profoundly increased after treatment with antisense gene therapy targeting VEGF.
Gorski et al. 41 indicated that a VEGF neutralizing antibody in combination with radiotherapy could enhance the local control rate better than radiotherapy alone or treatment with VEGF neutralizing antibody alone. Ansiaux et al. 42 investigated the efficacy of thalidomide in the treatment of FSAII tumor. The results indicated that thalidomide could induce tumor re-oxygenation within 2 days after antiangiogenetic treatment, which was correlated with a reduction in interstitial fluid pressure and an increase in perfusion, suggesting thalidomide radiosensitization. Epperly et al. 43 explored the effects of thalidomide on squamous cell carcinoma and multiple myeloma in an animal model, and they found that thalidomide could radiosensitize normal hematopoietic cells but not cancer cells. They postulated that the radiosensitization of thalidomide was selective. To date, no study has reported the efficacy of thalidomide alone or in combination with radiation on esophageal cancer cells.
In the present study, the proliferation of TE1 cells was detected by MTT assay after thalidomide treatment, and the results show that thalidomide was able to directly suppress the proliferation of TE1 cells in a concentration- and time-dependent manner. The proliferation of TE1 cells was markedly inhibited after treatment with 150 or 200 μg/mL thalidomide for 48 or 72 hours (both p < 0.05). Further, the inhibition rate of TE1 cell proliferation was as high as 87% after treatment with 200 μg/mL thalidomide for 72 hours. In addition, Marriott et al. 44 found that thalidomide could suppress the proliferation of colon cancer cells (SW620 cells), pancreatic cancer (T3M-4 cells), and prostate cancer cells (PC-3 cells) and ultimately resulted in the apoptosis of cancer cells. The suppressive effect was related to a down-regulation of bcl-2 expression and the activation of caspase-3. Ng et al. 45 found that thalidomide could suppress the proliferation of human umbilical vein endothelial cells, PC-3 cells, and human prostate cancer cells (LNCaP cells). This suppressive effect was related to the suppression of TNF and basic fibroblast growth factor and decreased the stability of COX-2 mRNA. Hideshima et al. 46 reported that thalidomide could induce apoptosis and G1 arrest in myeloma cells, thus exerting antineoplastic effects. In the present study, the cell cycle of TE1 cells after treatment with thalidomide was determined by flow cytometry, and the results indicate that thalidomide did not profoundly affect the cell cycle of TE1 cells, which is inconsistent with the work of Hideshima et al. This could be explained by different biological characteristics. The doubling time of TE1 cells is longer than that of myeloma cells. In addition, the length of the thalidomide treatments in the two studies were different and could be another cause of the conflicting results. The antineoplastic effects of thalidomide are controversial, and to date, the exact mechanism of these effects is poorly understood. These can be explained by different types of cancer cells, different sites at which cancer occurs, and different ways in which drugs are administered. Therefore, further studies to confirm the antineoplastic effects of thalidomide and clarify the potential mechanisms are needed.
In the present study, a colony-forming assay and multitarget click model were employed to delineate the survival curve and investigate the effect of thalidomide on the radiosensitivity of TE1 cells in vitro. Few studies have explored the effect of thalidomide on the radiosensitivity of cancer cells. Our results show that the survival rate of TE1 cells decreased exponentially with the increase of the irradiation dose. The dose-survival curve was a typical multitarget-click model having a shoulder area at low doses, which suggests the TE1 cells have a resistance to sublethal damage and radiation resistance. The D 0, D q, and SF2 decreased gradually with the increase of the thalidomide concentration, accompanied by an increased slope of the survival curve, suggesting a concentration-dependent effect. These results implied that the different concentrations of thalidomide were able to enhance the radiosensitivity of TE1 cells. When the concentration of thalidomide was 150 μg/mL, the shoulder area was profoundly narrowed, and the cells lost their resistance to sublethal damage. When the radiation dose was D 0 and D q, the SER was 1.44 and 1.78, respectively. These results suggest that thalidomide could significantly enhance the radiosensitivity of TE1 cells.
To explore the potential mechanism underlying the radiosensitization of thalidomide, RT-PCR and Western blot were used to detect the expression of VEGF mRNA and protein after treatment with thalidomide plus radiation. Our results show that the mRNA expression of VEGF decreased with the increase of the thalidomide concentrations. When compared with the control group, the mRNA expression of VEGF was markedly decreased after treatment with 100 or 150 μg/mL thalidomide alone (p < 0.01). However, the mRNA and protein expressions of VEGF were not dramatically changed by radiation alone (p > 0.05). When compared with thalidomide alone, treatment with thalidomide plus radiation decreased the mRNA and protein expressions of VEGF in a concentration-dependent manner (p < 0.01). Huang et al. 47 found that VEGF was implicated in the repair of sublethal damage and potential lethal damage. Our results imply that thalidomide can compromise the repair of sublethal damage and potentially lethal damage through the down-regulation of VEGF expression and subsequently enhance the radiosensitivity of esophageal cancer cells. As a result, more cancer cells were killed by radiation, and the secretion of VEGF was further decreased. Therefore, synergetic antineoplastic effects were observed during treatment with thalidomide plus radiation.
Presently, numerous studies have shown that the suppression of VEGF or VEGFR expression in combination with radiotherapy is promising in enhancing the efficacy of radiotherapy for cancers. However, the results of these studies were influenced by a variety of factors, including the VEGF pathway. The specific mechanism underlying the enhanced radiosensitivity by thalidomide is still unknown, and further studies are required to clarify whether other pathways are involved in the radiosensitization by thalidomide.
Taken together, our results demonstrate that thalidomide can suppress the growth of TE1 cells and VEGF secretion in vitro. In combination with X-ray radiation, thalidomide enhanced the radiosensitivity of TE1 cells, which was accompanied by an increased suppression of VEGF secretion. Our results may provide a theoretical basis for the experimental and clinical use of thalidomide in conjunction with X-ray radiation. However, the present evidence is not enough to support the clinical application of thalidomide as a radiosensitizing drug, and further studies are required.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 30870585), the Training Program of Young Scientists of Changzhou (CQ2008006), and the Guiding Project for Social Development of Changzhou (CS2008920).
Disclosure Statement
The authors declare that they have no competing interests.