
Abstract
Introduction:
Radioiodine is a routine therapy for differentiated thyroid cancers. In principle, undifferentiated thyroid cancers as well as nonthyroid cancers can concentrate and, thus, be treated with radioiodine after transfection with the human sodium iodide symporter (hNIS) gene. The human telomerase reverse transcriptase (hTERT) promoter is an effective tumor-specific promoter of gene expression and, thus, may be useful in targeted gene therapy of cancer.
Methods:
We used hTERT promoter-modulated expression of the hNIS and human thyroperoxidase (hTPO) genes in an experimental model of radioiodine-based treatment of malignant glioma. Cells were cotransfected by adenovirus in which the hNIS gene had been coupled to the hTERT promoter and the hTPO gene had been coupled to the human cytomegalovirus (CMV) promoter (Ad-hTERT-hNIS and Ad-CMV-hTPO, respectively), and they were evaluated in cells thus transfecting transgene expression by western blots, 125I uptake and influx, and clonogenecity after 131I treatment.
Results:
After cotransfection with two adenovirus, cells showed about 31–34 times higher 125I uptake than the control cells transfected with Ad-CMV-EGFP (enhanced green fluorescent protein) and almost 1.3–1.4 times higher 125I uptake than cells only transfected with Ad-hTERT-hNIS. Western blots revealed two bands of ∼70 and 110 kDa, respectively. The in vitro clonogenic assay indicated that, after exposure to 100–1000 μCi of 131I-iodide for 12 hours, 91%–94% of cells cotransfected with the hNIS and hTPO genes, 88%–93% of cells transfected with the hNIS gene, and only 62%–68% of control (nontransfected) cells were killed.
Conclusions:
The experiments demonstrated that an effective therapy of 131I was achieved in malignant glioma cell lines after induction of tumor-specific iodide uptake activity by the hTERT promoter-directed NIS expression in vitro. Cotransfection of the hNIS and hTPO genes can lead to longer retention of radioiodide, but did not increase cell killing over that achieved with transfection with the hNIS gene alone.
Introduction
Malignant glioma is the most commonly occurring primary malignant brain tumor. It is difficult to treat and usually associated with an inexorable, rapidly fatal clinical course, with a life expectancy of less than 1 year. 1,2 Current therapeutic modalities include surgery, radiotherapy, interstitial brachytherapy, chemotherapy, temozolomide, and so on. 3 Tumor-specific promoters should enhance the specificity and, thus, the effectiveness of tumor targeted therapeutic transgenes, although such promoters are generally less effective than nonspecific or constitutive promoters such as the cytomegalovirus (CMV) and simian virus 40 (SV40) promoters. 4
Telomerase is a DNA polymerase, and it is highly active in most immortalized cell lines and over 85% of human cancers but inactive in most somatic cells. 5 It has become a target of anticancer therapy. The promoter region of the human telomerase reverse transcriptase (hTERT) gene has already been cloned. The promoter is highly G/C-rich and lacks both TATA and CAAT boxes but contains binding sites for several transcriptional factors, including Myc and Sp1. 4 The cytotoxic effect is restricted to telomerase-positive tumor cells. The core promoter can also be activated in cancer cell lines. Recent investigations have demonstrated that the gene therapy using the hTERT promoter-driven expression vector of therapeutic gene such as caspase-8, bax, diphtheria toxin A, thymidine kinase, rev-caspase-6, FADD, and TRAIL can be tested successful. 4,6 –11 The hTERT promoter is, therefore, a promising tumor-selective promoter with potential application in targeted cancer gene therapy. 12,13
The human sodium iodide symporter (hNIS) belongs to the sodium/solute symporter family. It is an intrinsic membrane glycoprotein with 13 putative transmembrane domains, an extracellular NH2 terminus, and an intracellular COOH terminus. The hNIS mediates Na+/K+ ATPase dependent active transport of iodide across the membrane of thyroid follicular cells. 14 By targeted transfection with the hNIS gene, undifferentiated thyroid cancer as well as nonthyroid cancers will take up iodide and, thus, can potentially be treated with radioiodine. Radioiodine treatment also exerts a so-called bystander effect as a result of radiation cross-fire and may kill neighboring non-hNIS-expressing cells as well as hNIS-expressing, iodide-concentrating cells. 15 Tumor-specific transfection of the hNIS gene has been applied to a variety of tumors, including thyroid, prostate, colon, breast, liver, and lung cancer. 16 –19 Importantly, however, the hNIS expression in nonthyroidal cells results only in rapid internalization but not organification of radioiodide. Radioiodine, therefore, rapidly exits hNIS-transfected nonthyroidal cells, thus reducing the radiation dose to such cells. This is the major limitation of hNIS-mediated radioiodine therapy of cancer.
The human thyroperoxidase (hTPO) is the primary enzyme in thyroid hormone synthesis, catalyzing iodide oxidation and iodination of tyrosine residues. 20 Cotransfection of the hNIS and hTPO genes enhances both internalization and retention of radioiodide and, thus, enhances cell killing, overcoming the limitation of rapid efflux of activity from nonthyroidal cells transfected only with the hNIS gene. 21
The hTERT promoter can restrict expression of transfected hNIS and hTPO genes to telomerase-positive tumor cells and, thus, maximize tumor-specific uptake and minimize nonspecific uptake of radioiodine. In the current study, therefore, we developed and evaluated in glioma cell lines in vitro the potential functional and therapeutic effectiveness of an adenovector incorporating the hNIS and hTPO genes and the hTERT promoter.
Materials and Methods
Cell lines
The U251 and U87 human glioma cells and MRC-5 human lung fibroblasts cells were cultured in Dulbecco's modified Eagle's medium (DMEM) (Gibco BRL) with 10% fetal bovine serum (FBS) and 100 U/mL penicillin/streptomycin. All cells were grown at 37°C under 5% CO2 in air.
Real-time PCR analysis for the hTERT gene
The expression of the hTERT mRNA was analyzed by real-time PCR. Total RNA from cells was isolated by using TRIzol reagent (Invitrogen), and reversed to single-stranded cDNA by using PrimeScript RT-PCR Kit (TaKaRa). Real-time PCR was conducted as previously described. 22 Briefly, it was carried out in 96 sample tubes/assay, using SYBR Premix Ex Taq (Perfect Real Time) (TaKaRa). All PCR were developed using Stepone plus RT PCR system (ABI), and the relative expression was calculated using the ΔCT method and normalized to the expression of the β-acting mRNA. 23 The primers for the hTERT gene were as follows: forward primer 5′-CCACCTTGACAAAGTACAG-3′ and reverse primer 5′-CGTCCAGACTCCGCTTCAT-3′. The primers for β-acting gene were as follows: forward primer 5′-TCATGAAGTGTGACGTGGAC-3′ and reverse primer 5′-ACCGACTGCTGTCACCTTCA-3′. Results represented an average of three independent experiments.
Cloning of the hTERT promoter
The hTERT promoter region was amplified by PCR using LA-Taq polymerase (TaKaRa) from human genomic DNA. The hTERT promoter introduced both BglII and HindIII restriction sites at the 5′ and 3′ ends. The hTERT promoter 204 occupied the region from-204 to +56 (GenBank AF015950), which contained core promoter and two E-boxes. 24 The hTERT promoter 378 occupied the region from −378 ∼+78. 24 The hTERT promoter 1375 occupied the region from −1375 ∼+78, which contained all long promoter regions. 24 The PCR conditions were used for amplification (1 cycle: 94°C 2 minutes; 30 cycles: 94°C 30 seconds, 55°C 30 seconds, 70°C 1 minutes; 1 cycle: 72°C 10 minutes and hold 4°C).The PCR product was cloned into pMD-18T vector (Takara) and digested with BglII and HindIII. Then, it was ligated with PGL3-Basic vector (Promega),digested by BglII and HindIII, and named as PGL3-204, PGL3-378, and PGL3-1375.
Luciferase assay
The expression of luciferase gene by different regions of the hTERT promoter in tumor cells was determined by Dual-Glo Luciferase Assay System (Promega) according to the manufacturer's instructions. Briefly, cells were seeded in a 24-well plates that were exposed by transfection with recombinant luciferase reporter plasmids PGL3-204, PGL3-378, and PGL3-1375, and background control plasmid vector pRL-TK (Promega) for 6 hours at 37°C. The cells were harvested 24 hours after the transfection. Then, Luciferase assays were performed by Safire2 microplate reader (TECAN). The control vector (PGL3-control) that contains SV40 promoter was used as a positive control, and PGL3-Basic without promoter was used as a negative control. All experiments were done in triplicate.
Construction of recombinant adenovirus
The hNIS gene (sense: 5′-CCATGGATGGAGGCCGTGGAG-3′, antisense: 5′-TCTAGACTCCTGCTGGTCTCG-3′) was PCR cloned from full-length hNIS cDNA (GeneCopoeia) with NcoI and XbaI restriction sites at the 5′ and 3′ ends, then ligated with PGL3-hTERT plasmid digested by NcoI and XbaI, and named as PGL3-hTERT-hNIS. The PGL3-hTERT-hNIS contained the hTERT promoter 204 and the hNIS gene. Then, the fragment, which carried the hNIS gene under the control of the hTERT promoter, was PCR cloned from PGL3-hTERT-hNIS with XhoI and SalI restriction sites at the 5′ and 3′ ends, and inserted in a pShuttle vector. The pShuttle-CMV-hNIS was constructed by inserting the hNIS gene with XhoI and SalI restriction sites at pShuttle-CMV vector.
Cells transfected only with the hTPO gene cannot take up iodide, and this gene was, therefore, expressed under the control of the CMV promoter. The hTPO gene (sense: 5′-GCGGCCGCCACCATGAGAGCGCTCGCTGTGCT-3′, antisense: 5′-AAGCTTACTAATGATGATGATGATGATGGAGGGCTCTCGGCAGCCTGTGAGTATCCC-3′) was PCR cloned from pcDNA-TPO plasmid (previously constructed) with NotI and HindIII at the 5′ and 3′ ends and inserted in pShuttle-CMV vector.
Ad-hTERT-hNIS, Ad-CMV-hTPO, and Ad-CMV-hNIS were produced according to the protocol provided by the manufacturer of the AdEasy™ Adenoviral Vector System (Stratagene). The recombinant virus was stored at −80°C until used. Ad-CMV-EGFP (enhanced green fluorescent protein) is a control vector unrelated to iodine uptake and metabolism.
Transfection of U251, U87 cell lines by adenoviral infections in vitro
The cells were seeded in six-well plates to obtain between 5×105 and 1×106 cells per well at the time of infection, infected with multiplicity of infection (MOI) (i.e., the ratio of the number of viral particles to the number of cells) of 50 for Ad-hTERT-hNIS, Ad-CMV-hTPO, and Ad-CMV-hNIS in 1000 μL of serum-free medium for 6 hours, and followed by addition of 10% FBS new medium. After transfection, cells were incubated for 24 hours.
Western blot analysis
For western blot analysis, the EPS 2A200 electrophoresis system (Amersham Biosciences) was used. 25 Cells were harvested with a scraper after being washed with ice-cold phosphate-buffered saline (PBS) and lysed in a buffer containing 10 mM Tris·HCl (pH 7.5), 1 mM DTT, 20% (v/v) glycerol, 1 mM ethylenediaminetetraacetic acid, and a protease inhibitor mixture. Samples were then centrifuged at 4°C for 10 minutes 26 , and 40-μL aliquots were prepared for electrophoresis by addition of NuPAGE sodium dodecyl sulfate sample buffer. Samples were electrophoresed in bis-Tris·HCl buffered 4%–12% gradient polyacrylamide gels (Invitrogen). After gel electrophoresis at 140 V for 1 hour, proteins were transferred to nitrocellulose membranes using electroblotting. After blotting, membranes were preincubated for 8 hours in 5% low-fat dried milk in TBS-T to block nonspecific binding sites. Membranes were then incubated separately with a goat polyclonal antibody raised against the N-terminal extracellular domain of NIS of human origin (1:200 dilution, sc-48055; Santa Cruz), a mouse monoclonal His-tag antibody raised against the hTPO (1:1000 dilution, clone 4D 11; Millipore), a mouse monoclonal antibody raised against the β-acting (1:500 dilution, TA-09; Zhongshan Goldenbridge Biotechnology), and a goat polyclonal antibody raised against the C-terminus of hTERT of human origin (1:400 dilution, sc-7215; Santa Cruz) for 2 hours at room temperature. After washing with TBS-T, horseradish peroxidase-labeled goat anti-rabbit antibody was applied (1:2000 dilution, sc-2004; Santa Cruz) or mouse anti-rabbit antibody was applied (1:2000 dilution, sc-2357; Santa Cruz) for 2 hours at room temperature before incubation with Pierce ECL western blotting Substrate (Thermo) for 1 minute. Exposures were made at room temperature for ∼1–3 minutes using Fujifilm films. Prestained protein molecular weight standards (Spectra Multicolor Broad Range Protein Ladder, SM1841; Fermentas) were run in the same gels for comparison of molecular weight and estimation of transfer efficiency.
Iodide uptake and efflux assays
Cells were seeded in six-well plates and infected with recombinant adenovirus for 6 hours, then placed in fresh DMEM with 10% FBS, and incubated for an additional 24 hours. 14 The radioactivity was measured with a γ counter (LKB gamma 1261; LKB Instruments). To measure the 125I uptake, 5×105–1×106 cells per well were cultured with 1 mL 10% FBS-DMEM (contained 0.5 μCi Na125I) for 0, 10, 20, 30, and 40 minutes. Then, the 125I-containing medium was decanted, cells were washed twice with PBS, lysed with 0.3 M sodium hydroxide, and counted. To measure the 125I efflux, 5×105–1×106 cells per well were cultured for 1 hour with 1 mL 10% FBS-DMEM (contained 0.5 μCi Na125I), then the 125I-containing medium was decanted. After cells had been washed twice with PBS, fresh nonradioactive medium was added to the six-well plates. Then, the cells were again cultured for 0, 5, 10, 15, and 20 minutes. Thereafter, the cells were washed, lysed, and counted. All experiments were done in triplicate.
In vitro cell killing with 131I and clonogenic assay
Cells were seeded in 24-well plates and infected with 50 MOI of adenovirus for 6 hours and then in fresh DMEM with 10% FBS for an additional 24 hour, washed twice with PBS, and incubated with 1 mL of DMEM with 10% FBS containing 100, 500, or 1000 μCi 131I. 17,27 After 12 hours of incubation, cells were washed twice with PBS. For each condition (Ad-hTERT-hNIS infected, Ad-CMV-hNIS infected, Ad-hTERT-hNIS and Ad-CMV-hTPO infected, Ad-CMV-hTPO and Ad-CMV-hNIS infected and Ad-CMV-EGFR [the control adenovirus] infected), cells were plated in 24-well plates at densities of 100 cells/well and incubated for 1 week at 37°C. Then, the cells were washed twice with PBS, fixed with 0.5 mL Carnoy's solution (a freshly prepared 3:1 mixture of methanol and acetic acid), and stained with a crystal violet solution (for 250 mL, 0.5 g of crystal violet, 25 mL of 40% formaldehyde, 50 mL of ethanol, and 175 mL of water). Colonies of >20 cells were counted. All experiments were done in triplicate.
Statistical analysis
All experiments were performed in triplicates unless otherwise indicated. Statistical analysis was done using SPSS software (SPSS 13.0). The results are presented as mean±SD. Statistical significance was tested using the Student's t-test procedure.
Results
Expressions of the hTERT gene in U251, U87, and MRC-5 cell lines
The RT-PCR and western blot analysis showed that the hTERT gene was expressed in telomerase-positive U251 and U87 tumor cell lines, and was not expressed in the telomerase-negative MRC-5 cells (Fig. 1). Real-time PCR analysis confirmed that expression of the hTERT mRNA in U251 and U87 was similar, and the hTERT mRNA of U87 cells appeared to express slightly higher amounts of the hTERT transcripts than U251 (Table 1). 28

The expression of the hTERT and β-acting genes in tumor and normal cell lines. U251 and U87 were teloemerase-positive tumor cell lines, and MRC-5 was a telomerase-negative cell line. The PCR products were run on 1% agarose gel containing 0.5 μg/mL ethidium bromide. β-acting gene was used as a positive control and expressed in U251, U87, and MRC-5 cells
Data were expressed as mean (SD) of three experiments.
p<0.01 versus β-acting.
CT is the threshold cycle for target amplification, such as hTERT and β-acting genes expression in U87 and U251 cells. ΔCT is equal to the difference in threshold cycles (CT) for target (hTERT) and reference (β-acting), ΔCT=CThTERT−CTβ-acting, ΔCTU251 =7.05±0.52, ΔCTU87=7.22±0.12. ΔΔCT calculates the relative quantification of target, the U251 cells are chosen as the calibrator in Table 1, and the amount of hTERT mRNA in U87 is relative to U251. So, ΔΔCTU251=(7.05±0.52)−7.05=0±0.52, and ΔΔCTU87=ΔCTU87−ΔCT U251=(7.22±0.12)−(7.05±0.52)=0.17±0.12. The expression of the hTERT mRNA in U87 was slightly higher than U251, as the data showed that the hTERT amount relative to U251 was 1.0 (0.7–1.4) in U251 and the hTERT amount relative to U251 was 1.1 (1.0–1.2) in U87.
hTERT, human telomerase reverse transcriptase.
Luciferase assay
To assess the cell-specific transcriptional activity of the hTERT promoter, a reporter gene assay using luciferase was performed in transiently transfected cells. The transient transfection showed that the hTERT promoter could lead to luciferase gene expression in telomerase-positive U251 and U87 tumor cell lines, without expression in the telomerase-negative MRC-5 cells. However, the hTERT promoter 204, 378, and 1375 had different transcriptional activities, and the transcriptional activity of the hTERT promoter 204 was the most efficient among them with the relative luciferase activities of the hTERT promoter 204 in U87 and U251 reaching 50% of that in positive-control cells with the SV40 promoter (Fig. 2).These results indicated that the hTERT promoter 204 is the strongest and the most efficient promoter in U251 and U87 cell lines.
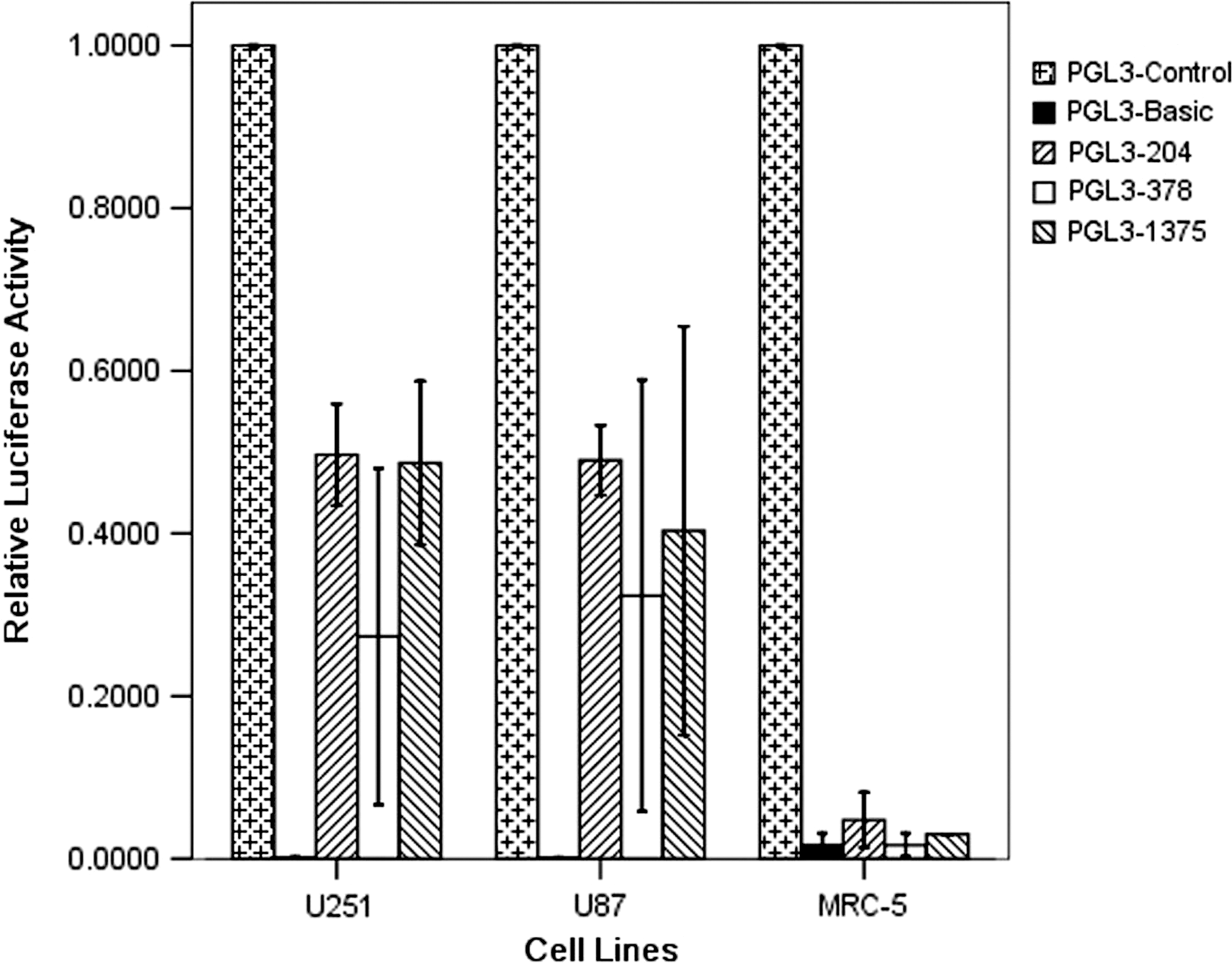
Transcriptional activity of the hTERT promoter 204, 378, and 1375 in tumor and normal cell lines. We compared the luciferase expression of reporter plasmid PGL3-204, PGL3-378, and PGL3-1375 inferring the hTERT promoter 204, 378, and 1375's transcriptional activities. Luciferase activity of reporter plasmid PGL3-204, PGL3-378, and PGL3-1375 with the hTERT promoter 204, 378, and 1375 were transiently transfected in cell lines U251, U87s and fibroblast cell line MRC-5. Luciferase activity ratio in each plasmid was compared with background control pRL-TK vector at first. Relative luciferase activity (RLA) for each experimental treatment was calculated by using this formula:
Western blot analysis
The protein level of the hNIS and hTPO genes was analyzed by western blotting. In U87 and U251 cells, the hNIS protein was detected as a major band corresponding to a molecular weight at 70 kDa. The hTPO protein was a band corresponding to a molecular weight of at 110 kDa, with hNIS and hTPO protein expression lower than that of the control tubulin protein. The activity of hTERT promoter was weaker than that of the CMV promoter (Fig. 3)
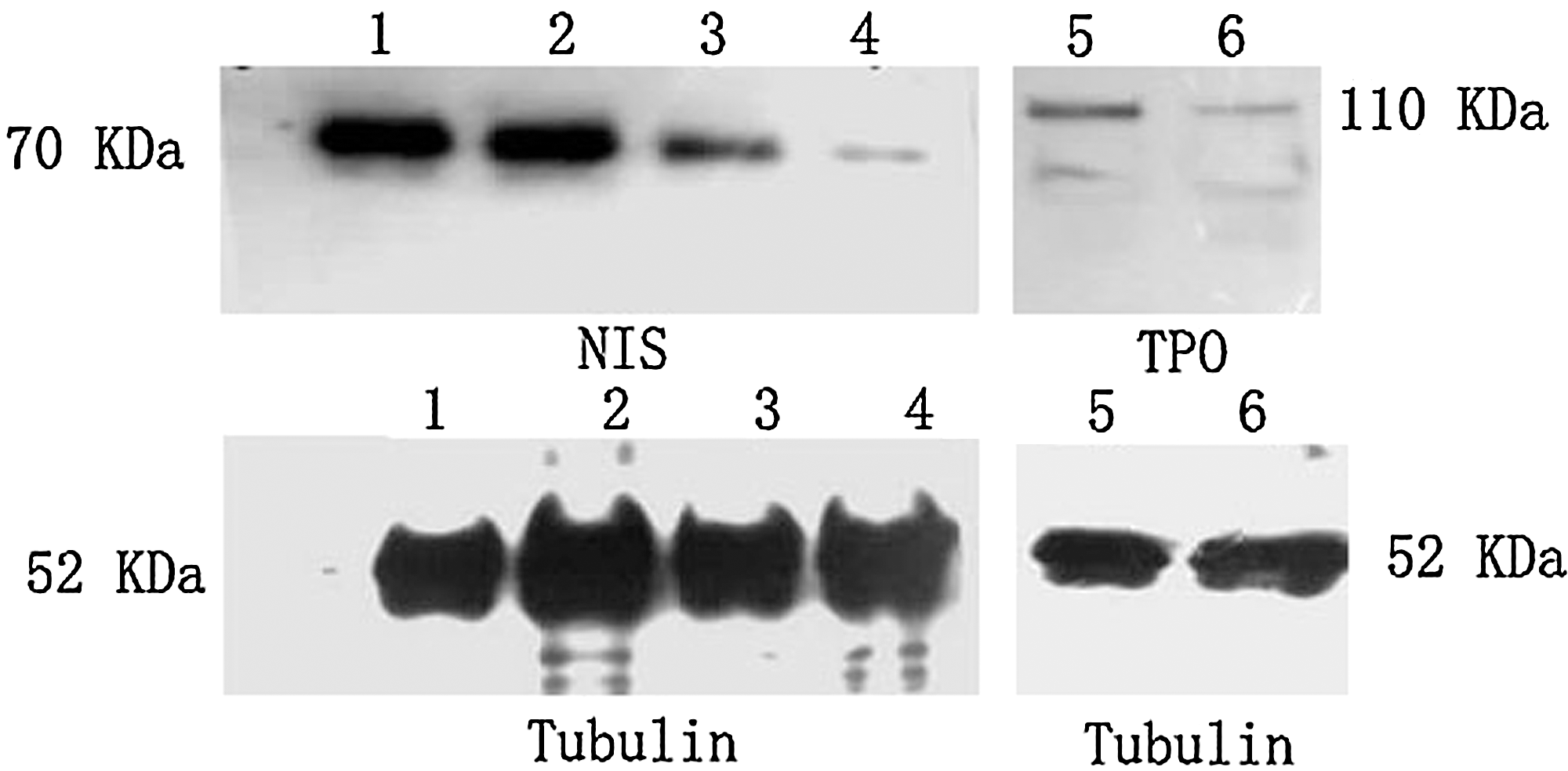
Western blot analysis for hNIS and hTPO protein levels. 1, Ad-CMV-hNIS infected U87 cells; 2, Ad-CMV-hNIS infected U251 cells; 3, Ad-hTERT-hNIS infected U87 cells; 4, Ad-hTERT-hNIS infected U251 cells; 5, Ad-CMV-hTPO infected U87 cells; 6, Ad-CMV-hTPO infected U251 cells. The hTPO and hNIS proteins were observed as major bands corresponding to molecular weights of 110 and 70 kDa, respectively, with much more lower levels in both U87 and U251 cells than those of tubulin protein. The hNIS protein expressed under control of the CMV promoter (1, 2) was higher than that expressed under control of the hTERT promoter (3, 4). The exposures of the NIS and TPO blots were 3 and 1 minute, respectively. hNIS, human sodium iodide symporter; hTPO, human thyroperoxidase; CMV, cytomegalovirus.
Iodide uptake and efflux assays
To determine the iodide uptake and efflux, we performed 125I timed activity measurements in hNIS- and hTPO-transfected and control (Ad-CMV-EGFP-transfected) U251 and U87 human glioma cells. Radioiodide uptake in Ad-CMV-hNIS- and Ad-CMV-hTPO-transfected U251 and U87 cells was rapid, reaching maximal levels within 30 minutes, and higher than that in control cells (Fig. 4A, B). To determine the iodide efflux, the iodide uptake was permitted to proceed for 1 hour, when the steady-state level of accumulation was achieved. After replacement of the 125I-containing medium with nonradioactive medium, the intracellular iodide was continuously released to the medium, and a rapid efflux of radioiodine from cells was evident, but 125I had a longer effective half-life in cells cotransfected with the hNIS and hTPO genes than in cells transfected with the hNIS gene alone (Fig. 4C, D). 125I has a long physical half-life, 60 days, and so the effect of decay over the 20–50 minutes duration of the foregoing experiments can be ignored.

Iodide uptake, efflux assays, and iodide effective half-life in U87 and U251 cell lines.
In vitro cell killing with 131I and clonogenic assay
Using the vitro clonogenic assay, we investigated whether 131I showed selective cytotoxic activity in the hNIS and hTPO genes expressing cells (Fig. 5). After being incubated with 100, 500, or 1000 μCi 131I for 12 hours, the survival rates of U251, U87 cells, and MRC-5 cells were reduced in a dose-dependent manner. The survival rates of U251, U87 tumor cells, and MRC-5 cells transfected with CMV promoter adenovirus were all lower than those of cells transfected with hTERT promoter adenovirus. Since hTERT promoter could not express the hNIS gene in telomerase-negative MRC-5 cells, the survival rates of cells transfected with hTERT promoter adenovirus and those of cells transfected with Ad-CMV-EGFR (control empty adenovirus) were similar.
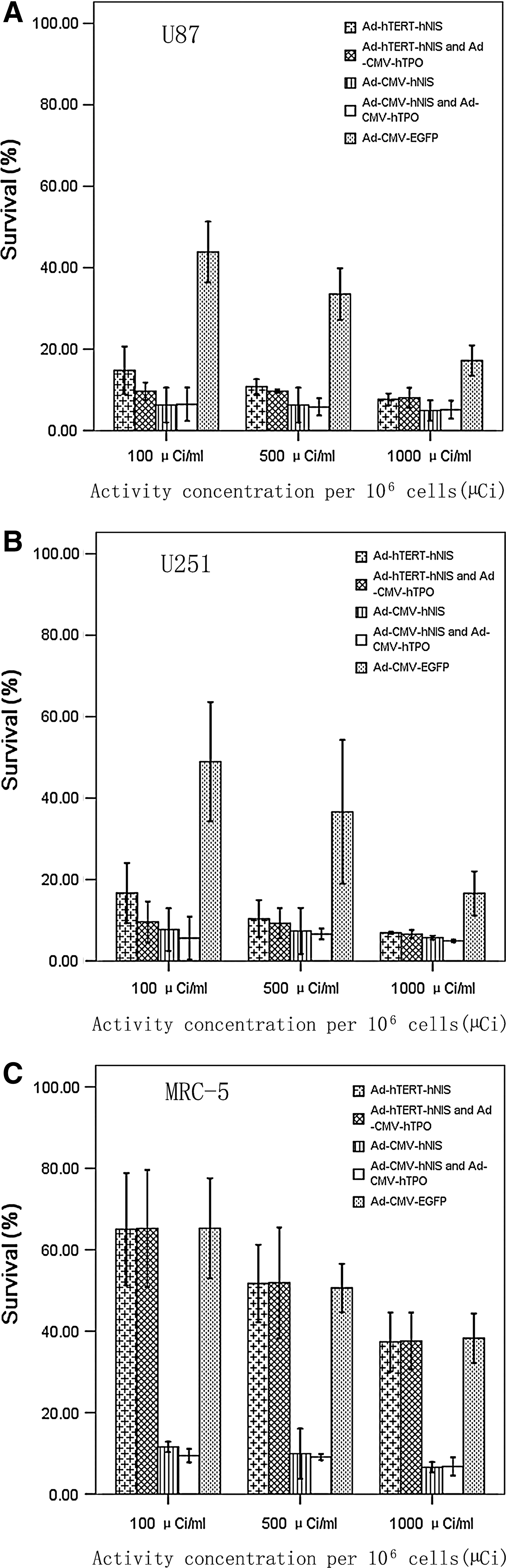
The vitro clonogenic assay of
The survival rates of U87 and U251 cells (treated with 100 μCi 131I) cotransfected with Ad-hTERT-hNIS and Ad-CMV-hTPO were slightly lower than those of cells transfected by Ad-hTERT-hNIS, but the survival rates of U87 and U251 cells cotransfected with Ad-CMV-hNIS and Ad-CMV-hTPO and transfected by Ad-CMV-hNIS were not significantly different (U87: p=0.91, U251: p=0.45). The survival rates of U251 and U87 (treated with 500, 1000 μCi 131I) cotransfected with the hNIS and hTPO gene and transfected by hNIS gene alone were also similar (p>0.05). The survival rates of transfected U251 and U87 cells not incubated with 131I were almost 90% (not showed in Fig. 5). These data indicated that transfection with the hNIS, whether with the hTERT or CMV promoter, resulted in more 131I-induced cell killing than without hNIS gene transfection, but that cotransfection with the hNIS and hTPO genes did not increase 131I-induced cell killing.
Discussion
Malignant gliomas represent one of the most aggressive forms of brain cancer. 1 Malignant gliomas (glioblastoma multiforme and anaplastic astrocytoma) comprise the most common types of primary central nervous system tumors. 29 Even with aggressive multimodality therapy, the overall long-term prognosis of patients with these tumors remains poor. 30 New therapeutic strategies being investigated for glioblastoma include signal-transduction pathway inhibitors (including those mediated by the epidermal growth factor receptor, Ras/MAPK, and PI3K), the anti-VEGF monoclonal antibody Bevacizumab, adoptive, passive, and active (i.e., vaccine based) immunotherapy, and Herpes simplex virus-thymidine kinase (HSV-tk) gene therapy. 31,32 However, none of these strategies has yet to improve the survival of patients with glioblastoma.
To improve selectivity, gene-therapy based on the specific expression of transfected genes in gliomas cells is currently being investigated. It presumably would have maximal tissue-specific cytotoxity and minimal extratumoral side-effect. Tumor-selective promoter, such as the hTERT promoter, is used for achieving such selective expression of therapeutic genes. 33
The hNIS mediates efficient iodide uptake by thyroid cells and, thus, is responsible for radioiodine-based diagnostic imaging and therapy of thyroid carcinomas and their metastases, and has been cloned from a human thyroid cDNA library in 1996. 34 Targeted expression of the functional hNIS gene in cancer cells would enable these cells to concentrate iodide and offer the possibility of radioiodine therapy. 19,21,35 Many researchers have reported that the hNIS gene transferred to various cancer cells and increased radioiodine uptake by such cells relative to that of nontransfected control cells. However, they also found that cancer cells transfected with the hNIS gene showed a rapid loss of radioiodine once radioiodine had been removed from the medium. It was subsequently reported that cotransfection with hNIS and hTPO genes would increase retention as well as uptake of radioiodine and result in greater radioiodine induced cell death. 21 The hTPO gene is a glycosylated hemoprotein enzyme located at the apical plasma membrane of the thyroid follicle cell, and it plays a key role in thyroid hormone biosynthesis. 36 There is a balance between hNIS-mediated iodide influx and hTPO-inhibited efflux. However, Boland et al. 37 found that coinfection of nonthyroid tumor cells with an adenoviral vector for hNIS and hTPO could lead to the significant increase of iodide organification in the presence of hydrogen peroxide, but the levels of iodide organification were too low to significantly increase the iodide retention time in the target cells.
In the current study, we constructed and evaluated in U251 and U87 glioma cell lines in vitro the potential functional and therapeutic effectiveness of an adenovector incorporating the hNIS and hTPO genes and hTERT promoter. Some researchers found that cotransfection with the hNIS and hTPO genes may be one way of inhibiting rapid intracellular iodide efflux after radioiodine had been removed from the medium. Huang et al. 21 found that cells cotransfected with the hNIS and hTPO genes would increase radioiodine uptake and retention, and lead to greater tumor cell death. Conversely, in 2002, Boland et al. confirmed that nonthyroid tumor SiHa cells coinfected with AdNIS and AdTPO could increase the iodide organification level, but the addition of exogenous H2O2 in the medium was required. Moreover, even in the presence of exogenous hydrogen peroxide, the levels of iodide organification were low and insufficient to significantly increase the iodide retention time in the tumor cells. Boland et al. 37 suggested that the efficiency of the iodide retention process might be cell-type dependent. In the current study, we found that cells cotransfected with the hNIS and hTPO genes under the control of either the hTERT or CMV promoter exhibited increased radioiodine uptake and retention: the maximal 125I iodide uptake of cells cotransfected with Ad-CMV-hNIS and Ad-CMV-hTPO, cotransfected with Ad-hTERT-hNIS and Ad-CMV-hTPO, U87 cells was 74- and 31-fold higher, respectively, for U87 cells and 65- and 34-fold higher, respectively, for U251 cells than for the corresponding control cells (i.e., transfected with Ad-CMV-EGFR). There was no significant difference in the survival rates of 131I-treated U251 and U87 cells cotransfected with the hNIS and hTPO genes or transfected by hNIS alone, except that the survival of cells cotransfected with Ad-hTERT-hNIS and Ad-CMV-hTPO was slightly lower than that of cells transfected by Ad-hTERT-hNIS treated with 100 μCi 131I group.
By targeted transfection with the hNIS gene, undifferentiated thyroid cancer as well as nonthyroidal cancers will take up iodide and, thus, can potentially be treated with radioiodine. However, hNIS also mediates the uptake of anions other than iodide, including perchlorate, perrhenate, bromide, and astatide. 38,39 Thus, for example, perrhenate labeled with the beta-particle emitter 188Re and the alpha-particle emitter 211At may be therapeutically used with hNIS gene-transduced cancer cells. By cross-fire of beta rays, both 131I and 188Re treatment may exert a bystander effect, with killing of neighboring non-hNIS-expressing cells as well as hNIS-expressing cells. On the other hand, although the short-range alpha particles emitted by 211At do not have a significant cross-fire effect, they have a high linear energy transfer and relative biological effectiveness and, thus, should induce more cell killing than 131I and 188Re for far lower activities concentrated per hNIS-expressing cell.
In conclusion, significant 131I-iodide-induced killing of U251 and U87 human glioma cells transfected with the hNIS and tumor-specific hTERT promoter genes has been demonstrated in vitro. However, although cotransfection with hNIS and hTPO genes increased the uptake and retention of 131I-iodide, it did not increase 131I-iodide-induced cell killing. Further study and optimization of radioiodine therapy of cancer after hTERT tumor-specific hNIS gene transfections are, therefore, clearly needed.
Footnotes
Acknowledgments
The authors thank Dr. Ning Li for his contribution to this work. This study was supported by grants from Tianjin Basic Research and Leading Edge Science Project of China (to Jian Tan) (No. 08JCZDJC23900).
Disclosure Statement
This article is our own work; we have no specific disclaimer or conflicts of interest.