
Abstract
Purpose:
Bcl2/adenovirus EIB 19 kDa-interacting protein 3 (BNIP3) is a proapoptotic member of the Bcl-2 family. To address its potential as a therapeutic target for radiosensitization, this study investigated the effect of Bnip3 expression on radiosensitivity of cervical cancer in vitro and in vivo.
Materials and Methods:
In vitro: A plasmid expressing the BNIP3 gene was transfected into human cervical cancer HeLa cells using Lipofectamine2000, and western blot and immunohistochemistry analysis were performed to evaluate the expression of BNIP3 in transfected cells. The effects on radiation-induced apoptosis were investigated using a clone formation assay and flow cytometry. In vivo: A total of 6 × 106 HeLa cells were subcutaneously inoculated into the dorsal flank of nude mice, and plasmids expressing the BNIP3 gene were injected into the mice via the tail vein. Tumor volume was calculated, and immunohistochemistry was used to detect the expression of BNIP3 in tumor cells. TUNEL assays were performed to determine the apoptosis rates in tumor tissues.
Results:
Transfection with the recombinant BNIP3 plasmid increased expression of the Bnip3 protein in tumor cells. This apoptosis regulator significantly decreased the viability of cells (p < 0.01) and increased the apoptosis rates (p < 0.01) both in vitro and in vivo. The antitumor effect of radiotherapy was enhanced by overexpression of BNIP3, as revealed by tumor growth curve analysis.
Conclusions:
Radiosensitization in human cervical cancer cells was observed after treatment with the recombinant BNIP3 plasmid in vitro and in vivo. Results suggested that BNIP3 may play a role in enhancement of radiotherapy efficiency, and its expression may have a synergistic effect on radiation treatments.
Introduction
Currently, cervical cancer is a major risk to a women's health and life. Radiotherapy is the primary approach for the treatment of cervical cancer; however, clinical observations indicate that radiotherapy often confronts many problems, including poor efficacy and cancer radioresistance, and these problems seriously influence the outcome of cervical cancer treatments. Therefore, identification of genes that influence radiosensitivity is of significant interest. If a gene has a dose-dependent or temporal effect on radiosensitivity, 1,2 is able to influence cell apoptosis at certain radiation doses, or causes changes to the cell cycle that influence radiosensitivity, then this gene is a potential therapeutic target for increasing the efficacy of radiotherapy, similar to the p53, p16, and Fas genes. 3 Moon et al. transfected the wild-type p53 gene into tumor cells and confirmed that its expression has obvious effects on radiosensitization in small cell lung cancer and head and neck squamous cell carcinoma. These results are consistent with other studies. 4 –8 The research presented by Rhee et al. 9 also demonstrated that p16 has a similar effect on radiosensitization in head and neck cancer.
Bcl-2/adenovirus E1B 19 kDa-interacting protein 3 (BNIP3) is a mitochondrial protein and a proapoptotic member of the Bcl-2 family. 10 BNIP3 expression is reduced in most tumors, including most pancreatic cancers, hematopoietic tumors, and gastric cancers. 11 –13 In a study of cervical cancer, Leo et al. 14 performed immunohistochemistry on tissues from 50 cervical cancers, and BNIP3 protein expression was observed in 82% of the investigated samples. However, only 20% of all samples demonstrated strong expression, whereas weak expression of BNIP3 was found in 32%, moderate staining in 30%, and the remaining 18% of the samples were BNIP3 negative. There are a variety of possible reasons for this variability in BNIP3 expression. The most important factor is methylation of the BNIP3 promoter. 11,12,15 Further, tumor chemoresistance correlates with abnormal expression of BNIP3. Murai et al. 12 found that expression of BNIP3 was decreased in colon cancer cell lines that were chemoresistant to 5-fluorouracil. Tang et al. 16 observed that colon cancer cell lines resistant to oxaliplatin expressed low levels of BNIP3, and this cell line was also resistant to 5-fluorouracil. Other studies have made similar conclusions, indicating that BNIP3 expression correlates with chemoresistance. 17 –19 Whether the absence of BNIP3 expression is correlated with tumor radioresistance is unknown. Therefore, the present study investigated whether BNIP3 overexpression in cervical cancer could improve radiation sensitivity.
Materials and Methods
Cell culture
The human cervical cancer HeLa cell line was purchased from the School of Life Sciences, Sichuan University, Sichuan Province, China. HeLa cells were cultured in RPMI-1640 culture medium containing 10% calf serum in an atmosphere of 5% carbon dioxide (CO2) at 37°C.
Transient transfection
Plasmids pDsRed-N1 and pDsRed-BNIP3 were acquired from Doctor Chen Ni (Department of Pathology, West China Hospital, Sichuan University, Sichuan Province, China). The plasmids were sequenced by Invitrogen (Carlsbad, CA). Plasmid DNA from Escherichia coli cell lysates was extracted and purified using a PureLink™ Hipure Plasmid DNA Purification Kit (Invitrogen). HeLa cells were transfected with pDsRed-N1 or pDsRed-BNIP3 using Lipofectamine2000 (Invitrogen). In brief, cells were trypsinized and plated onto six-well plates. Then, transfection reagent was added and incubated at room temperature for 5 minutes. The appropriate volume of plasmid DNA was then added and the cells were incubated for an additional 20 minutes. Fluorescein-labeled pDsRed-N1- or pDsRed-BNIP3-transfected cells were examined using a fluorescence microscope to determine transfection efficiency after 24 hours. The cells were then prepared for western blot analysis, clone formation assays, or flow cytometry.
Western blot analysis
Total protein was extracted using lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 0.1% SDS, 1 mM phenylmethylsulphonyl fluoride, 0.5 mM EDTA, 0.61 mM leupeptin, and 0.1% pepstatin). Equal amounts of protein were separated via SDS-PAGE and then electrophoretically transferred onto nitrocellulose membranes. The membranes were blocked with 5% nonfat milk in TBST and incubated with BNIP3 (Sigma-Aldrich, St. Louis, MO) and β-actin antibodies (Sigma-Aldrich) overnight at 4°C. After the membranes were exposed to the respective secondary antibody for 2 hours, the blots were analyzed by chemiluminescence detection and autoradiography.
Groups
Six groups were analyzed in this study: control (NS), pDsRed-N1 (null), pDsRed-BNIP3, radiation, pDsRed-N1 + radiation (null + radiation), pDsRed-BNIP3 + radiation.
In vitro irradiation
Cells were transiently transfected with plasmid DNA (pDsRed-N1 or pDsRed-BNIP3) for 24 hours before irradiation and then trypsinized and made into single-cell suspensions in fresh medium for clone formation assays. Alternatively, after irradiation, the cells were incubated at 37°C for 24 hours before flow cytometry analysis. The cells were irradiated at room temperature with single X-ray doses generated by a Siemens linear accelerator operating at 6 MV at a rate of 400 cGy/minute and an irradiation field of 14 × 10 cm.
Clone formation assay
Immediately after irradiation, appropriate numbers of cells were seeded into plastic culture dishes (6 cm in diameter) and incubated at 37°C with 5% CO2 for about 2 weeks. The colonies were fixed with ethanol and stained with 2% Giemsa. Colonies composed of 50 or more cells were scored, and plating efficiencies and survival rates were calculated. 20 The cells were exposed to doses of 0, 2, 4, or 6 Gy. The clone formation rate was calculated by the following equation: PE = colonies/cells seeded × 100%. This experiment was performed in triplicate and repeated at least two times.
Flow cytometry
HeLa cells were transiently transfected for 24 hours and then irradiated with X-rays at 6 MV and a rate of 400 cGy/minute; total doses were 2 Gy. The cells were harvested 24 hours after irradiation and apoptosis was analyzed via flow cytometry using an Annexin–fluorescein isothiocyanate (FITC)/propidium iodide (PI) apoptosis detection kit (KeyGEN Biotechnology, Nanking, Jiangsu, China) following the manufacturer's instructions. In brief, the cells were harvested with trypsin–EDTA, pelleted by centrifugation, washed twice with 1 × phosphate-buffered saline (PBS), and resuspended in Annexin V binding buffer. FITC-conjugated Annexin V and PI were added to the cells, incubated for 30 minutes at room temperature, protected from light, and then analyzed via flow cytometry (ESP Elite; Beckman Coulter, Fullerton, CA). Annexin V-positive cells were considered in the early stages of apoptosis, whereas the cells in the late stages of apoptosis were both Annexin V and PI positive.
Tumor model and treatment
Human cervical cancer xenografts were established in 5–6-week-old female nude mice (Balb/c nu/nu) through subcutaneous inoculation of 6 × 106 HeLa cells into the dorsal flank. Tumor cell suspensions were prepared from HeLa cells grown as monolayers in vitro. The animals used in this study were obtained from the Animal Center, Sichuan University, Sichuan Province, China, and maintained under specific pathogen-free conditions. Tumors were measured every 3 days, and the tumor volume was calculated using the following equation: A × B 2 × 0.5, where A and B are the largest and smallest diameters. 21 Each tumor reached a diameter of ∼5–6 mm at 10 days after inoculation. The nude mice were then randomly divided into 6 groups (5 mice per group) and plasmid therapy was started via injection of DNA/liposome complexes into the tail vein at 5 μg/dose every 2 days for a total of 10 times. The DNA/liposome complexes were formed by adding liposome solution to the DNA solution, at a ratio of 3 μg of liposome to 1 μg of DNA, then diluted with 5% glucose solution to 100 μl, gently mixed, and incubated for 30 minutes at room temperature. The tumors were irradiated at 3 Gy/day per tumor for 4 days after the seventh plasmid treatment.
Immunohistochemistry
The tumor tissues removed from the animals used in this study were formalin fixed, paraffin embedded, and cut into 4-μm-thick sections for immunohistochemical analysis. In brief, sections were deparaffinized and rehydrated as follows: 3 × 3 minutes with xylene, 3 × 2 minutes with 100% ethanol, 2 minutes with 95% ethanol, 2 minutes with 80% ethanol, 2 minutes with 70% ethanol, and 5 minutes with PBS (NaCl 137 mM, KCl 2.7 mM, Na2HPO4 4.3 mM, and KH2PO4 1.4 mM [pH 7.2–7.4]). Antigen retrieval was performed by heating sections in Tris–ethylenediaminetetraacetic acid (pH 9) for 15 minutes in a microwave oven. Endogenous peroxidases were blocked by soaking slides in a solution of 3% H2O2 for 10 minutes at room temperature and then washed 3 × 5 minutes with PBS. Blocking buffer was immediately added to each section and then incubated for 1 hour at room temperature. A total of 50 μL of Bnip3 antibody (Sigma-Aldrich) in blocking buffer (1:250) was added per section and incubated for 1 hour at room temperature in a humidified chamber. Slides were then washed 3 × 5 minutes with PBS. A total of 50 μL of secondary antibody was added per section and incubated for 1 hour at room temperature. The secondary antibody was removed, and the slides were washed 3 × 5 minutes in PBS + 0.1% Tween-20. DAB solution was made in 5 mL ddH2O with the addition of two drops of buffer, two drops of H2O2, and four drops of DAB and then immediately added to the slides. After the color change, slides were drained and place into ddH2O for 5 minutes. DAB was removed with bleach, and the slides were counterstained with hematoxylin, washed 10 minutes with ddH2O, and immediately dehydrated in 70%, 80%, and 100% ethanol. The slides were mounted with Permount and sealed with a coverslip and nail polish.
TUNEL assay
Cells undergoing apoptosis in the tumor tissues were identified via TUNEL assay (DeadEnd™Fluorometric TUNEL System; Promega, Madison, WI) following the manufacturer's instructions. In brief, this TUNEL assay detects fragmented DNA of apoptotic cells by incorporating fluorescein-12-dUTP at the 3'-OH ends of the DNA using the enzyme terminal deoxynucleotidyl transferase. Apoptotic cells were counted under a fluorescence microscope from at least five randomly selected fields. A total of 1000 tumor cells were counted per slide. The apoptotic index was defined as follows: Apoptotic index (%) = 100 × apoptotic cells/total tumor cells.
Statistical analysis
Results are reported as means ± SD. To determine the statistical significance of differences, an independent sample t-test was performed using SPSS 16.0 software. A value of p < 0.05 was considered statistically significant.
Results
Recombinant BNIP3 expression in vitro
The pDsRed-N1 and pDsRed-BNIP3 plasmids were each transfected into HeLa cells, with transfected rates of about 80%–85% and 78%–82%, respectively (Fig. 1). Recombinant plasmid expression was confirmed by western blot analysis. Western blot analysis for BNIP3 revealed that cells transfected with pDsRed-BNIP3 expressed increased levels of the 60 kDa form of the Bnip3 protein compared with cells transfected with pDsRed-N1 and untransfected controls after a 24-hour transfection. No change in the 30 kDa form of the Bnip3 protein was visible (Fig. 2).

Fluorescein-labeled pDsRed-N1- or pDsRed-BNIP3-transfected cells were examined using a fluorescence microscope to determine transfection efficiency after 24 hours.
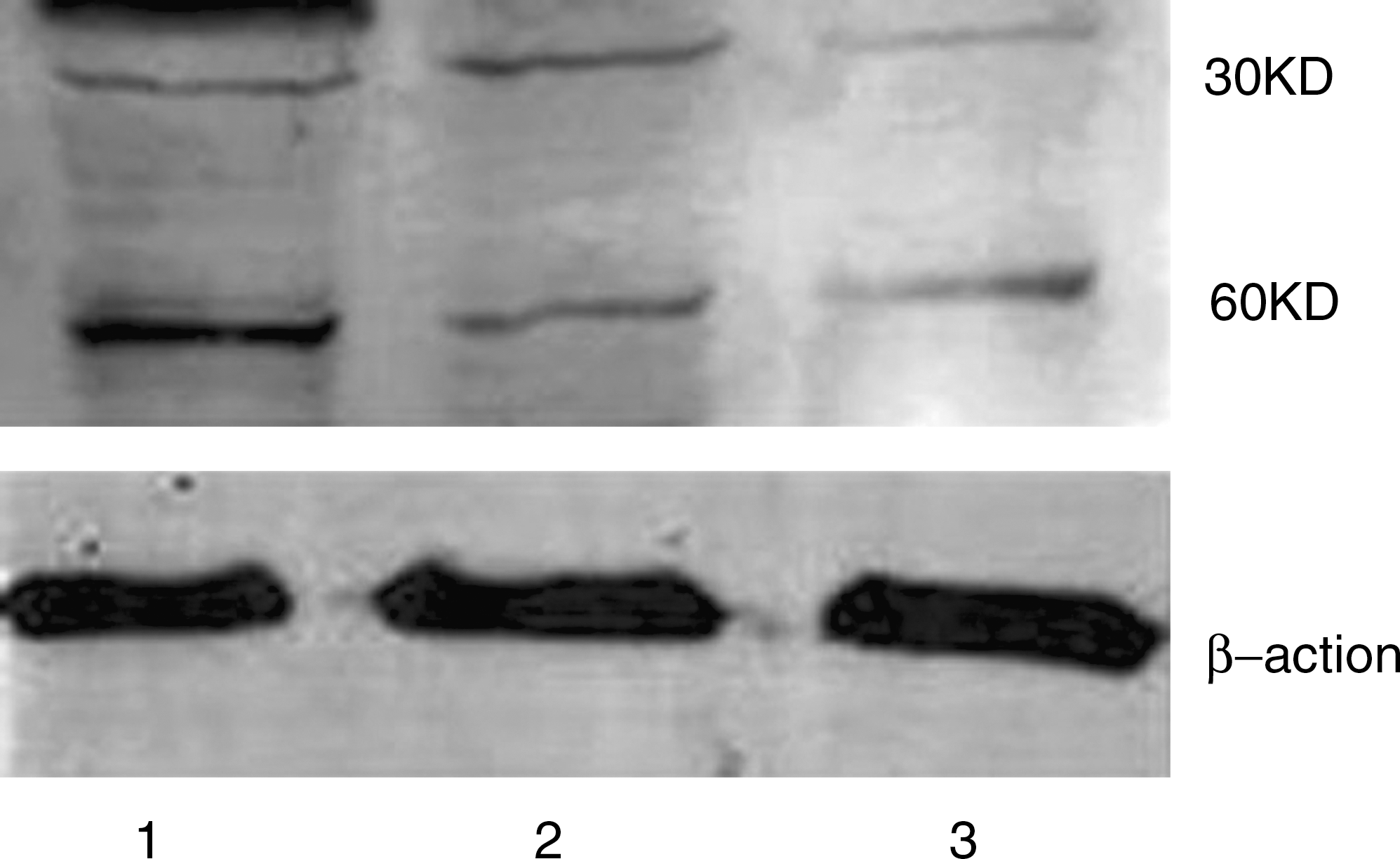
Western blot analysis showing the expression of Bnip3 protein after transfection with recombinant plasmid at 24 hours in HeLa cells. Recombinant human Bnip3 protein was expressed as two bands of ∼30 and 60 KD in pDsRed-BNIP3-transfected HeLa cells (lane 1). The 60 kDa form of Bnip3 demonstrated a visible increase relative to the empty plasmid (pDsRed-N1)-transfected (lane 2) or untreated (lane 3) HeLa cells.
BNIP3 expression enhanced the antitumor effects of irradiation in vitro
The clone formation experiments showed that at the same radiation doses, the surviving fraction of the pDsRed-BNIP3-transfected cells was decreased significantly (p < 0.01) compared with the control group, whereas that of the pDsRed-N1-transfected cells remained unchanged (p > 0.05). This indicated that the Bnip3 protein could inhibit clone formation of HeLa cells after irradiation (Fig. 3).

The clone formation experiments showed that the Bnip3 protein could inhibit clone formation of HeLa cells after irradiation. HeLa cells were transfected with pDsRed-N1 or pDsRed-BNIP3. After 24 hours, the cells were exposed to radiation and then immediately trypsinized and plated at the specified cell numbers. Colony formation efficiency was determined after 2 weeks. The results showed that the cloning efficiency of cells transfected with pDsRed-BNIP3 at different radiation doses was significantly lower (**p < 0.01) relative to that of the control and pDsRed-N1-transfected groups.
HeLa cell apoptosis as detected by Annexin V–FITC/PI
Figure 4 and Table 1 show that pDsRed-BNIP3-transfected cells treated with radiation showed a significantly increased apoptosis rate at both the early and late periods (p < 0.01) compared with the control groups and untransfected cells treated with radiation. Additionally, the analysis indicated that the total rate of apoptosis in pDsRed-BNIP3-transfected cells treated with radiation was significantly increased compared with the rate in untreated cells transfected with pDsRed-BNIP3 and in untransfected cells receiving radiation (p < 0.01), especially for late-period apoptosis. These results revealed that expression of the Bnip3 protein had a synergistic effect on apoptosis in combination with radiation treatment.

HeLa cell apoptosis as detected by Annexin V–FITC/PI. Quadrant 1, PI (+) (cells undergoing necrosis); Quadrant 2, Annexin V–FITC (+), PI (+) (cells in the late period of apoptosis and undergoing secondary necrosis); Quadrant 3, Annexin V–FITC (−), PI (−) (living cell); Quadrant 4, Annexin V–FITC (+), PI (−) (cells in the early period of apoptosis). The total apoptosis rate was calculated using the following equation: quadrant 2 + quadrant 4.
The pDsRed-BNIP3 + radiation (synergistic effect) group showed a significantly increased apoptosis rate (a p < 0.01) when compared with the control groups and untransfected cells treated with radiation. Its total rate of apoptosis was also significantly increased (b p < 0.01) when compared with the pDsRed-BNIP3 + radiation (additive effect) group, especially for late-period apoptosis. The result revealed that expression of the Bnip3 protein had a synergistic effect on apoptosis in combination with radiation treatment.
FITC, fluorescein isothiocyanate; PI, propidium iodide; SD, standard deviation; BNIP3, Bcl2/adenovirus EIB 19 kDa-interacting protein 3.
BNIP3 expression enhanced the antitumor effects of radiation in vivo
The effects of radiation in pDsRed-BNIP3-transfected cells were assessed in mice bearing cervical cancer. The tumor growth curves (Fig. 5) demonstrated that the combined treatment of pDsRed-BNIP3 and radiation significantly reduced the mean tumor volume on day 18 (measure the tumor volume every 3 days, in total seven times, beginning at the 10th day of injection of tumor cells) when compared with the radiation or pDsRed-BNIP3 treatment alone (p < 0.05) or the pDsRed-N1 plus radiation treatment (p < 0.05). The result shows that the antitumor effect of the combined treatment of pDsRed-BNIP3 and radiation was greater than their additive effect.

The tumor growth curves of nude mice bearing cervical cancer. Nude mice bearing cervical cancer were randomly divided into 6 groups and treated with NS, pDsRed-N1, pDsRed-BNIP3, radiation, pDsRed-N1 + radiation, or pDsRed-BNIP3 + radiation. The combined treatment involving pDsRed-BNIP3 and radiation significantly reduced the mean tumor volume when compared with the pDsRed-BNIP3, radiation, or pDsRed-N1 + radiation groups (n = 5; *p < 0.05, **p < 0.01).
Expression of BNIP3 combined with radiation increased the expression of BNIP3 in vivo
The immunohistochemical staining grades assigned to tumor tissue samples was determined as described by Shimizu et al. 22 As shown in Figure 6, tumor samples from pDsRed-BNIP3- and radiation-treated mice were determined to be strongly positive (+++) for Bnip3 expression, samples from pDsRed-BNIP3 mice were moderately positive (++), and all other groups were only faintly positive (+). This analysis indicated that the Bnip3 protein was expressed in the cytoplasm of tumor cells and was especially prominent in cells adjacent to the tumor tissues undergoing necrosis. This was likely due to the high expression of hypoxia inducible factor-1 surrounding these tissues, as previously shown. 23

Immunohistochemical analysis of BNIP3 in the tumor tissues.
BNIP3 expression enhances radiation-induced apoptosis in vivo
The results of TUNEL assays indicated that cell apoptosis in cervical cancer HeLa xenografts were significantly different between the groups. As shown in Figure 7, increased numbers of apoptotic cells were observed in groups expressing recombinant BNIP3 and/or receiving radiation treatment, compared with the control and pDsRed-N1-treated groups. Importantly, tumors from animals treated with pDsRed-BNIP3 and radiation had the highest apoptotic rate when compared with the control group (Fig. 7). As such, these data confirmed the synergistic effect of BNIP3 expression in combination with radiation treatment on tumor apoptosis rates.
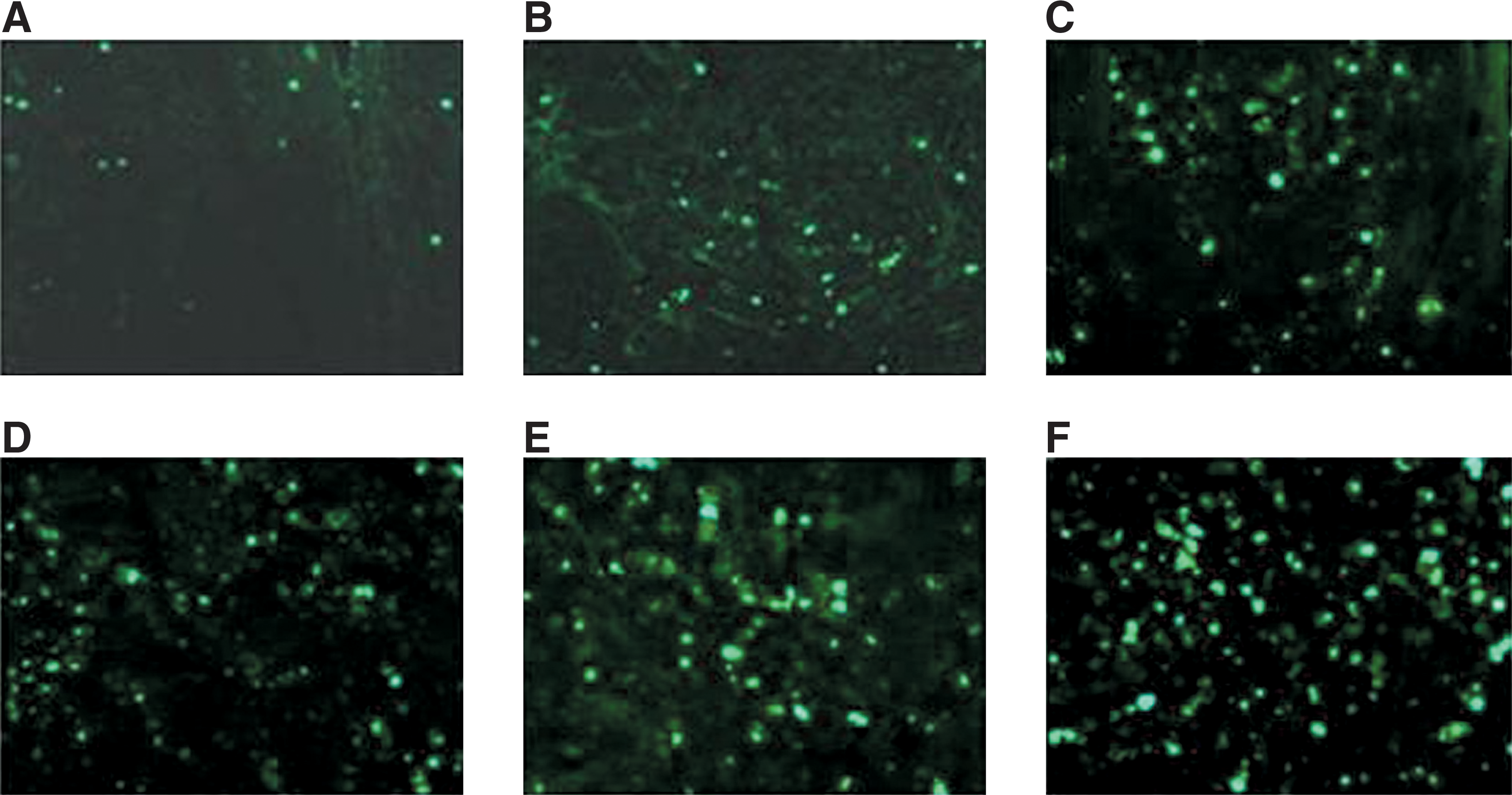
Cells undergoing apoptosis in the tumor tissues were identified via TUNEL assay. NS

Discussion
Bnip3 is a mitochondrial protein that induces apoptosis when transiently overexpressed, and it is classified among the BH3-only Bcl-2 family of proteins. 24 –26 The Bnip3 protein functions to open the mitochondrial permeability transition pore and release cytochrome c from the mitochondrial intermembranous space into the cytoplasm and, subsequently, promote apoptosis. 10,27,28 During radiotherapy, radiation-induced apoptosis is one of the primary mechanisms of radiation-induced cell death. Radiation directly causes DNA damage and destroys the mitochondrial membrane potential, bringing about mitochondrial damage, and it also induces apoptosis by releasing cytochrome c. 29 –31 There are common links between Bnip3-induced apoptosis and radiation-induced apoptosis, and they may interact to increase the rate of apoptosis when present in combination. However, Bnip3 expression was decreased in most tumor tissues compared with healthy tissues. As previously shown, approximately half of all cervical cancers express low levels of the Bnip3 protein. Radiotherapy is the main treatment for cervical cancer, and the absence of Bnip3 expression may cause radioresistance as well as chemoresistance, as shown in other cancers. 12,16 –19
In the present study, to determine the potential of BNIP3 as a therapeutic target for radiosensitization, BNIP3 was overexpressed in cervical cancer in vitro and in vivo and the presence of apoptotic cells was detected by flow cytometry and TUNEL assay. The transient transfection with pDsRed-BNIP3-induced apoptosis of the cervical cancer cells in vitro, and the apoptosis rate of the combination treatment (radiation combined with pDsRed-BNIP3) increased when compared with pDsRed-BNIP3 or radiation alone, especially that of late-period apoptosis. Further, the present study showed that the increased expression of BNIP3 significantly enhanced the effects of radiation treatments in vivo. The findings indicated that BNIP3 is an attractive therapeutic target because of its ability to suppress tumor growth and enhance radiosensitivity in cervical cancer.
In the present study, it was able to achieve increased expression of Bnip3 in cervical cancer cells by using the pDsRed-BNIP3 treatment strategy. The increased expression of Bnip3 induced the strong suppression of clone formation in the HeLa cervical cancer cell line and increased the apoptosis rate of these cells, as detected by AnnexinV-FITC/PI double staining. This result suggested that Bnip3 is an essential molecule for induction of tumor cell apoptosis and a good target for halting proliferation and triggering apoptosis. More strikingly, overexpression of BNIP3 strongly inhibited tumorigenesis in mice cervical cancer xenografts. This result suggested that the absence of BNIP3 expression is strongly associated with the in vivo tumorigenic ability of cervical cancer, as shown in a previous study on colorectal cancer. 11 These observations lead to the expectation that treatment with exogenous BNIP3 may be useful for stopping the progression of cervical cancer in vivo.
Further, in attempting to determine whether BNIP3 is a good therapeutic target for radiosensitization, BNIP3 expression was increased in cervical cancer cells both in vitro and in vivo, and subsequently, the presence of apoptotic cells was detected by flow cytometry and TUNEL assays. It was found that expression of the BNIP3 gene synergistically enhanced the apoptotic effect of radiotherapy. The data showed that transient transfection with pDsRed-BNIP3-induced apoptosis of cervical cancer cells in vitro. Further, the apoptosis rate of the combination treatment (radiation combined with pDsRed-BNIP3) was significantly increased when compared with pDsRed-BNIP3 or radiation treatment alone, and this was even higher than the collective apoptosis rates of the pDsRed-BNIP3 group and the radiotherapy group. This was especially true for the late-period apoptosis rates (p < 0.01). It was also shown that the increased expression of BNIP3 significantly enhanced the effects of radiation in vivo. TUNEL assays were used to detect tumor cell apoptosis in the tumor tissues. The results indicated that expression of BNIP3 combined with radiation had a higher apoptosis index relative to that of cells treated with BNIP3 or radiation alone. The apoptosis index of cells receiving the combined treatment was even higher than the collective apoptosis rates of the pDsRed-BNIP3 group and the radiotherapy group. The findings indicated that BNIP3 is an attractive candidate for therapeutic targeting because of its suppression of tumor growth and enhancement of radiosensitivity in cervical cancer.
Conclusions
In conclusion, expression of BNIP3 sensitized human cervical cancer cells to ionizing radiation both in vitro and in mouse xenografts, and combined treatment with BNIP3 and radiation produced a synergistic effect on tumor apoptosis rates. However, the exact mechanism by which BNIP3 expression increased radiation-induced apoptosis requires further study. As such, subsequent clinical trials aimed at evaluating the radiosensitization effect of BNIP3 expression on cervical cancer are warranted.
Footnotes
Disclosure Statement
The authors declare that no competing financial interests exist.