
Abstract
Human leukocyte antigen G (HLA-G) is an immunotolerant nonclassical major histocompatibility complex Class Ib molecule. It is expressed by trophoblastic placental cells during pregnancy to protect the fetus from maternal alloreactivity. HLA-G is overexpressed in tumors and involved in cancer immune evasion. Reverse transcription–polymerase chain reaction and immunohistochemistry (IHC) were used to examine HLA-G expression in normal mammary and breast cancer cell lines and normal and human breast cancer tissues. Reverse transcription–polymerase chain reaction confirmed that normal epithelial MCF-12A cells had no HLA-G mRNA expression, whereas cancer cell lines MCF-7, T47D, and MDA-MB-231 and NCI/Adr-Res had various levels of HLA-G mRNA expression. Twelve (12) normal and 38 breast cancer tissues were examined by IHC. Fifty-eight (58) percent (22/38) of cancers had medium to strong staining to HLA-G, whereas only 8% (1/12) of normal breast tissues had medium to strong staining, and the difference was significant (p < 0.05). HLA-G staining was found in the membranes and cytoplasm of cancer cells. In conclusion, breast cancer cells overexpress HLA-G mRNA and protein, and this probably contributes to immune evasion.
Introduction
Human leukocyte antigen G (HLA-G) is a nonclassical major histocompatibility complex (MHC) Class Ib molecule with immunomodulatory and immunotolerant properties. HLA-G is normally selectively expressed in extravillous (intermediate) trophoblastic cells of the placenta at the maternal–fetal interface. 1 Since that discovery, HLA-G expression in human tumors has been demonstrated, which supports the view that HLA-G may be involved in tumorogenesis by promoting immunosuppression in the tumor microenvironment. 2
The first evidence that functional HLA-G protein expression may play a role in tumor escape from the host immune system was reported by Paul et al. in 1998. 3 They showed that HLA-G expression protected melanoma cells from NK-mediated cell cytolysis. Others 2,4 have shown that HLA-G tremendously affects almost every aspect of human innate and adaptive immunity. Lin et al. 5 have shown that HLA-G expression in human ovarian carcinoma inhibits NK cell function. HLA-G inhibits the cytolytic function of peripheral NK cells, 6,7 the antigen-specific cytolytic function of cytotoxic T-lymphocytes, 8 the alloproliferative response of CD4 + T-cells, 9,10 the proliferation of T and NK cells, 11,12 and the maturation and function of dendritic cells (DC). 13,14 Carosella et al. 15 have elegantly discussed all of the above HLA-G functions and have investigated many increasing complexities of the HLA-G immunomodulatory molecule.
Uroservic and Dummer 16 have done an excellent job discussing the role of HLA-G in cancer immunoediting. They suggested that HLA-G was associated with tumor growth and progression, as it was preferentially detected in tumor tissue and only rarely in adjacent normal tissue. The HLA-G role in all phases of immunoediting, namely (1) elimination, (2) equilibrium, and (3) escape, were addressed in remarkable detail. Their paper has contributed tremendously to understanding the role of HLA-G in human malignancy. Ye et al. 17 have shown that HLA-G expression is a prognostic indicator in patients with colorectal cancer; Kleinberg et al. 18 demonstrated the expression of HLA-G in malignant mesothelioma and clinically aggressive breast carcinoma.
In the present study, the expression of HLA-G in human breast cancer cell lines and breast cancer tissue was evaluated. It was confirmed that there was an upregulation of the nonclassic MHC Class Ib HLA-G immunomodulatory molecule in breast cancer tissue.
This article is not intended to be a complete review or a large research study on HLA-G. The goal of the present study was to bring more awareness to the role of HLA-G in cancer immunosuppression and stimulate more research in this area.
Materials and Methods
Reverse transcription–polymerase chain reaction (RT-PCR) was used to evaluate the expression HLA-G in the normal breast epithelial MCF-12A cell line, the breast cancer cell lines MCF-7, T47D, and MDA-MB-231, and a drug-resistant cell line NCI/Adr-Res. Immunohistochemistry (IHC) for HLA-G was performed on 12 normal breast tissue specimens obtained from breast reduction cases and on 38 breast cancer specimens obtained at the time of mastectomy. The percentage of cell staining was noted and the staining intensity was graded 0, 1 + , 2 + , and 3 + , with 3 + being the most intense. The acquisition of paraffin tissues was approved by the authors' in-house IRB.
Immunohistocytochemistry
Twelve (12) normal breast tissue specimens from plastic surgery cases and 38 breast cancer specimens obtained at time of mastectomy were examined. All paraffin blocks were requested from pathologists. All diagnoses were confirmed by pathology. Five (5)-millimeter-thick paraffin sections were cut. The sections were deparaffinized using xylene, rehydrated through a graded series of ethanol, and rinsed in phosphate-buffered saline. The sections were submitted to antigen retrieval with 0.1% trypsin for 15 minutes at 37°C and rinsed in phosphate-buffered saline for 10 minutes. Histostain-Plus Kits (Zymed Laboratories, San Francisco, CA) were used to detect HLA-G staining. In brief, nonspecific antibody binding was prevented though serum blocking solution. The sections were incubated for 60 minutes at 37°C with a primary monoclonal antibody against human denatured HLA-G heavy chain (Abcam, Cambridge, MA). After washing, the sections were covered with biotinylated second antibody and incubated at room temperature for 10 minutes. The sections were washed again and incubated with streptavidin peroxidase conjugate at room temperature for 10 minutes. Slides were washed and covered with DAB chromogen for 5 minutes at room temperature. Then, the slides were washed with water and counterstained with Mayer's hematoxylin and mounted. All sections were examined by light microscopy.
RT-PCR
Total RNA from the normal and all cancer cell lines was purified by RNA microisolation kit (Gibco Invitrogen, Carlsbad, CA). One (1) microgram of RNA was used as the template for RT. The RT reaction mixture contained 0.5 mM dNTP, 0.1 nm Oligo(dT)12–18, and 0.25 units AMV reverse transcriptase (Promega, Madison, WI) and was incubated at 42°C for 60 minutes. The PCR was performed in a total of 25 μL with 2 μm forward and reverse primers, 5 mM MgCl2, 20 μM each of dGTP, dATP, dCTP, and dTTP, and 0.125 unit Taq DNA polymerase. HLA-G–specific RT-PCR amplifications were carried out as described using G.257 (exon 2) and G3.U (3′-UT) primers to detect all alternatively spliced HLA-G mRNA forms. 19,20 Primers used were as follows: forward primer: 5′-GGA AGA GGA GAC ACG GAA CA-3′; reverse primer: 5′-TGA GAC AGA GAC GGA GAC AT-3′. mRNA levels of the β-actin gene were used as an internal control of gene expression. The primers for amplification of β-actin gene were 5′-CTTCTACAATGAGCTGCGTG-3′ and 5′-GAGGATCTTCATGAGGTAGTC-3′. The amplification procedure involved denaturation at 94°C for 30 seconds, annealing at 56°C for 45 seconds, and extension at 72°C for 90 seconds in an Omn-E DNA Thermal Cycler (National Labnet Co., Woodbridge, NJ). After 35 PCR cycles, there was a final elongation step at 72°C for 10 minutes. The PCR products were run on a 2% agarose gel and visualized by ethidium bromide fluorescence. The sizes of PCR products were determined by comparison with standard PCR DNA markers.
Results
RT-PCR
The normal breast epithelial cell line MCF-12A had no HLA-G expression by RT-PCR, whereas the breast cancer cell lines MCF-7, T-47D, and MDA-MB-231 and the drug-resistant cell line NCI/Adr-Res had various levels of HLA-G mRNA expression. There were three mRNA isoforms expressed in the breast cancer cell lines (Fig. 1)
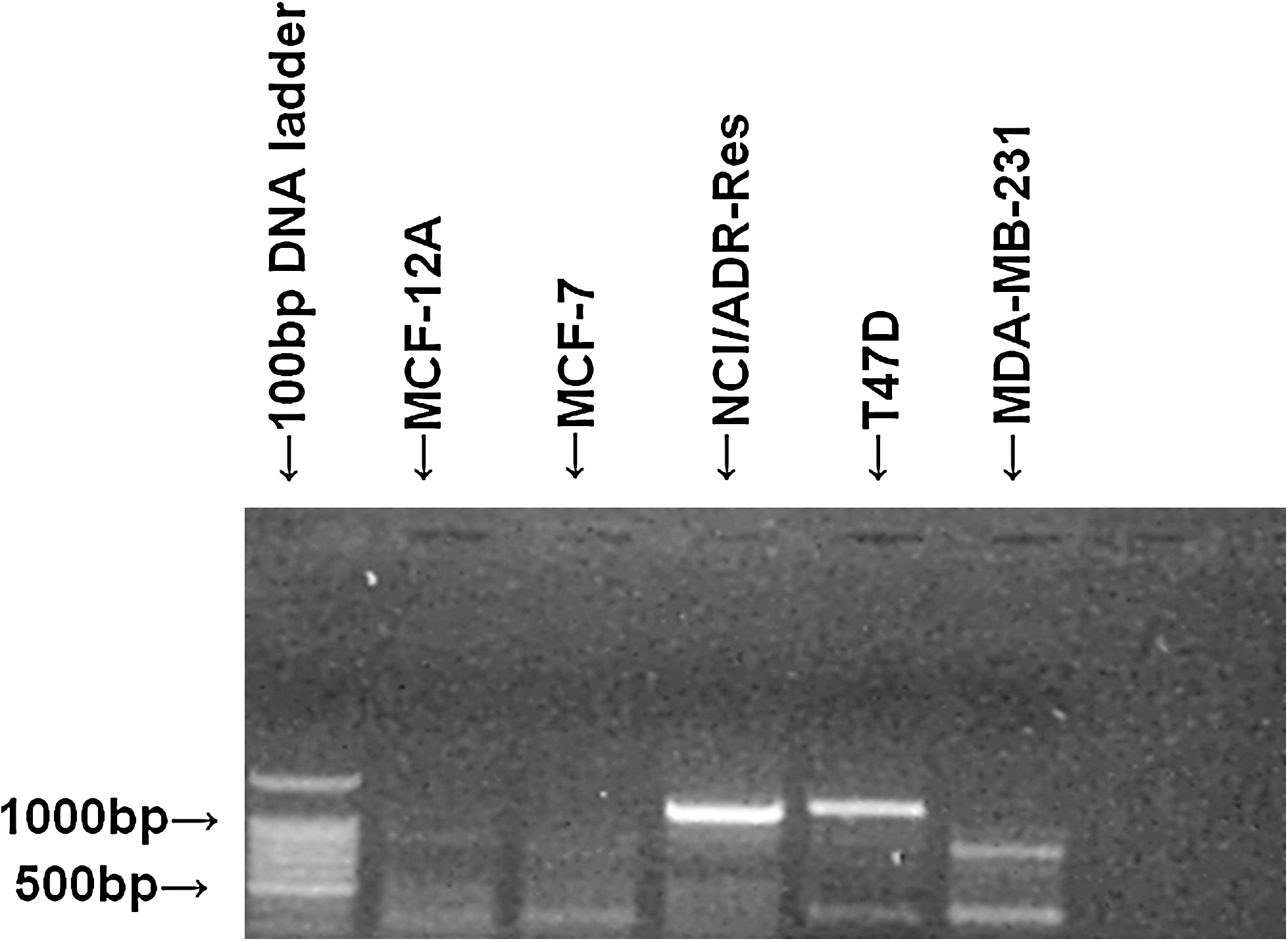
Expression of HLA-G mRNA (reverse transcription–polymerase chain reaction). HLA-G, human leukocyte antigen G.
Immunohistocytochemistry
Twelve (12) normal breast tissues from plastic surgery cases and 38 breast cancer tissue from mastectomy specimens were examined by IHC. Fifty-eight (58) percent (22 of 38 samples) of breast cancer samples had medium to strong staining (2 + or 3 + staining) for HLA-G, whereas only 8% (1 of 12 samples) of normal breast tissues had medium to strong staining (Fig. 2A, B, respectively ), and the difference was significant (p < 0.05, Fisher's exact test). The staining of HLA-G was noted to be on the cell membranes and was diffuse in the cytoplasm of the cancer cells and some normal mammary epithelium. There was a rare, weak HLA-G staining in fibroblast, vascular endothelium, and some inflammatory cells.
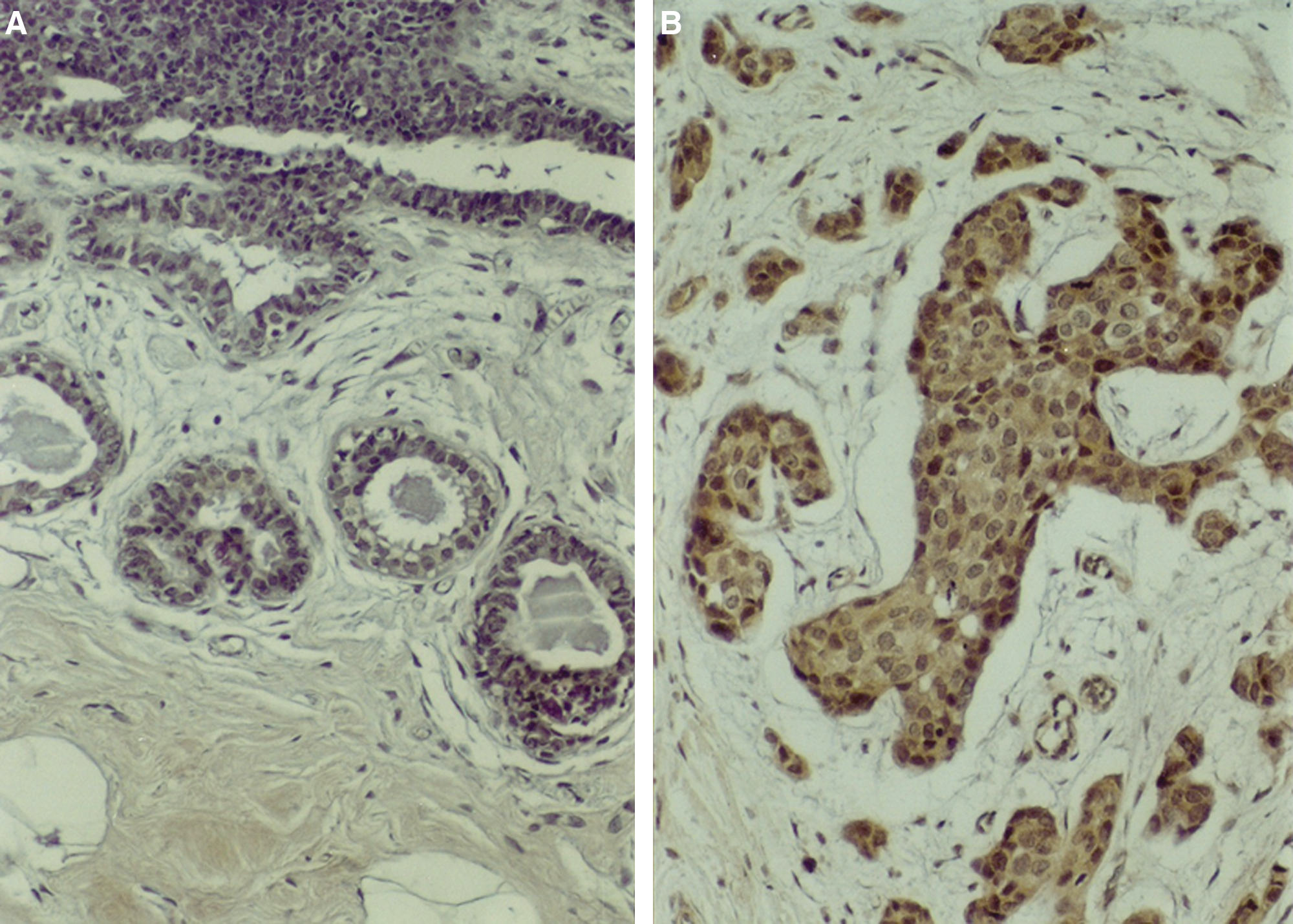
Photomicrograph of a normal breast specimen with no HLA-G staining
The results confirm the presence of HLA-G mRNA and protein in breast cancer cell lines and that tumor specimens stain much more intense for HLA-G by IHC than normal breast tissue.
Discussion
This study, as others, 5,17,18 confirmed that the nonclassical MHC Class Ib molecule HLA-G is preferentially expressed in malignant breast tissue and poorly expressed in normal breast tissue. There was no correlation of HLA-G expression and tumor biology, such as estrogen receptor, progesterone receptor, Her-2-neu status, tumor grade, or node status, in the randomly selected breast cancer cases examined.
Cancer immunosuppression and tumor escape mechanisms are extremely numerous, complex, confusing, and, at times, paradoxical. The evidence is clear that HLA-G is involved in many aspects of tumor immune evasion 5 –8,11 –14 ; however, the exact role of HLA-G in tumor immune suppression is yet to be elucidated. Many of the players in tumor immune evasion are well known (Box 1), and some of their individual functions in cancer immune evasion have been elucidated. However, many questions remain unanswered, creating the so-called (1) “which comes first the chicken or the egg syndrome”; (2) is there a main primary initiator (father) of cancer immune evasion, or is it just a heterogenous random event that once initiated causes a cascade of many immunosuppressive actions? It makes sense that, although complicated with many entities involved, cancer immunosuppression is an extremely well-organized cascade of molecular events. The evidence suggests that HLA-G may play a central role in initiating cancer evasion, as HLA-G serves as a nature's way of protecting the fetus from maternal allorecognition.
Immunosuppressive Mechanisms in Tumor Microenvironment
(1) Fox P3 T-regulatory cells (T-regs)
(2) Myeloid-derived suppressor cells (MDSC)
(3) Tumor-associated macrophages (TAMs)
(4) Tumor-associated fibroblast (TAFs)
(5) Mast cells (very paradoxical)
(6) STAT3-NF-kB interactions (STATs) are important proteins central in determining whether immune responses in tumor microenvironment inhibits or promotes cancer growth
(7) HLA-G (a natural main initiator of immunosuppression)
(8) IL-6
(9) IL-1B
(10) IL-10
(11) IL-17
(12) IL-23
(13) VEGF
(14) MMP2
(15) MMP9
(16) CCL21
(17) CTLA-4
(18) TGF-B
(19) Arginase
(20) Indolamine 2,3-dixoygenase (IDO)
(21) Nitric oxide (NO) and reactive oxygen species (ROS)
(22) Th17 T-cells
(23) Prostaglandins E2—cyclooxyenase-2 (COX-2)
(24) Toll-like receptor actions (TLR4 and TLR9)
Many are paradoxical and depend on interaction with each other.
Liang et al. 21 have shown that the modulation of DC differentiation by HLA-G and ILT4 requires the IL-6 STAT3 signaling pathway. Their results suggested that different isoforms of HLA-G have distinct roles in suppression of maturation and activation of DC through the ILT4 receptor. They concluded that there was an HLA-G strategy used by different isoforms of HLA-G to downregulate DC activation and function. Rajagopalan et al. 22 have shown that there is a monomeric HLA-G5 that binds an activating receptor, KIR2DL4, on human resting NK cells and initiates the expression of a group of cytokines and chemokines that drive a proinflammatory and angiogenic response.
Yu et al. 23 have published a tremendous review on the role of STATs in cancer inflammation and immunity and discussed in detail the role of STAT3. They point out that “STAT proteins are central in determining whether immune responses in the tumor microenvironment promote or inhibit cancers.” They noted that persistently activated STAT3 increases tumor cell proliferation, invasion, and survival, while supporting tumor immunosuppression. They suggest that STAT3 might be a promising target for cancer therapy. However, throughout their excellent review, which discussed the complexity of cancer immune evasion, the possible role of HLA-G was not once mentioned, and of 157 references there was none concerning HLA-G.
The fact that there was no mention of HLA-G in the above review is somewhat baffling, but not surprising. It is no surprising because at the 101st meeting of the AACR there were many sessions on tumor immunology, tumor microenvironment, tumor escape mechanisms, cancer vaccines, and immunotherapy and there was not one comment about HLA-G and its possible role in cancer immunosuppression. Yet, there is much evidence in the literature to suggest that HLA-G plays an important and probably central role in cancer evasion, at least in a large percentage of epithelial malignancies.
Paul et al. 3 studied more than a thousand lesions of malignant melanoma and showed that HLA-G transcription and protein expression could be switched on in tumor lesions to protect them from NK cytolysis. Later, several investigators 24,25 reported that tumor lesions were protected from NK cell cytolysis by HLA-G expression. This also correlates with breast and ovarian carcinomas. 26 Morandi et al. 27 have observed high HLA-G plasma levels in patients with neuroblastoma and that increased levels correlated with relapse.
A number of studies of HLA-G protein expression on epithelial tumors determined by immunohistochemistry confirmed a large percentage of HLA-G protein expression in many different types of carcinomas. 28 –33 There were different results in the various types of malignancies, which might reflect the differences in tumor biology of the different tumor types analyzed or could reflect the sensitivity of the methods used to detect the HLA-G antigens in the malignant lesions.
Tripathi and Agrowal 4 stated that changes take place during the progression of cancer, and they are varied, such as expression of tumor markers and changes favorable for tumor development and escape from immunosurveillance. They suggested that one such molecule with immunosuppressive and immunomodulatory roles in cancer development is HLA-G. Hofmeister and Weiss 34 have explained in detail some mechanisms of HLA-G immunosuppression. They showed that HLA-G antigens play a role in establishing and maintaining immune tolerance by inhibiting the immunocompetent cell functions. These inhibitory effects are initiated by HLA-G binding to its receptors ILT2, ILT4, and KIR2DL4. Therefore, HLA-G through these receptors interacts with NK cells, antigen-presenting cells, and T-lymphocytes and mediates its immunosuppressive and tolerant functions at different stages of the immune response.
Carosella et al. 15 have elegantly reported on the aspects of HLA-G biology and shown the critical HLA-G pathologic relevance that should help design HLA-G–based diagnostics. First is the demonstration of HLA-G multiple structures and that these multimers carry most of the HLA-G inhibitory function. They suggest that the pathologic relevance of HLA-G may have been underestimated because of the lack of a dimer-specific detection method. Second, HLA-G is not just a shield against effector immune attack, but can also exert a long-term inhibitory function through regulatory cells. Third, HLA-G can migrate from cell to cell and induce effector cells to take on regulatory functions. Therefore, the HLA-G inhibitory functions extend beyond the reach of only HLA-G–expressing cells. All of the above data confirm the extreme importance of even a few HLA-G–expressing cells.
It seems reasonable that HLA-G should be evaluated in at least all epithelial malignancies and in all HLA-G–positive patients; HLA-G should constitute a therapeutic target. It could also be used as a tumor marker, and the blocking of HLA-G could delete its effect as a contributor to tumor escape and tumor immunosuppression. The present study suggests that HLA-G should receive more research and attention as a very important molecule involved in cancer immune evasion and progression. HLA-G should be considered as a target in most cancer immunotherapeutic protocols.
Footnotes
Acknowledgments
This research was supported by funds from The Sallie Astor Burdine Breast Foundation, Baton Rouge, LA.
Disclosure Statement
None of the authors has any institutional or commercial affiliations that might pose a conflict of interest regarding the publication of this article.