
Abstract
Polo-like kinase 1 (PLK1) showing a high expression in various kinds of tumors is considered a candidate target for cancer therapy. The aim of our study was to explore the effects of silencing PLK1 gene on human colorectal carcinoma cell line HCT-116 in vitro and in vivo. In vitro, the plasmids generating short hairpin RNA (shRNA)-targeting PLK1 were transfected into HCT-116 by using FugeneHD reagent, and the silencing potency was measured by RT-PCR, western blot, flow cytometry, and Caspase-Glo 3/7 assay, respectively. In vivo, the growth inhibition capacity of PLK1-shRNA on HCT-116 xenograft was measured in nude mice. Then, the silencing effect of PLK1 was analyzed by RT-PCR, western blot, and immunohistochemistry, respectively. Apoptosis, angiogenesis, and proliferation in tumor tissues were measured by TUNEL, CD31, and PCNA stain, respectively. The RNA interference targeting PLK1 significantly decreased the expression of PLK1 in vitro. More importantly, anti-PLK1 treatment in HCT-116 xenograft decreased tumor weight by 81.58% compared with the control group (p<0.001), accompanied with decreased PLK1 mRNA and protein expression, increased cell apoptosis, and reduced angiogenesis and proliferation (p<0.001). Our study showed that knockdown of PLK1 by shRNA might be the potential therapeutic approach against human colorectal carcinoma.
Introduction
Polo-like kinase 1 (PLK1) is the most thoroughly studied member of the polo family that is the conserved serine/threonine kinase. 1,2 It has an amino-terminal kinase domain and a carboxy-terminal polo box domain (PBD). The N-terminal kinase domain of PLK1 incorporates most of the invariant residues that are the hallmarks of serine/threonine protein kinases. This region of PLK1 includes the consensus motif, Gly-X-Gly-X-X-Ala. The C-terminal region of PLK1 encompasses two polo-boxes and the PBD that contains a phosphopeptide-binding motif. The PBD could help PLK1 target to specific subcellular locations. This localization could then lead to recognition by post-translational modification pathways, the nucleation of multiprotein signaling complexes, and control of the conformation, activity, and substrate specificity of particular enzymes for regulating the kinase activity of PLK1. 3 The PLK1 has been shown to play critical physiological functions in ensuring a smooth and error-free progression through mitosis, as a key regulator of cell division in eukaryotic cells. The PLK1 expression and activity normally appear at the onset of mitosis and fluctuate in a cycle-dependent manner. They play very important roles in the G2/M transition, including initiation of mitosis by activating Cdc25C and cyclin B/cdc2 complex, promoting the centrosome maturation, driving microtubule nucleation, regulating spindle assembly checkpoint, activating the components of the anaphase-promoting complex for mitotic exit, and facilitating cytokinesis. 4 –6 Lu et al. generated PLK1 knockout mice and found that PLK1 homozygous null mice were embryonic lethal, and early PLK1−/− embryos failed to survive after the eight-cell stage. Immunocytochemistry studies revealed that PLK1-null embryos were arrested outside the mitotic phase, suggesting that PLK1 was important for proper cell cycle progression. 7 In addition, PLK1 also participates in many kinds of activities in the cells by interaction with many other molecules. For example, PLK1 binds to the tumor suppressor p53 in mammalian cultured cells, inhibits transactivation activity of p53 as well as its pro-apoptotic function, and decreases the p53-mediated transcription from the p53-responsive p21WAF1, MDM2, Apaf-l, BAX promoters. 8 The p63 and p73 are actually the oldest evolutionary conserved members of the p53 family, seemed as the tumor suppressor; they could also be phosphorylated by PLK1 and then lose their transcriptional as well as pro-apoptotic activity. 9,10 The PLK1 expression varies among different types of tissues. Normally, PLK1 is expressed at very high levels in the issues with active cell proliferation, such as the placenta, spleen, and testis, and at very low levels or even undetectable in most other adult tissues, including the liver, kidney, brain, thymus, intestine, lung, pancreas, heart, stomach, and skin. 11 Overexpression of PLK1 could let cells cross those mitotic checkpoints without proper chromosome alignment and segregation; it would engender the immature cell division, the generation of aneuploidy, and tumorigenesis. 12,13 Based on various analyses of human tumors and tumor cells lines, PLK1 has been found overexpressed in the breast cancer, colorectal cancer and endometrial carcinoma, esophageal carcinoma, gliomas, malignant melanoma, nonmelanoma skin cancer, nonsmall cell lung cancer, ovarian cancer, pancreatic cancer, prostate cancer, and so on, and its overexpression was usually associated with tumorigenesis and poor outcome. 14 –24 Recent studies indicate that PLK1 has an effect on motility, migration, and invasive ability of cancer cells. The PLK1 has been found as a novel marker of metastases and the mediator of invasion via vimentin and beta1 integrin. 25,26 These researches indicate an important role of overexpression of PLK1 in tumor.
Nowadays, many literatures describe the function of PLK1 in molecule level and its antitumor effect in vitro. 6,27,28 In our study, to observe the feasibility and effectiveness of knockdown PLK1 as gene therapeutic target for colorectal carcinoma both in vitro and in vivo, the HCT-116 cell line was chosen and the PLK1 expressions of the cell line were diminished by RNA interference (RNAi).
Materials and Methods
Cell culture
Human colorectal carcinoma cell line HCT-116 was obtained from ATCC; cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 U/mL streptomycin (all from Invitrogen); and maintained in a humidified atmosphere of 5% CO2 at 37°C.
The short hairpin RNA expressing plasmids construction and the cell transfection
All the short hairpin RNA (shRNA) expressing plasmids in our study were designed and constructed by our lab. The PLK1-shRNA contained a region specific to target 1412–1430 bp of human PLK1 mRNA (5′-CCT TGA TGA AGA AGA TCA C-3′). The control sequence, named HK, had no homology to any mammalian sequence (5′-GAC TTC ATA AGG CGC ATG C-3′). The recombinant plasmids were named pshPLK1 and pshHK, respectively.
The shRNA expressing plasmids were transfected into HCT-116 cells by FugeneHD (Roche) according to the manufacturer's instructions. Approximately 2×105 HCT-116 cells were plated per well in six-well plates. After 24 hours, 2 μg plasmids (pshPLK1 or pshHK) and 5 μL FugeneHD were diluted in 250 μL media without serum, respectively, and then combined together at the final volume of 500 μL at room temperature for 20 minutes. The plasmid/liposome complexes were added into the cell cultures. The cell that added only FugeneHD 5 μL was considered a blank control. At 48 hours post-transfection, the effects in all the cells were observed, and the cells were harvested for RT-PCR, western blot, flow cytometry assay, and Caspase-Glo 3/7 assay.
Semiquantitative RT-PCR
The total RNA of the HCT-116 cells and tumor tissues was extracted using the TRIzol reagent (Invitrogen), according to the manufacturer's protocol. All the RNA concentration was measured by UV absorbance spectroscopy. The RT-PCR was performed with 1 μg of the isolated total RNA in a final volume of 25 μL by using one-step RT-PCR kit (TaKaRa). The GAPDH was used as the internal control. Primers specific for human PLK1 was designed as follows: PLK1-F: 5-CGAGTTCTTTACTTCTGGCTAT-3, PLK1-R: 5-GGAGACTCAGGCGGTATGT-3. After incubation at 55°C for 30 minutes, the reverse transcription reaction was terminated by heating at 94°C for 2 minutes. The PLK1 and GAPDH cDNA were amplified by 27 cycles of denaturation for 0.5 minute at 94°C, annealing for 50 seconds at 55°C, and extension for 45 seconds at 72°C. Finally, 72°C extended 10 minutes. The PCR products were checked by electrophoresis on a 1.5% agarose gel, imaged by Quantity One (Bio-Rad).
Western bolt analysis
For the western blot analysis, about 1×106 transfected cells were lysed in 100 μL of RIPA lysis buffer (50 mmol/L Tris, 150 mmol/L NaCl, 1% Triton, 0.5% deoxycholate, and 2 mmol/L EDTA) supplemented with protease inhibitor cocktail (Sigma-Aldrich). The xenografts from each group were lysed after the tissues pulverized in liquid nitrogen and crushed by the ultrasonic wave. The total protein was quantified by the Bradford method. Total protein (about 100 μg) was separated on 10% polyacrylamide gel by electrophoresis and then transferred (at 100 V for 45 minutes) to the polyvinylidene difluoride membrane (Bio-Rad) according to the protocol. Membranes were washed in TBS once, and then, nonspecific binding sites were blocked by 5% nonfat milk for 1 hour at room temperature. Membranes were incubated with primary antibodies overnight in TBST with 5% powdered nonfat milk at 4°C. Monoclonal mouse antibodies were used against PLK1 (1:1000; Upstate Biotechnology), and monoclonal mouse antibodies were used against GAPDH (1:20,000; Sigma). The membranes were washed thrice by TBST and then incubated for 1 hour in 5% powdered nonfat milk with peroxidase-conjugated second antibody (1:5000, goat anti-mouse; ZSGB-Bio ) at 37°C for 1 hour. The membranes, washed thrice by TBST and once by TBS, were added to the reagent for enhanced chemiluminescence (Thermo scientific) and detected by X-ray film.
Flow cytometry assay
Transfected cells were harvested, washed twice by 4°C phosphate-buffered saline, then collected, and expanded for double staining using Annexin V-FITC Apoptosis Detection Kit (KeyGEN) to detect cell apoptosis, or PI (50 μg/μL) staining to test cell cycle, according to the manufacturer's protocol. Then, cells were measured on an FACScan flow cytometer analysis, and the data were analyzed by using Cell Quest software (both from Becton Dickinson).
Caspase-Glo 3/7 assay
The cell apoptosis caused by RNAi was assessed by using the Caspase-Glo® assay (Promega) following the manufacturer's directions. The HCT-116 cells were plated in an opaque 96-well plate (Sigma) at 5000 cells per well in complete media. After 24 hours, all the cells were transfected with 0.2 μg plasmid DNA (pshPLK1 or pshHK), and the cells added to the FugeneHD alone were considered as blank control. After 48 hours, 100 μL Caspase-Glo 3/7 reagents were then applied to each well, and the reagent, vehicle, and cell culture medium without cells were treated as background. A luminometer (Applied Biosystems) was used to quantitate luminescence.
Human colorectal cancer xenograft model and treatment
All the animal studies were approved by the Institutional Animal Care and Treatment Committee of Sichuan University (Chengdu, Sichuan, The People's Republic of China). Female nude mice (BALB/c, 5–6 weeks of age) were fed in the standard microisolator caging system, which was free of pathogens. Human colorectal carcinoma cancer xenograft model was established by injecting 1×106 HCT-116 cells subcutaneously into the right flank of the mice. When the tumors reached the volume of nearly 80 mm3, the mice were stochastically divided into three groups (5 mice per group): 5% glucose (GS treatment), pshHK treatment, and pshPLK1 treatment. The acquisition of the cationic liposome was from Avanti Polar Lipids (Alabaster), and the cholesterol was bought from Sigma. The cationic liposome was prepared in our lab using the procedure previously described. 29 Five (5) μg Plasmid DNA and 25 μg cationic liposome were complexed gently in 100 μL of 5% glucose, then the complex was injected into the mouse tail vein thrice a week for totally ten times. Tumor diameters were measured every 2 days during the treatment period. The mean tumor volume was calculated according to the formula: volume=length×(width2)/2. One day after the last treatment, animals were sacrificed and tumors were excised and weighed. Some tumor specimens in each group were fixed in 4% formaldehyde, embedded in paraffin for immunohistochemical analysis, meanwhile some were stored in liquid nitrogen immediately for RT-PCR and western blot, and others were embedded in optimal cutting temperature freezing compound for immunohistochemistry-frozen analysis. The treatment-related toxicity was mainly estimated by general status of nude mice, including the significant weight lose, cachexia, toxic death, ruffling of fur, strange behavior, and so on. To detect possible damage in the process of tumor therapy with RNAi, the tissues of heart, liver, spleen, lung, and kidney were stained and sequentially detected by microscopic examination.
Immunohistochemistry
The expression level of PLK1, PCNA, and CD31 were measured by immunohistochemistry on 3 μm paraffin section and 8 μm frozen sections, respectively. The primary antibodies were mouse anti-human PLK1 (1:1000; Upstate Biotechnology); mouse anti-human PCNA (1:500; Santa Cruz Biotechnology); and rat anti-mouse CD31 (1:250; BD Pharmingen) that were immunohistochemistry frozen. The Ultrasensitive S-P (mouse) KIT-9701 (Maixin-Bio) and nonbiotin system (rat) pv-6004 (ZSGB-Bio) applied the blocking reagent, 3% H2O2, and bound antibody. Then, for color development, a DAB system (ZSGB-Bio) was used. Immunohistochemical analysis was performed according to the procedure of the manufacturer's protocol. To calculate the percentage of PLK1 and PCNA-positive cells, five high-power fields (400×) were randomly chosen and analyzed, and the vessels of each slide were scanned and counted at middle power magnification (200×). All specimens were observed by the Olympus BX600 microscope with a Spot Fiex camera. To certificate no specific staining, the secondary antibody alone was added to serve as the negative control.
In situ TUNEL staining
DeadEnd™ Fluorometric TUNEL System (Promega) was used to evaluate the apoptotic cells of the tumor paraffin section in situ, according to the manufacturer's instructions. From each tumor tissue section, the numbers of green fluorescence-positive cells were counted at high-power fields (200×) in five random fields.
Statistical analysis
The data were presented as the mean±SD. For comparison of individual time points, differences between groups were tested by performing one-way analysis of variance (ANOVA, S-N-K) using the Statistical Package for the Social Sciences (Statistic 17.0). A value of p<0.05 was considered significant, and all p-values were two sided.
Results
Downregulation of PLK1 expression in HCT-116 cells by RNAi targeting PLK1 in vitro
The effect of pshPLK1 transfection on the silencing of PLK1 gene expression was evaluated by using both RT-PCR and western blot analysis. As shown in Figure 1A and 1B, at 48 hours after transfection, the expression of mRNA and protein level were, respectively, decreased by PLK1 targeting RNAi in HCT-116 cells; whereas the expression was insignificantly affected by pshHK, comparing with blank control.

Suppression of PLK1 expression by shRNA transfection at the mRNA and protein level in human colorectal carcinoma HCT-116 in vitro.
Downregulation of PLK1 expression inhibited cell growth and increased apoptosis in vitro
To validate the effects of HCT-116 cell growth and cell apoptosis after the inhibition of PLK1 expression, a series of experiments in vitro was done. First, HCT-116 cells were observed at 48 hours after transfection. The cells with pshPLK1- transfected showed growth inhibition, shrunken and floating, quite different with the blank control and pshHK-transfected, as shown in Figure 2A. Next, Caspase-Glo 3/7 assay was used to measure the cell apoptosis accurately. In this assay, the luminescence value with the subtracted background of each group was evaluated. We supposed the blank control (FugeneHD alone) as base, compared with it, the Caspase 3/7 activity of other groups expressed as relative times (mean%±SD): 1.87±0.17 (pshHK) versus 3.26±0.05 (pshPLK1), as shown in Figure 2B. The pshPLK1-treated cells showed a significant increase in cell apoptosis comparing the pshHK-treated cells (p<0.001), and the experiment was repeated thrice with similar results. Then, the apoptosis of each group was examined by flow cytometry. As shown in Figure 2C, 8 hours after transfection, the apoptotic proportion of blank control, pshHK group, and pshPLK1 group were 7.2%, 11.0%, and 31.7%, respectively. We also found that the pshPLK1 group induced the G2/M arrest on comparing the pshHK-treated group with the blank control as shown in Figure 2D. The proportion of G1 was 63.7% (blank group), 68.4% (pshHK group), and 32.5% (pshPLK1 group); and the proportion of G2 was 11.1% (blank group), 16.0% (pshHK group), and 35.9% (pshPLK1 group), respectively. These results indicated that PLK1 suppression was correlated to the cell growth inhibition, increased cell apoptosis, and cell cycle arrest.
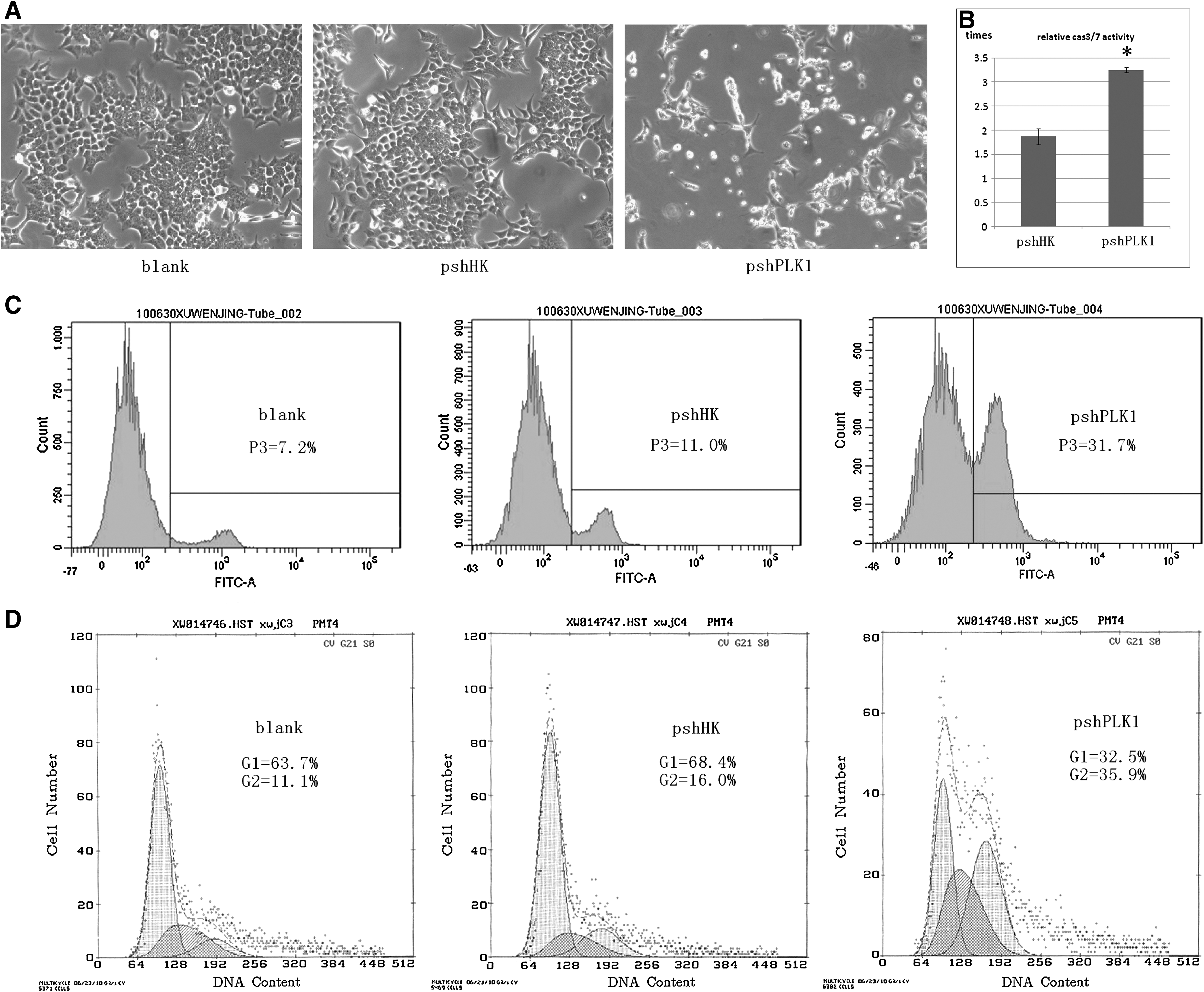
Effect of shRNA transfection on cell growth and apoptosis in human colorectal carcinoma HCT-116 in vitro.
Antitumor effect of RNAi targeting PLK1 in vivo
When the palpable tumor developed and reached nearly 80 mm3 at the site of injection, mice bearing tumors were treated as described earlier. According to Figure 3A and B, it could be seen that anti-PLK1 RNAi caused a conspicuous suppression of tumor growth. Comparing with the GS group, the pshPLK1 treatment effectively slowed down the tumor growth by 75.92% (p<0.001). The mean tumor weight was 0.76±0.08 g, 0.62±0.06 g, and 0.14±0.01 g in GS group, pshHK group, and pshPLK1 group, respectively. Comparing with the GS group, the pshPLK1 treatment reduced the tumor weight by 81.58% (p<0.001). Since pshPLK1 caused significant tumor growth inhibition and tumor weight reduction comparing with pshHK, it suggested that the antitumor effects were produced by PLK1 targeting RNAi rather than by nonspecific reactions. It was observed that the pshHK treatment also showed the tumor inhibition effect comparing with GS treatment, the result might be explained as misinterpreting the therapeutic effects of RNAi caused by immune stimulation as described. 30,31
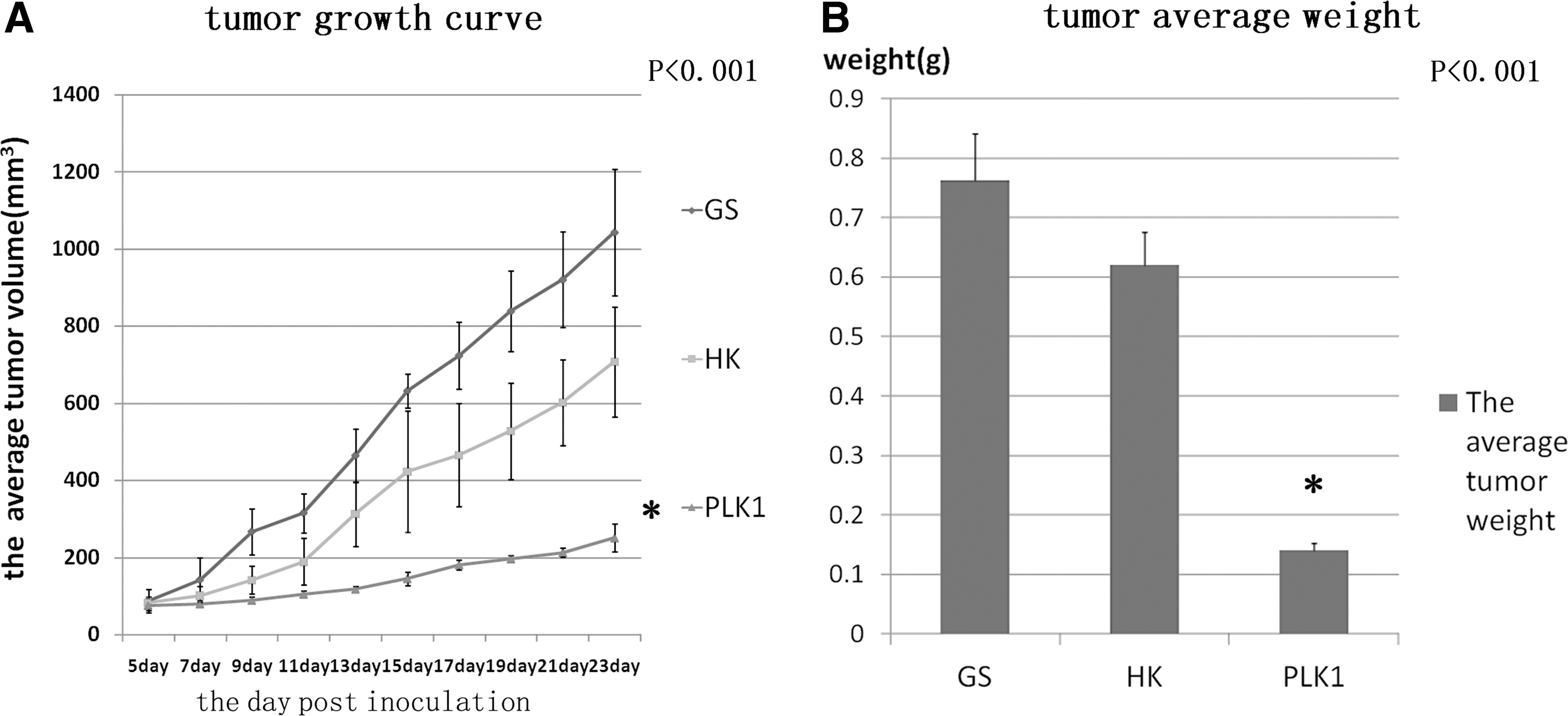
Anti-cancer effect of RNAi targeting PLK1 in HCT-116 xenografts.
Downregulation of PLK1 expressions by RNAi targeting PLK1 in vivo
To confirm that the therapeutic effects were attributed to decreased PLK1 by shRNA knockdown, the RT-PCR was used to measure the mRNA expression level, and immunohistochemistry and western blot were used to measure PLK1 expression in protein level in the tumor tissues. As shown in Figure 4A, we found the mRNA level notably decreased by RNAi targeting PLK1, compared with other groups. We detected the downregulation of PLK1 expressions in protein level (Fig. 4B) and observed a general decrease of PLK1 staining in the tumor tissue of pshPLK1 treatment, whereas the tumors treated with pshHK group and GS group exhibited significantly more PLK1 staining (Fig. 4C). The average number of PLK1-positive cells was counted, respectively, to be 412.80±32.03 (GS group), 309.60±20.28 (pshHK group), and 70.40±7.09 (pshPLK1 group), as shown in Figure 4D (p<0.001).
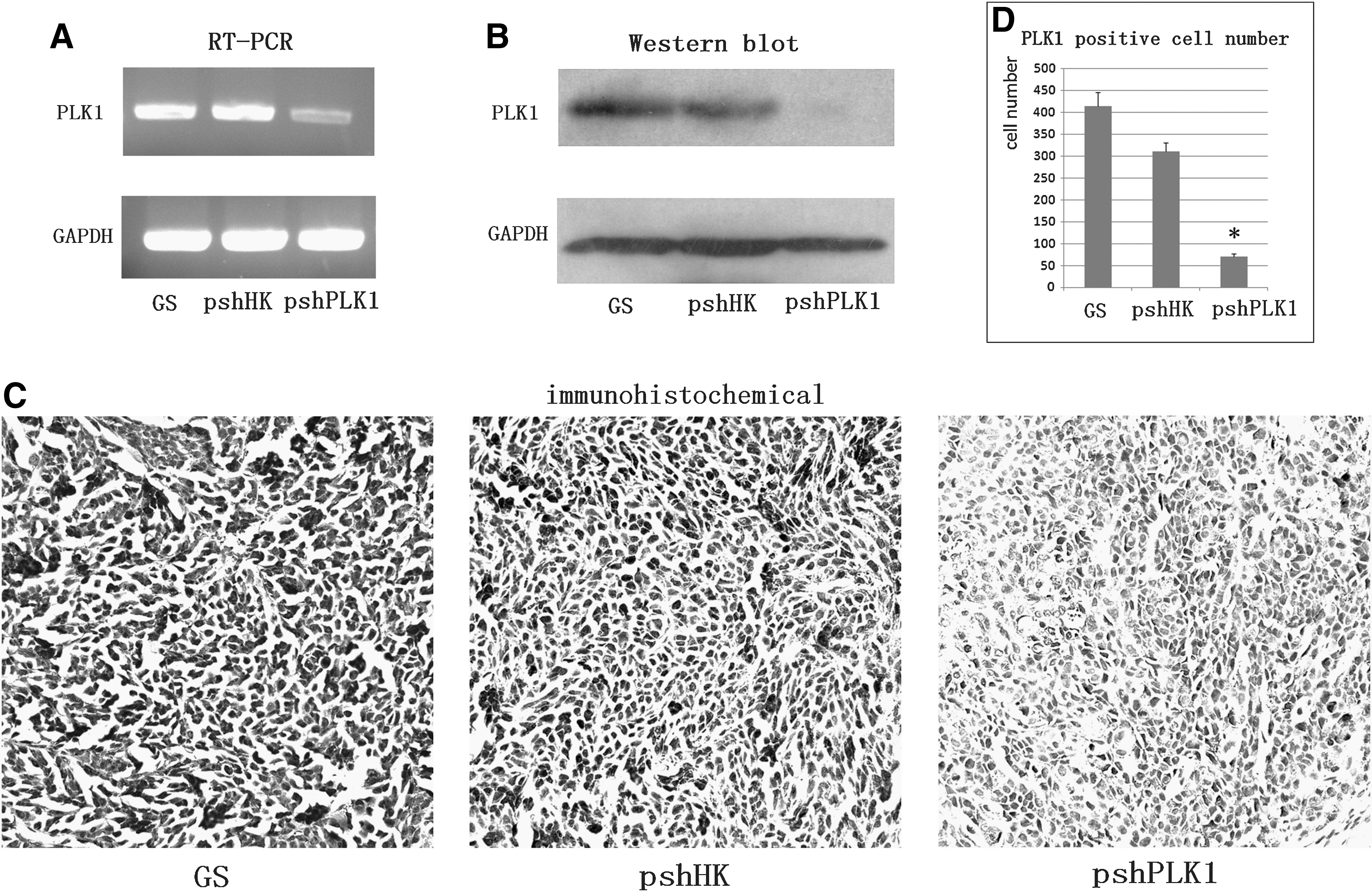
shRNA targeting PLK1 decreased the PLK1 expression in vivo.
The effect of RNAi targeting PLK1 on cell apoptosis, angiogenesis, and proliferation
We measured the tumor cell apoptosis by in situ TUNEL staining. The apoptosis cells were counted and analyzed in five random fields, and the tumor cell apoptosis was augmented with the treatment of pshPLK1 when compared with GS or pshHK, (88.00±15.68 versus 11.00±3.39 or 16.80±2.59, p<0.001), as shown in Figure 5A. Then, the mean microvessel density (MVD) was judged by counting the vessel of CD31 staining: the mean MVD number was counted, respectively, to be 83.40±7.06 (GS group), 51.00±3.16 (pshHK group), and 15.60±2.30 (pshPLK1 group). The significant reduction in MVD occurred in the tumors of the mice receiving the pshPLK1 treatment in comparison with pshHK and GS groups (p<0.001) in Figure 5B. The PCNA staining was used to evaluate the tumor cell proliferation. As shown in Figure 5C, the PCNA-positive staining cells were obviously reduced in the tumors of the mice treated with pshPLK1, the data showed that the PCNA-positive cells were 411.20±21.60, 404.20±43.46, and 149.20±18.67 in GS group, pshHK group, and pshPLK1 group, respectively (p<0.001), and there was no significant difference between GS group and pshHK group. All the positive cells were counted and analyzed in five random fields.
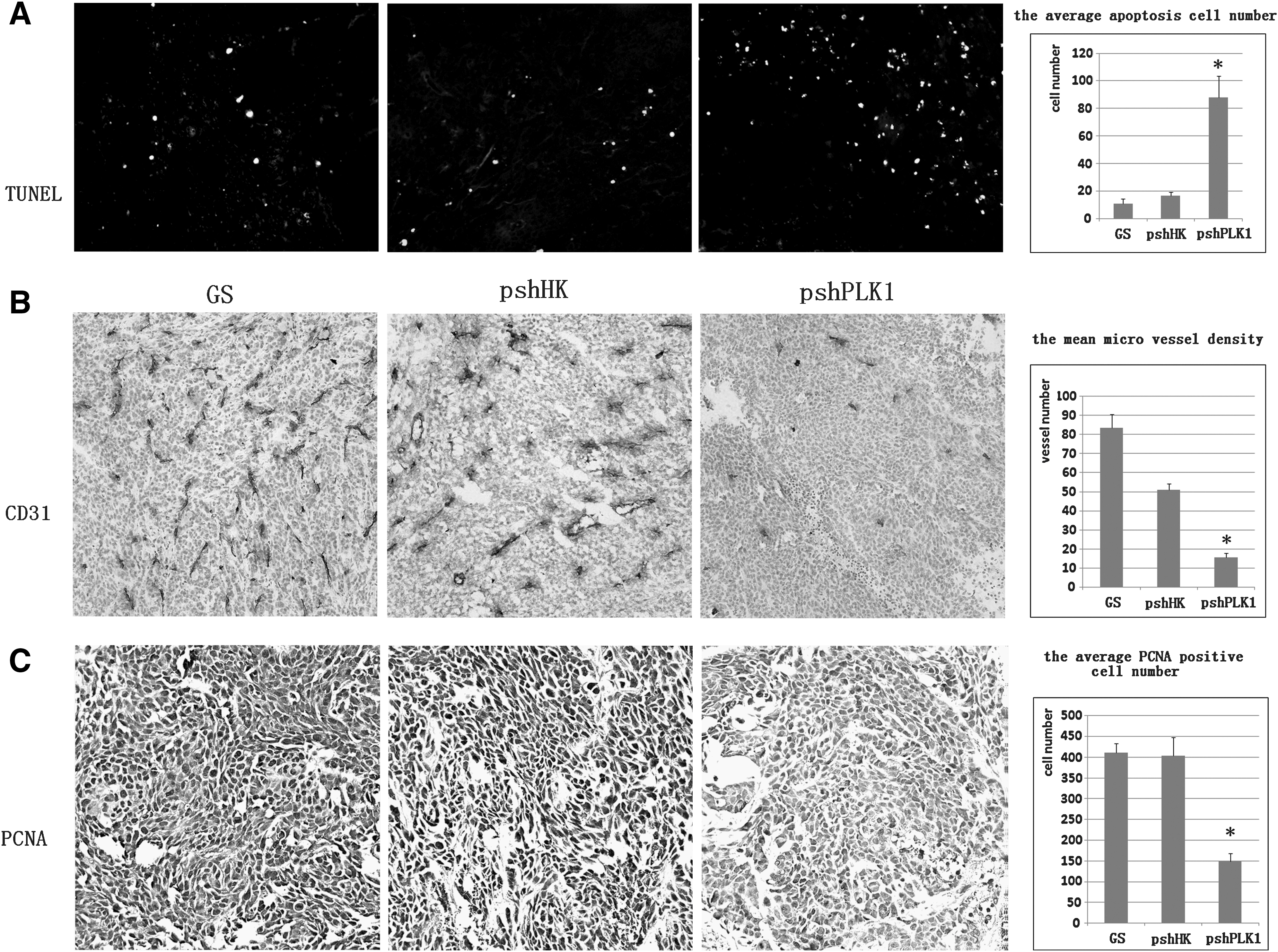
Effect of RNAi targeting PLK1 on cell apoptosis, angiogenesis, and proliferation in vivo.
At the end of the therapy, according to our daily observation and the evidence obtained by tissue slices of the main organs, no obvious toxicity was found. No mice showed toxic death, strange behavior, ruffled fur, significant weight loss, or other consequences of therapy-related toxicity. Microscopic and macroscopic examination of tissue section revealed no pathologic indication of toxicity-associated damage (data not shown).
Discussion
Colorectal carcinoma is the most common malignancy and it ranks fourth in frequency in men and third in women. It has become more and more a severe health problem all over the world. 32 Surgical oncology plays an important role in the treatment; the chemotherapy, radiation therapy and molecular targeting therapy are good auxiliary treatments. Although colorectal carcinoma has quite a good prognosis, mortality rate is still about one half that of incidence. 32 The patient alive with unresectable colorectal cancer still has a poor quality of life. Since people have already realized the limitation of any single traditional therapy, a new method is needed to enrich the treatment tools and to improve the curative effect. With the molecular biology and genetic engineering development, gene therapy has become the hot spot. Gene therapy is relatively effective, and it has a high potential for clinical application. Although gene therapy might not be the first choice for the anticancer treatment, its development provides a new choice for nonsurgical treatment to improve patient outcome.
Nowadays, more and more genes have been found to play a critical role in the progress of tumorigenesis, 33 and the mitotic cancer genes are considered as new insights into tumor cell biology research and cancer therapy. 34 Regular cell cycle progression, especially mitotic progression, maintains the genomic stability. The studies of preneoplastic specimens of humans have demonstrated that DNA damage markers exist in the precancerosis. The failed DNA repair causes chromosomal instability, and then DNA damage propagates to daughter cells by mitosis, which seemed to be the primary cause of carcinogenesis. 35,36 The mitotic progression is mainly promoted by cyclin-dependent kinases that are controlled by several critical kinases including PLK1. Recent studies have shown downregulation of PLK1 could induce tumor cell apoptosis and improve the sensitivity of antitumor drug, whereas normal cells are rarely influenced. 37 From the researches nowadays, therapy targeting PLK1 achieves a good effect in many tumor models in vitro and in vivo. 3,38 There are also many investigations about PLK1 in the colorectal cancer model. Kaiser et al. 39 reported that compared with normal tissue of adult colon, all mouse and human tumors overexpressed a massive cluster of genes including PLK1, which were highly enriched for functions related to cell-cycle progression, proliferation, and migration. The PLK1 is also found overexpressed at both the transcriptional and protein level in rectal cancer specimens. Overexpression of PLK1 is associated with poor outcome and treatment resistance in cancer. Takahashi et al. found that progression of primary colorectal cancer was associated with increased expression level of PLK1, and they speculated that overexpression of PLK1 might be of pathogenic, prognostic, and proliferative importance. Apart from the potential diagnostic value, modulation of PLK1 activity in tumors by chemotherapeutic agents or gene therapy might prove to be of clinical value. 15 Since tumor cells that overexpressed PLK1 exhibit a more radioresistant phenotype and the irradiation could influence PLK1 expression of cells in a time- and dose-dependent manner, PLK1 might be a novel predictive marker for radiation response as well as a promising therapeutic target in rectal cancer patients. 40 Recently, the effects and the molecular mechanism of PLK1 have been well studied, and the anticancer effect has been commendably verified in vitro. Fan et al. 41 demonstrated that knockdown expression of PLK1 using antisense oligonucleotides in SW480 cells induced apoptosis and caused arrest of cells in the G2/M phase of cell cycle, but the practical potency in animal model is still poorly known. In our study, we demonstrated that the delivery of plasmids DNA encoding for anti-PLK1 shRNA could downregulate PLK1 expression in the human colorectal cancer model both in vitro and in vivo. Based on the results of RT-PCR and western blot, the specific shRNA targeting, PLK1 was considered to successfully reduce the expression of PLK1, whereas nonspecific shRNA caused no effect on PLK1 expression, indicating that shRNA could be used as an effective specific inhibitor of PLK1. More importantly, in HCT-116 xenografts experiments in nude mice, tumor growth was obviously inhibited in the anti-PLK1-treated group comparing with the pshHK group or GS group, providing a proof of the true antitumor activity of PLK1-shRNA. In our study, the preliminary anti-tumor mechanism was also studied. Our data showed that transfection of HCT-116 cells with pshPLK1 resulted in the G2/M arrest, tumor cells apoptosis and increased Caspase 3/7 activity in vitro, as well as the increased tumor cell apoptosis and the decreased cell proliferation in vivo. Further, MVD in anti-PLK1-treated group was significantly lower than that in GS group and pshHK group. Our results suggested that cell-cycle arrest, increased apoptosis, and reduced proliferation might be the mechanism of antitumor effect of pshPLK1. It demonstrated the effectiveness of anti-PLK1 therapy once again, providing the evidence for clinical application.
There are many optional ways to inhibit PLK1, and some molecular inhibitors are currently evaluated in preclinical and clinical trials. 42 Considering their poor target selectivity, these inhibitors should be substituted by more specific and more potent therapy such as RNAi. It is a new form of antisense gene therapy with an expression plasmid encoded for the shRNA that was composed of a stem-loop structure. This shRNA is processed in the cell to an RNA duplex, and this short RNA duplex mediates post-transcriptional gene silencing by RNAi-induced silencing complex (RISC) or RISC-like complexes. 43 Several approaches for delivering RNA in vivo are selectable. Virus-mediated delivery has been reported to be effective, but the safety of viral vectors is a problem. 44 Therefore, a nonviral gene delivery system has been chosen. Liposome-mediated RNAi is usually considered effective to deliver drugs with low toxicity unless administered at very high doses. The advances in synthetic small interfering (siRNA) delivery provide solutions to many of the challenges in siRNA therapeutics, and give RNAi-based gene silencing technologies more comprehensive application for clinical trial. 45 Nowadays, RNAi has been considered a vigorous tool for anticancer therapy. We got an apparent effect by using RNAi to reduce PLK1 expression of the tumor cells in vitro and in vivo. According to the previous study in our lab, 29 intravenous administration of plasmids were given every 2 days at a dose of 5 μg per mouse, and the strictly executed dose/scheduled strategy attained good treatment effectiveness without severe toxicities.
There are still several limitations to discuss in our study. First, we consider that the subcutaneous implantation is not the ideal microenvironment for colon cancer cells and, hence, can influence the delivery of drugs to the tumor. In fact, we thought that the better animal model for research setting would be the spontaneous model, as the tumor tumorigenesis, the microenvironment, and the response to the tumor therapy are similar to the clinical setting. Recently, genetic modification could successfully build up the tumor models. These animal models are usually useful for the research on etiology, genetics, and immunology of tumors. However, these models also have the following shortcomings: the model would require a long term and large expenditures to be established; the incidence is hard to control; and so on. The spontaneous tumor model still need a quite long time for large-scale application. However, our study focused on the therapeutic research, and our aim was to establish the animal model of human colon tumor for gene therapy. Thus, the spontaneous tumor model or the genetic model was not very appropriate for our study. On the other hand, the subcutaneous implantation model was widely used in therapeutic research, because of its advantages: stable, easy to observe and easy to build, and easy to control. Therefore, we chose the subcutaneous tumor model for our study. Second, we found that pshPLK1 treatment had obvious effectiveness than pshHK or blank/GS treatment in vitro and in vivo. However, at the same time, pshHK treatment, compared with blank/GS treatment, also had a certain effect of promoting apoptosis. In recent studies, in vitro cytotoxicity of cationic liposome and its complex with pDNA (lipoplex) was observed and investigated regardless of the pDNA type, cell lines, or the concentration of serum. The cytotoxicity induced by lipofection could largely increase, reduce, or even mask the desired effects of gene-based therapy. 46 In recent years, a phenomenon named “off-targeting” has been demonstrated by a lot of investigations. Some investigations have shown that siRNAs with no prospective physiological target sequences, which are often used as negative controls, might have specific off-target effects that induced “toxic phenotype.” 47 The siRNA-induced “toxicity” described in the study is sequence dependent and target independent. These could be the reasons that the HK treatment increased apoptosis, whereas the PLK1 expression was not obviously affected in vitro experiments. The circumstances are more complex in vivo. The mammalian innate immune system could be stimulated by canonical siRNA duplexes, and then excessive cytokines (such as TNF-α, interleukin-6, and interferons (IFNs), particularly IFN-α) would be released. Activation of innate immunity could evidently inhibit tumor growth, angiogenesis, and inflammatory and other immunological processes. 48 So, these studies demonstrate how important it is to design and deliver shRNA, and it would prompt us to pay attention to its application in future studies.
Conclusions
In summary, our study showed that the RNAi targeting PLK1 was fairly effective in inhibiting tumor growth in vitro and in vivo without overt toxicity. The enhanced antitumor efficacy may be attributed to decreased angiogenesis and increased induction of apoptosis. Our studies suggested its potential clinical application in the treatment of colorectal carcinoma.
Footnotes
Acknowledgments
This work was supported by the National Key Basic Research Program (973 Program) of The People's Republic of China (2010CB529900) and the Hi-Tech Research and Development Program (863 Program) of The People's Republic of China (2007AA021008) and New Drugs Research and Development Importance Special Program (2009ZX09102-241).
Disclosure Statement
No competing financial interests exist.