
Abstract
Administration of natural or synthetic agents to inhibit, delay, block, or reverse the initiation and promotional events associated with carcinogenesis opens a new avenue for cancer prevention and treatment to reduce cancer morbidity and mortality. Eugenol, a potential chemopreventive agent, is a component of clove and several other spices such as basil, cinnamon, and bay leaves. A number of reports have shown that eugenol possesses antiseptic, analgesic, antibacterial, and anticancer properties. The present study was undertaken to evaluate the chemopreventive potential of eugenol alone and in combination with a chemotherapeutic agent such as gemcitabine. Eugenol showed dose-dependent selective cytotoxicity toward HeLa cells in comparison to normal cells, pointing to its safe cytotoxicity profile. A combination of eugenol and gemcitabine induced growth inhibition and apoptosis at lower concentrations, compared with the individual drugs. The analysis of the data using a combination index showed combination index values of <1 indicating strong synergistic interaction. The combination thus may enhance the efficacy of gemcitabine at lower doses and minimize the toxicity on normal cells. In addition, the expression analysis of genes involved in apoptosis and inflammation revealed significant downregulation of Bcl-2, COX-2, and IL-1β on treatment with eugenol. Thus, the results suggest that eugenol exerts its anticancer activities via apoptosis induction and anti-inflammatory properties and also provide the first evidence demonstrating synergism between eugenol and gemcitabine, which may enhance the therapeutic index of prevention and/or treatment of cervical cancer.
Introduction
Current treatment modalities for cancer are effective to combat the transformed cells, but these treatments result in bystander effect, which typically refers to the death, altered growth, or damage of cells that are not the direct target of chemotherapy or irradiation. 1,2 Little progress has been made in the treatment of cervical cancer because of the absence of effective systemic therapies. It is therefore of primary interest to search for new therapeutic agents able to contrast this aggressive tumor without accompanying side-effects on neighboring normal cells, to reduce cancer morbidity and mortality.
In the struggle against cancer, novel therapeutic approaches are constantly being sought to complement existing therapeutic agents. One such potential approach is the use of chemopreventive agents, natural or synthetic noncytotoxic nutrients and/or pharmacological agents, during the prolonged time period between tumor progression and frank malignancy, to inhibit, delay, block, or reverse the initiation and developmental events associated with carcinogenesis. Further, these agents should possess relatively safe toxicity profiles and may act as prospective chemo- and radio-sensitizers. 3,4
A previous study in the authors' lab demonstrated the chemopreventive potential of clove. 5 Eugenol is one of the main components in the essential oil extracted from cloves, comprising 72%–90% of the total, and is also found in reasonable quantities in the essential oils of various other spices such as bay leaves and cinnamon leaf. Various scientific reports have proved that eugenol possesses antiseptic, antibacterial, analgesic, antiviral, apoptosis-inducing, cytotoxic, antioxidant, antiangiogenic, antiproliferative, anti-inflammatory, and anti-invasive properties. 6 –11
Eugenol has been proved to be noncarcinogenic in long-term carcinogenicity experiments by various groups, and other studies point toward its tumor-inhibiting properties. 12,13 In an interesting study by Ghosh and coworkers, eugenol resulted in a significant tumor growth delay and decrease in tumor size. 14 Administration of eugenol has been found to significantly reduce the incidence of MNNG-induced gastric tumors. 15 Eugenol downregulates various molecular targets, that is, it suppresses NF-κB activation, inhibits prostaglandin biosynthesis, blocks COX-2 activity, arrests the cells in the S phase of the cell cycle, increases ROS generation, reduces Bcl-2 level, inhibits inflammatory cytokine expression, and results in subsequent apoptotic cell death. 10,14 –18
Further, chemopreventive agents might be used singly or in combination with chemotherapy or radiotherapy for the more effective treatment of cancer by enhancing the efficacy of these modalities with minimal side-effects and toxicity. 19,20 Considering that eugenol may be a prospective chemopreventive agent, its potent antitumor ability to interfere with cervical cancer cell growth and its molecular mechanism were evaluated as an initiative toward the development of a novel strategy for cancer treatment. Also, the combination of eugenol with gemcitabine, a chemotherapeutic agent, was investigated to potentiate the efficacy of the latter at lower doses and thereby reduce its side-effects on normal cells.
Materials and Methods
Cell culture
The human cervical carcinoma cell line, HeLa was maintained in DMEM (Sigma) supplemented with 10% fetal bovine serum (Sigma). The cell line was grown in a humidified atmosphere of 5% CO2 in air at 37°C. Lymphocytes were isolated from healthy nonsmoking donors using HiSep Media (HiMedia) as described previously. 5
Preparation of drug solutions
Eugenol (purity 99%) was procured from Sigma, and a stock solution of 100 mM was prepared in ethanol. Further dilutions were made in complete medium to required concentrations between 10 μM and 1 mM for the treatment of HeLa cells and lymphocytes.
A stock solution of 133 mM (40 mg/mL) of gemcitabine (Intas Biopharmaceuticals) was prepared in DMSO (Sigma) and further dilutions were made in complete medium at concentrations of 15, 25, and 35 mM.
Cell viability assay
The antiproliferative activity of eugenol, alone (10 μM–1 mM) or in combination with gemcitabine (15 mM and 25 mM), on HeLa cells was evaluated using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay, as previously described. 5 Cells were seeded at 24 hours prior to treatment in a 96-well plate at plating densities ∼7000 cells/well (in triplicates) to obtain semiconfluent cultures. This was followed by treatment with varying concentrations of eugenol (alone or in combination with different concentrations of gemcitabine) in complete medium for 24 hours. MTT (Sigma) (final concentration: 0.5 mg/mL) was added to each well at appropriate time and incubated for 2–4 hours at 37°C. The formazan crystals thus formed were dissolved in 100 μL of DMSO (Sigma). The absorbance was read at 570 nm using an Absorbance Microplate Reader (BioTek). Similarly, lymphocytes were also treated with the same concentrations of eugenol (10 μM to 1 mM) for 24 hours. Cell viability was calculated as percentage of control (untreated cells) and averaged from three independent experiments.
Calculation of combination effects of eugenol and gemcitabine
Calculations of combination effects were calculated as previously described and were expressed as a combination index (CI). 21 CI analysis provides qualitative information on the nature of drug interaction, and a CI <, =, or >1 represent synergy, additivity, and antagonism of SFN and eugenol, respectively.
Morphological studies of HeLa cells before and after treatment with eugenol
Morphological changes in HeLa cells elicited by eugenol were studied using a normal inverted microscope (Labomed). The untreated cells and cells treated with EC50 dose of eugenol (500 μM) for 24 hours were used for the morphological studies.
Caspase 3 activity assay
Cell apoptosis assay was performed by measuring caspase-3 levels with a Caspase-3 Colorimetric Assay Kit (Sigma) as per the manufacturer's instructions. Briefly, HeLa cells (∼107) were seeded in a 25-cm2 flask and incubated overnight before being treated with eugenol alone (0, 150, 300, 500, and 750 μM) or in combination with gemcitabine (15 and 25 mM) for 4 hours. Following incubation, the cells were lysed in the lysis buffer and the cell lysates were collected for caspase-3 assay as per the instructions. The optical density of the reaction was spectrophotometrically quantified at a wavelength of 405 nm. The concentration of the pNA released from the substrate by caspase-3 was calculated from the absorbance values at 405 nm or from a calibration curve prepared with defined pNA solutions. Normalized caspase-3 levels were expressed as percentage of control. All data were derived from three independent experiments.
RNA isolation and expression analysis of Bcl-2, COX-2, and IL-1β by reverse transcription–PCR
Total RNA was extracted from eugenol-treated and untreated HeLa cells using GenElute Mammalian Genomic Total RNA Isolation Kit (Sigma) as per the manufacturer's protocol. The extracted RNA samples were then subjected to reverse transcription (RT)–PCR using ProtoScript M-MuLV Taq RT-PCR Kit (New England Biolabs) and gene-specific primers. 22 –24 The PCR cycle was as follows: initial denaturation at 94°C for 5 minutes, followed by 35 amplification cycles (denaturation at 94°C for 30 seconds, annealing Tm [β-actin: 56°C; Bcl-2: 55°C; COX-2: 57°C; IL-1β: 51°C] for 30 seconds, and extension at 72°C for 45 seconds), with final extension at 72°C for 7 minutes. Amplified products were separated on a 2% agarose gel containing ethidium bromide.
Statistical analysis
All data are expressed as mean+SD of at least three experiments. Fisher's exact test was adopted for statistical evaluation of the results. Significant differences were established at p<0.05. All statistical analyses were performed in GraphPad InStat Software.
Results
Eugenol induces selective cytotoxicity in HeLa cells
The effect of eugenol on the viability of HeLa cells was determined by treating the cells with different concentrations of eugenol (10 μM to 1 mM) for 24 hours and evaluating the cell viability using MTT assay. Eugenol inhibited the viability of HeLa cells in a concentration-dependent manner (Fig. 1). After 24 hours of treatment, eugenol caused approximately 50% inhibition of cell growth in HeLa cells at a concentration of 500 μM, compared with the untreated controls.
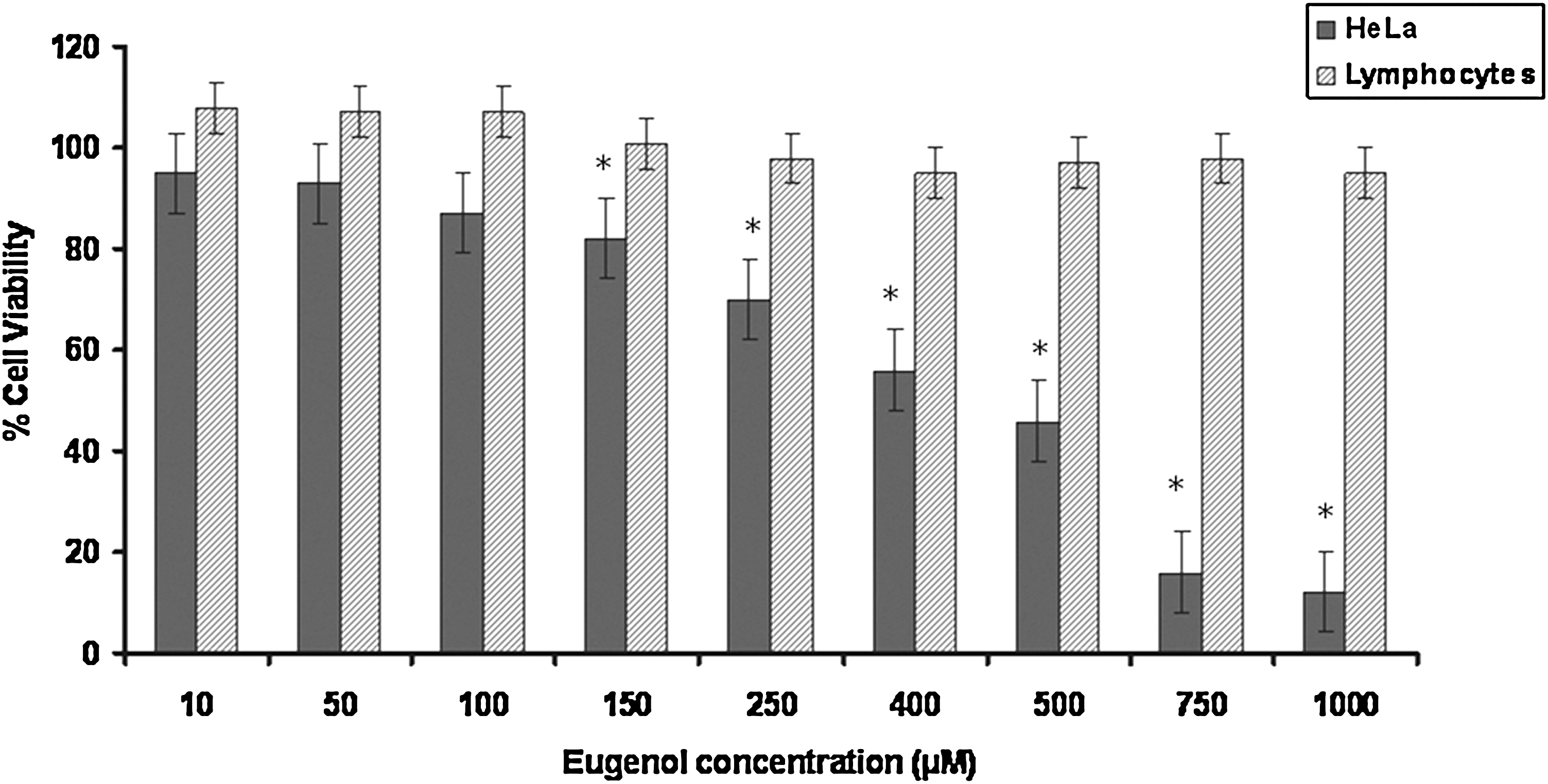
Dose-dependent curve of cell viability of HeLa cells and lymphocytes after treatment with 10 μM to 1 mM eugenol for 24 hours. The LD50 of eugenol on HeLa cells was found to be 500 μM for 24 hours. Values are mean+SD of three independent experiments. *Significant differences between HeLa cells and lymphocytes (p<0.005).
Additionally, the microscopic examination of cells treated with 500 μM eugenol for 24 hours compared with untreated control showed characteristic rounding off of dying cells, indicating that the cell death induced by eugenol is through the apoptotic pathway (Fig. 2A, B).
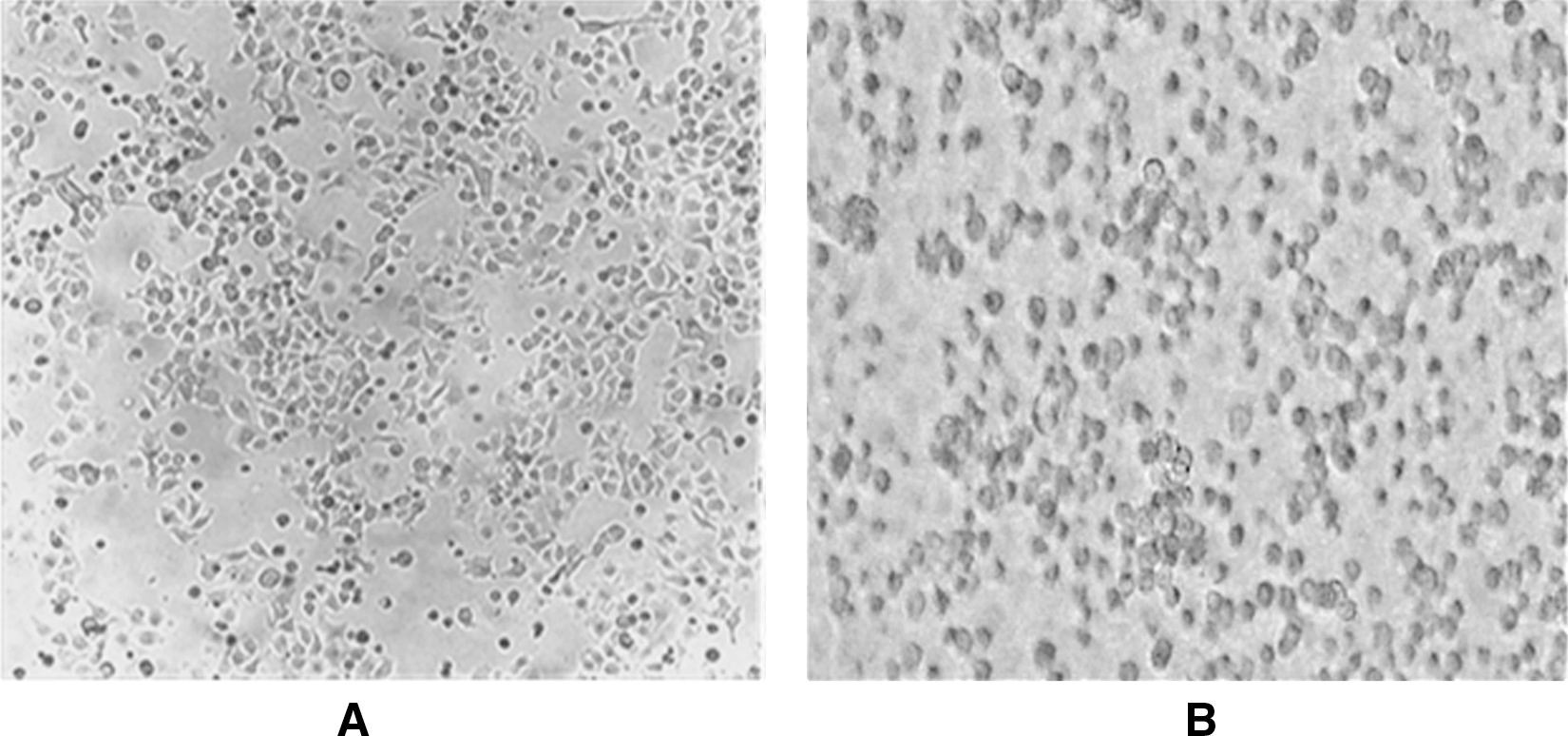
Microscopic features of HeLa cells before treatment
It has been well established that chemopreventive drugs have differential effects on cancer cells and normal cells in vivo and in vitro. 3,4 In light of the aforementioned fact, the effect of eugenol on lymphocytes (as normal cells) was assessed. Isolated lymphocytes from healthy nonsmoker adult were plated in triplicates followed by treatment with eugenol at varying doses ranging from 10 μM to 1 mM for 24 hours (Fig. 1). Treatment of lymphocytes with eugenol for 24 hours showed no significant effect on cell viability, that is, there was no significant cytotoxic effect of eugenol on lymphocytes. This property of eugenol can be utilized in the development modalities for cancer management as it might be safe on normal cells, thereby overcoming the adverse physiological reactions during chemotherapy.
Eugenol synergistically enhances the efficacy of gemcitabine by potentiating its cytotoxic effect on HeLa cells
Common cancer therapies combined with the dietary compounds may exert enhanced antitumor activity through synergic action or compensation of inverse properties, decreasing the systemic toxicity, because lower doses could be used. The present study, performed for the first time, evaluated the efficacy of gemcitabine in combination with eugenol. The combined effects of eugenol and gemcitabine on cell growth inhibition of the HeLa cells are shown in Figure 3. With increasing gemcitabine concentration from 15 to 35 mM, the cell viability decreased from 73% to 51%. Then, the effect of different combinations of gemcitabine with eugenol was investigated. Gemcitabine (15 mM) used in combination with sublethal dose of eugenol (150 μM) resulted in significant decrease in cell viability (47%), compared with either of the compounds alone (73% and 84%, respectively, for gemcitabine and eugenol). Similar results were observed with another combination of 150 μM eugenol with 25 mM gemcitabine resulting in significant decrease in cell viability (39%) in comparison to the individual doses. These results confirm the hypothesis that these drugs when used in combination work in a synergistic manner and enhance the efficiency of gemcitabine at lower doses.

Combination effect of 150 μM eugenol and gemcitabine (15 mM and 25 mM) on HeLa cells for 24 hours. The simultaneous treatment of eugenol and gemcitabine resulted in a synergistic decrease in cell viability, compared with individual drug treatment. Each value is a ratio of the level in the treated cells to that in the control cells. Values are mean+SD of three independent experiments. Each value with eugenol and gemcitabine combination treatment differs from the control value (p<0.05).
Eugenol treatment individually and in combination with gemcitabine induces apoptosis in HeLa cells
The caspase family of cysteine proteases plays a key role in apoptosis and inflammation. Caspase-3 is an important protease that is activated during the early stages of apoptosis and catalyzes specific cleavage of many key cellular proteins. The enzymatic activity of caspase 3 induced by eugenol was assayed using a colorimetric assay kit according to the manufacturer's instructions (Sigma).
Caspase 3 activity was found to increase in a dose-dependent manner (0, 150, 300, 500, and 750 μM) on eugenol treatment in comparison to untreated cells. Interestingly, both the combinations of eugenol and gemcitabine (150 μM eugenol with 15 mM and 25 mM gemcitabine) synergistically augmented caspase 3 activity when compared with the single doses (Fig. 4). These results suggest that eugenol alone and in combination with gemcitabine induce caspase 3–mediated apoptosis in HeLa cells.
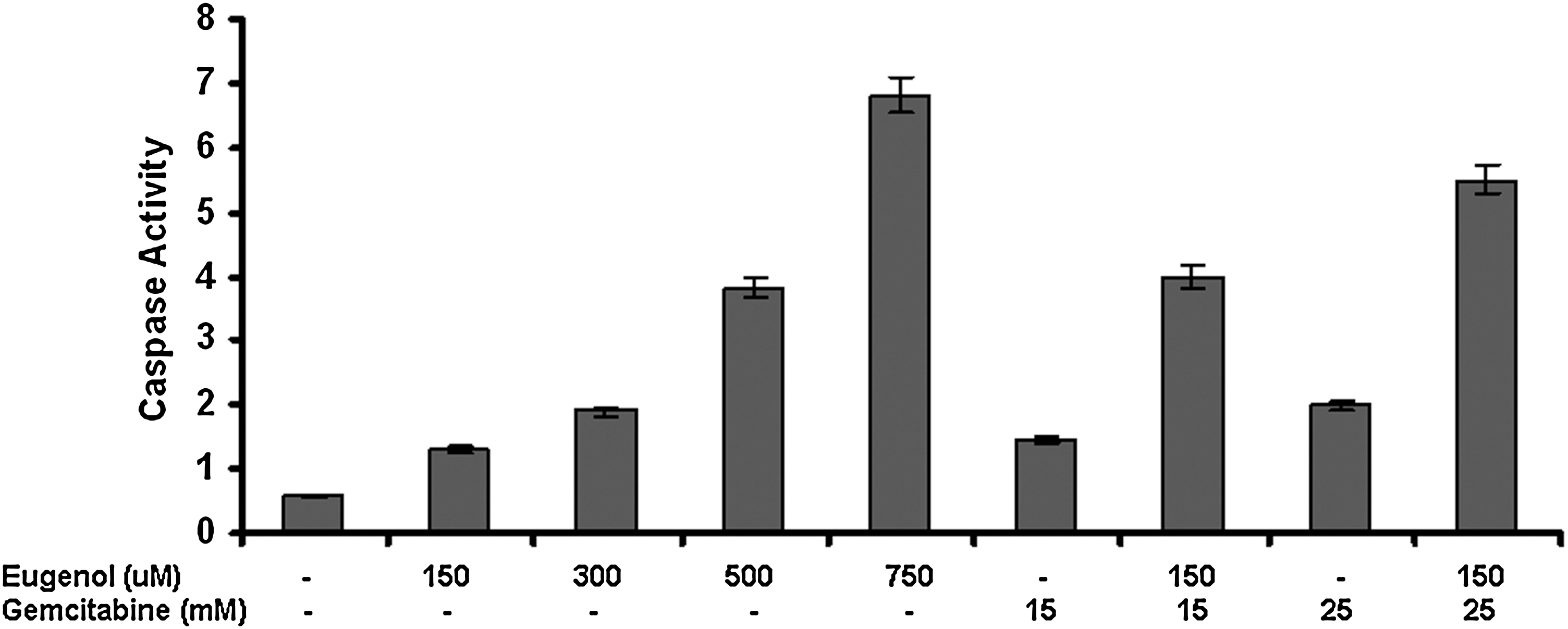
Caspase 3 activity in response to eugenol (0, 150, 300, 500, and 750 μM) and gemcitabine (15 and 25 mM) alone and in combination. The combination treatment showed significantly increased apoptosis in comparison to the drugs singly. Values are mean+SD of three independent experiments. Each value with sulforaphane alone or in combination treatment differs from the control value (p<0.05).
Eugenol significantly downregulates the expression of Bcl-2, COX2, and IL-1β in HeLa cells
To analyze the downstream targets of eugenol in HeLa cells, the expression of Bcl-2, COX2, and IL-1β was evaluated before and after treatment with 300 μM eugenol for 24 hours. β-Actin, which is a widely used house-keeping gene, was used as an internal control for comparison of samples.
Bcl-2 (B-cell lymphoma) is an antiapoptotic gene and has been implicated in a number of cancers, including melanoma, breast, prostate, and lung carcinomas, as well as schizophrenia and autoimmunity. It is also thought to be involved in resistance to conventional cancer treatment. An elevated expression of Bcl-2 was observed in untreated HeLa cells. Its expression was found to be significantly downregulated in eugenol-treated HeLa cells when compared with the untreated control cells (Fig. 5).

Expression analysis of Bcl-2, COX-2, and IL-1β after treatment with 300 μM eugenol on HeLa cells. Eugenol treatment significantly reduced the expression of these genes. Lane U shows untreated HeLa cells; lane T shows eugenol-treated HeLa cells; lane N shows negative control for RT-PCR. β-Actin was used as an internal control. Representative gels from one of the three experiments were used.
Moreover, the effect of eugenol treatment on the expression of COX-2 was analyzed, which has been shown to be involved in inflammation and its upregulation has been reported in various cancers. The untreated HeLa cells showed overexpression of COX-2, which was significantly reduced on eugenol treatment in HeLa cells (Fig. 5).
Proinflammatory cytokines are important in modifying the activity, differentiation, and mitigation of antigen-presenting cells and have been shown to influence the survival of cancer patients. IL-1β, one such proinflammatory cytokine, is involved in proliferation, differentiation, and apoptosis of HeLa cells. The expression of IL-1β gene was analyzed in untreated and eugenol-treated HeLa cells. A high expression of IL-1β was observed in untreated HeLa cells, but after the treatment with eugenol, its expression was significantly abrogated (Fig. 5). These results indicate that Bcl-2, COX-2, and IL-1β are the targets of eugenol and this property can be utilized in the designing of alternate strategies for cancer treatment.
Discussion
Despite advances in the development and improvement of chemotherapeutic drugs, conventional cancer therapy often falls short of the goal of controlling tumor progression and is marred by nonselective nature of these therapies as well as development of resistance. 25,26 Lack of success with targeted monotherapy has forced researchers to utilize novel strategies for cancer treatment such as combination therapy or agents that interfere with multiple cell-signaling pathways. These are considered more promising for higher effectiveness, resulting in improved survival. 27 –30 One such approach is the use of chemopreventive agents that might reduce the cancer risk through various mechanisms and by targeting different stages of carcinogenesis. 27
The use of combinations of low-dose chemotherapeutic and chemopreventive agents may result in achieving a therapeutic synergy between individual drugs, while reducing the toxicity of other agents on normal cells. 31 –33 Recent studies point toward many dietary compounds (i.e., genistein, 3, 3′-diindolylmethane, eugenol, sulforaphane, indole-3-carbinol [I3C], curcumin, (–)-epigallocatechin-3-gallate [EGCG], resveratrol, etc.) as potential chemopreventive agents because of their anticarcinogenic activity. 34 –36 In addition, these compounds also exert the antitumor activities through regulation of distinct cell signaling pathways. 37 Therefore, common anticancer modalities in combination with these chemopreventive agents may exert enhanced antitumor activity through synergism or compensation of inverse properties. Thus, lower doses can be used, resulting in reduction of systemic toxicity caused by chemotherapies or radiotherapies.
Eugenol (4-allyl-2-methoxyphenol) is a main constituent of the essential oil obtained from commonly consumed spices such as Pimenta racemosa (bay leaves), Cinnamomum verum (cinnamon leaf ), and Syzygium aromaticum (clove). 12 In recent times, it has been studied for a variety of promising biological properties and is reported to possess antioxidant, anti-inflammatory, and anticancer activities. 14 –18,38 However, eugenol undergoes a pronounced first-pass effect; in serum, unconjugated eugenol was not detected after an oral dose of 150 mg. 39 In this vein, further in vivo analysis is required to confirm the bioavailability and bio distribution of eugenol.
In the present study, eugenol treatment resulted in a significant dose-dependent growth inhibition of HeLa cells (Fig. 1). This is consistent with other studies that also found a dose-dependent effect of eugenol on various human cancer cells. 38 –41
As previously discussed, chemotherapy not only targets malignant tumors but also rapidly dividing normal cells such as bone marrow, fetal cells, germ cells, hair follicles, etc. 1 Hence, a desirable characteristic property of alternate treatment approaches such as chemoprevention would be to possess differential effects on cancer cells and normal cells. Thus, use of such dietary compounds with relatively safe toxicity profiles would forego the side-effects associated with other cancer treatment modalities. 5 Normal cells (isolated lymphocytes) treated with eugenol at similar concentrations had no significant effect on cell viability (Fig. 1). Various studies have established that chemopreventive agents do not have cytotoxic effect on normal cells even at high dosages. 32 –34 Thus, eugenol was found to be selectively toxic to the cancer cells at the doses studied.
A number of in vitro and in vivo studies have demonstrated that chemopreventive agents could enhance the antitumor activity of chemotherapeutics, thus improving the treatment outcome. 32 –34,42,43 This may be through the regulation of multiple signaling pathways, including Akt, NF-κB, c-Myc, cyclooxygenase-2, apoptosis, and others, suggesting a multitargeted nature of chemopreventive agents. 41 –43
A previous study indicates that gemcitabine is cytotoxic toward both cancer and normal cells. 5 At this juncture, for the first time, the in vitro synergism in proliferation-inhibiting and apoptosis-inducing effects of combined treatment with 15 and 25 mM gemcitabine and 150 μM eugenol on a cervical cancer cell line was demonstrated (Fig. 3). The present study's results are in line with the previous studies that observed a synergistic crosstalk between different therapeutic agents. 19,20,26,27,29 –34 One such study found increased sensitivity of cisplatin-induced cytotoxicity in the presence of eugenol by decreasing the expression of multidrug-resistance protein 2 (MRP2). 44 Also, eugenol in combination with gamma-radiation induced radiosensitization of various tumors by initiation of membrane oxidative damage and intracellular reactive oxygen species (ROS) generation. 42 Thus, combining chemopreventive agents may allow to minimize the adverse effects of chemotherapeutic drugs by reducing their effective concentration, should the agent be developed for human cancer intervention. However, further in-depth mechanistic studies, in vivo animal experiments, and clinical trials are needed to investigate the effects of chemopreventive agents in combination treatment of cancer with conventional cancer therapies.
Apoptosis or programmed cell death is a highly regulated process that involves activation of a cascade of molecular events, leading to cell death that is characterized by plasma membrane blebbing, shrinkage, chromatin condensation, chromosomal DNA fragmentation, and formation of membrane-bound apoptotic bodies that are eventually phagocytozed by neighboring cells. 45 Failing to trigger apoptotic cell death may lead to the development of many serious disorders, such as neural degeneration, AIDS, autoimmune disease, and cancers. 10,46 Therefore, cytotoxicity effect via the induction of apoptosis is considered as a criterion for the identification and development of therapeutic agents that can selectively modulate apoptotic pathways, thus providing an effective approach for the prevention and treatment of many diseases including cancer. 45,46 The microscopic examination of cells treated with eugenol showed characteristic rounding off of dying cells, indicating that the cell death induced by eugenol is through the apoptotic pathway (Fig. 2A, B).
To elucidate the mode of cell death induced by eugenol alone and in combination with gemcitabine in HeLa cells, caspase 3 activity colorimetric assays were performed. Caspase-3 is a member of caspases and plays a key role in mediating apoptosis or programmed cell death. Upon activation, it cleaves a variety of cellular proteins, causing morphological and functional changes to cells undergoing apoptosis. 47 In the present study, it was found that the eugenol treatment alone resulted in the activation of caspase 3 in a concentration–response manner (Fig. 4). Induction of apoptosis by natural phytochemicals in malignant cells validates a promising strategy for cancer chemoprevention. 3,8 –10 Various studies demonstrated that eugenol treatment induced caspase cleavage and activation and showed an apoptotic effect in a time- and dose-dependent manner. 8 –10,38
Previous studies have shown that the cytotoxic effects of gemcitabine occur through induction of programmed cell death (apoptosis). 5,32,48,49 In the present study, it was observed that eugenol in combination with gemcitabine significantly enhanced the caspase 3 activity in a synergistic manner, compared with individual drugs. This is along the lines of another study that suggested that gemcitabine with resveratrol can result in a synergistic increase in apoptotic cell death. 50 These results suggest that chemopreventive agents (alone and in combination) induce apoptosis through a caspase-dependent mechanism, which may contribute to the chemopreventive functions of these agents.
In many human cancers, the antiapoptotic Bcl-2 proteins are overexpressed and found to be inversely related to the apoptotic cell index, resulting in resistance to a wide variety of cell death stimuli including chemotherapeutic drugs, γ and UV-irradiation, and cytokine withdrawal. 43,51 –53 Therefore, finding new cytotoxic agents that are able to decrease Bcl-2 expression or restore the ability of tumor cells to undergo apoptosis are vital. The present study investigated the effect of eugenol on the expression of the antiapoptotic gene, Bcl-2, in HeLa cells. RT-PCR results showed that expression of Bcl-2 was inhibited in response to the eugenol treatment in the HeLa cells (Fig. 5). These results are consistent with other studies that also showed downregulation of Bcl-2 on treatment with eugenol. 8 –10,15,18 Considering that reduction in the level of Bcl-2 increases sensitivity to anticancer drugs, reduction in expression of Bcl-2 can be used to potentiate the antitumor effect of chemotherapeutic drugs.
COX-2, which is induced by mitogens, cytokines, and growth factors, is primarily responsible for prostaglandins production in inflammatory sites. 54 –56 Eberhart and coworkers have shown that COX-2, but not COX-1, mRNA expression is markedly elevated in the tissues of most human colorectal cancers compared with accompanying normal mucosa. 55 Overexpression of COX-2 in colonic epithelial cells was found to promote tumor development by causing resistance to apoptosis and facilitating alterations in cell adhesion properties. 56 It has therefore been suggested that COX-2 is an important target for the chemopreventive effects of these agents. 57,58
In the present study, HeLa cells were shown to express high levels of COX-2, which was subsequently downregulated on treatment with eugenol (Fig. 5). According to a previous research by Subbaramaiah and Dannenberg, COX-2 is overexpressed in practically every premalignant and malignant condition involving the colon, liver, pancreas, breast, lung, bladder, skin, stomach, head and neck, and esophagus. 57 Depending upon the stimulus and the cell type, several transcription factors including AP-1 and NF-κB can stimulate COX-2 transcription. 59 Thus, all the dietary agents that can suppress these transcription factors have the potential of inhibiting COX-2 expression. Several dietary components including eugenol have been shown to suppress COX-2. 7,10,57 –60
IL-1β is a member of the interleukin 1 cytokine family and is an important mediator of the inflammatory response. It is involved in a variety of cellular activities, including stimulation of the immune system, production and release of prostaglandins, pituitary hormones, and collagenases, and cell proliferation, differentiation, and apoptosis. The induction of cyclooxygenase-2 (PTGS2/COX-2) by this cytokine in the central nervous system is found to contribute to inflammatory pain hypersensitivity. 61 Therefore, a significant balance exists between the beneficial and harmful effects of IL-1β, thus rendering it an important therapeutic target in a number of pathological conditions including rheumatoid arthritis, atherosclerosis, and diabetes mellitus type I, inflammatory bowel disease and other autoimmune disorders, Alzheimer's disease, leukemia, and solid tumors. 61
Solid tumors in which IL-1β has been shown to be upregulated include breast, nonsmall cell carcinoma, colon, lung, and head and neck cancers, and melanomas, and patients with IL-1β-producing tumors have generally bad prognoses. 62 –65 Contemplating on these facts, the expression of IL-1β in HeLa cells was studied. It was highly expressed in untreated HeLa cells, but after treatment with 300 μM eugenol, its expression was significantly downregulated (Fig. 5). The present study is consistent with another study that found that resveratrol blocks IL-1β-induced activation of the nuclear transcription factor NF-κB. 66 Also, curcumin was shown to suppress IL-1β-mediated microsomal prostaglandin synthase 1. 67
Based on these studies, it can be hypothesized that eugenol may be targeting IL-1β, which then blocks the nuclear translocation of NF-κB and subsequently abrogates the expression of NF-κB downstream targets, Bcl-2 and COX-2. Thus, this study provides an insight into the development of eugenol as a potential anti-inflammatory and anticarcinogenic therapeutic agent acting through multiple signaling pathways. Further insights into the signaling network and interaction points modulated by eugenol as well as in vivo studies and clinical trials are necessary to provide a rational strategy for novel drug development for the prevention and treatment of cervical cancer.
Footnotes
Acknowledgments
The authors appreciate the support and encouragement provided by Dr. B. Ramjee, Director, and Dr. Firdos Alam Khan, Chairperson, Department of Biotechnology, Manipal University, Dubai. The authors are grateful to Ms. Seena Biju, Department of Management Studies (Statistics), for the help provided in statistical analysis. This work was funded by an Intramural Research Program, Department of Biotechnology, Manipal University, Dubai, United Arab Emirates.
Disclosure Statement
No conflicts of interest exist for any of the authors.