
Abstract
Nontargeted cellular effects by ionizing radiation are well documented. The bystander effect is a nontargeted phenomenon wherein the irradiated cells communicate and induce changes in nonirradiated cells. The nature of the bystander signal and how it impacts unirradiated cells remain to be elucidated. Examination of molecular changes could lead to the identification of molecular pathways underlying the bystander effect. In this study, mitochondrial gene transcriptional changes in bystander cells were monitored to gain insight into the participation of mitochondria in this response. The modulation of mitochondrial gene expression in medium-exchanged bystander cells was determined in human lymphoblast TK6 cells by employing the real-time polymerase chain reaction technology. The examination of the relative expression of mitochondrial genes involved in various metabolic functions indicated that MT-ND1, MT-ND5, and MT-ND6 encoding NADH dehydrogenases were upregulated in directly irradiated cells but repressed in bystander cells. The differences in the expression levels were statistically significant among irradiated and bystander cells. The adenosine triphosphate (ATP) synthases MT-ATP6 and MT-ATP8 were upregulated in both irradiated and bystander cells. These results point to the involvement of mitochondrial gene modulation in directly irradiated and bystander cells and provide evidence that mitochondrial gene expression response is part of a complex stress response operating in radiation-treated cells.
Introduction
For many years it was assumed that the cells must be directly hit by ionizing radiation (IR) in order for any biological effect to be manifested. This assumption is no longer true based on the current knowledge of radiation-induced, so-called nontargeted effects. It is clear that the irradiated cells have the capability to signal other cells in the neighborhood. This phenomenon is known as the bystander effect. 1,2 It is shown that the culture medium from irradiated cells has the ability to induce cell killing of nonirradiated bystander cells. 3,4 Understanding the mechanisms underlying the bystander effects of IR has significant implications for effective radiation therapy and risk assessment. 5
The biological responses in bystander cells include an increase in the frequency of micronuclei formation, sister chromatid exchanges, gene mutations, and chromosomal instability. 1,6 –9 Mutations occur in radiation-induced bystander lymphoblast cells, 10 and mutations in the mitochondrial genome have been found in bystander cells. 11 The expression levels of p53, p21, MDM2, CDC2, cyclin B1, and RAD51 were modulated in bystander cells. 12,13 The gene expression profiling identified increases in expression of mitogen-activated protein kinase (MAPK) target genes in radiation-induced bystander cells. 14 The IR-induced reactive oxygen species (ROS) production is involved in the activation of signaling pathways in bystander cells. 15 The linear energy transfer (LET) of radiation influences bystander responses. Although the existence of bystander effect after high LET radiation, such as α-particles, is very well documented, 16 other studies have argued that low LET X-irradiation is also capable of inducing bystander effect. 17 X-irradiation can induce medium-mediated bystander responses in unirradiated human cells cocultured with irradiated cells in a transwell insert culture dish. 9,17
A number of investigations have attempted to understand the underlying mechanism of bystander effect. The production of ROS, direct cell-to-cell signaling via gap junctional intercellular communication, or soluble factors released into the medium surrounding irradiated cells have been suggested as possible mechanisms for the manifestation of bystander effect. 1,3,13,18 –20 The inhibition of the phosphorylation of extracellular signal-related kinase suppressed the bystander response, indicating the participation of the MAPK signaling in the bystander process. 21 The investigation of the involvement of free radicals in the bystander effect identified cellular membranes and mitochondria as ROS producers. 22 The ROS-scavenging agents such as catechins protect bystander cells from radiation. 23 Cellular targets including mitochondria have been postulated to play a significant role in this process. In the present study, the mitochondrial gene expression in irradiated and bystander cells was examined with a view to take an insight into the subcellular processes involved in the bystander effect.
Mitochondria and IR overlap in a number of features; for instance, both generate harmful ROS, and the radiation can induce cell death through the intermediary of mitochondria. Although gene expression arrays have been used to identify stress-induced responses, all of these studies have mainly taken into consideration the modulation of nuclear genes. Very few studies have investigated the response of mitochondrial genes after IR exposure. The mitochondria are responsible to carryout many cellular processes, including energy production by oxidative phosphorylation (OXPHOS), calcium homeostasis, and apoptosis. 2 Mitochondrial DNA (mtDNA) has the ability to replicate independently of the nuclear DNA and the mitochondria are capable of repairing mtDNA damage. 24 The mitochondrial genome encodes 13 proteins necessary for the OXPHOS process, involved in adenosine triphosphate (ATP) generation. The remaining proteins needed for the OXPHOS process are encoded by the nuclear genome. 25 Recently, the mitochondrial gene expression machinery was inventoried and found to have links to human diseases. 25 The changes in the expression of mitochondrial genes in bystander cells cocultured with irradiated cell-conditioned medium have not been reported. This study was undertaken to investigate whether the signal transmission from irradiated cells to bystander cells has the ability to induce changes in the mitochondrial gene expression levels of unirradiated bystander cells.
Materials and Methods
Cell culture and maintenance
The human lymphoblast cell line TK6 was obtained from Dr. Howard Liber (Colorado State University, Fort Collins, CO). These cells were grown exponentially in suspension in a T75 flask using RPMI 1640 supplemented with 10% fetal bovine serum (Invitrogen, Grand Island, NY), 100 mg/mL streptomycin, and 100 U/mL penicillin. The cell cultures were maintained at a density of 2–5×105 cells/mL in a 37°C incubator with 5% CO2 and 100% humidity.
IR treatment and cell coculture
Irradiation of 3×106 cells was performed with an RAD Source 2000 X-ray biological irradiator (Alpharetta, GA) available at the University of Vermont. The cells were seeded at a density of 3×106 per mL and 2 Gy dose at 1.7 Gy/min was administered at room temperature.
The cell culture Transwell inserts (Becton Dickinson Biosciences, Franklin Lakes, NJ) were used to study the bystander effect. 17 The bottom of the insert is a membrane with 1-μm pores at a density of 1.6×106 per cm2 to allow the passage of molecules. The polyethylene terephthalate membranes are highly permeable and allow increased rate of basolateral diffusion of nutrients and molecules of interest for transport, secretion, or binding studies. The Transwell culture insert dish has a growth area of 4.2 cm2, and the companion well of a six-well plate has a growth area of 9.6 cm2. The distance from the membrane of the inset dish to the bottom of the well of the companion plate is 0.9 mm.
The cells were plated into six-well dishes at a density of 2×105 per mL. The inserts containing 2×105 per mL irradiated cells were put into the wells of dishes and the six-well plates along with inserts were returned to the incubator. The irradiated and bystander cells were cocultured to allow the induction of X-ray–induced bystander effects until the time at which cells were harvested. The control sample was treated in the same way, except for irradiation. The treated cells and bystander cells were incubated at 37°C and harvested at 0, 4, 8, 12, and 24 hours for isolating RNA. The experiment was repeated in triplicate.
RNA isolation
The cells for RNA isolation were counted with a hemocytometer and approximately 5×106 cells were pelleted by centrifugation at 1500 rpm for 5 minutes and washed with 1 mL Dulbecco's phosphate-buffered saline without MgCl2 and CaCl2 (Invitrogen, Carlsbad, CA). Total RNA was isolated with RNAeasy™ kit (Qiagen, Valencia, CA). The quantity and quality of the total RNA was measured on NanoDrop 2000 (Thermo Scientific, West Palm Beach, FL) and by analysis on 2% agarose gels stained with ethidium bromide on BioSpectrum® Imaging System (UVP, Upland, CA).
Mitochondrial gene targets
Assays-on-demand for MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6, MT-CO1, MT-CO2, MT-CO2, MT-CYB, MT-ATP6, and MT-ATP8 (Table 1) were purchased from Applied Biosystems (Foster City, CA). Standard TaqMan assays have been designed using PrimerExpress software. RNA samples for gene expression analysis were normalized based on the TaqMan Gene Expression Assays for human endogenous hypoxanthine phosphoribosyltransferase (HPRT) gene.
Reverse transcription and cDNA synthesis reactions
Total RNA was treated with DNAse prior to reverse transcription to avoid genomic DNA contamination. The cDNA were generated from total RNA with random hexamer primers using a cDNA synthesis kit from Applied Biosystems following recommendations of the manufacturer. Reverse transcriptase reactions contained RNA samples, 50 nM hexamer primer, 1×RT buffer, 0.25 mM each of dNTPs, 3.33 U/μL MultiScribe™ reverse transcriptase, and 0.25 U/μL RNase inhibitor. The 15 μL reaction mixtures were incubated in Techne TC-312 thermocycler (Burlington, NJ) for 30 minutes at 16°C, 30 minutes at 42°C, and 5 minutes at 85°C and then held at 4°C. All reverse transcriptase reactions, including no-template controls and RT minus controls, were run in duplicate.
Quantitative real-time polymerase chain reaction and data analysis
Quantitative real-time polymerase chain reaction (QRT-PCR) was performed on an Applied Biosystems 7900HT Sequence Detection System by using a standard TaqMan PCR kit protocol. The 10 μL PCR mixture contained 0.67 μ RT product, 1×TaqMan Universal PCR Master Mix, 0.2 μM TaqMan probe, 1.5 μM forward primer, and 0.7 μM reverse primer. The reactions were incubated in a 384-well plate at 95°C for 10 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. The relative expression values of cycle thresholds were calculated using the comparative delta delta cycle threshold, ΔΔCT method, 26 by normalization to the endogenous control HPRT and to the control nonirradiated sample. The threshold cycle (C T) is defined as the fractional cycle number at which the fluorescence passes the fixed threshold. Statistical significance was determined using analysis of variance. The statistical and data analyses were performed with ABI prism and GraphPad Prism 5 software, both licensed to the University of Vermont. C T values of the 13 mitochondrial genes in irradiated and bystander cells were statistically evaluated using a one-way t-test (p<0.05). The experiments were repeated three times and the expression differences between the irradiated and bystander cells were statistically determined.
Results
TK6 cells were irradiated with 2 Gy dose of X-rays and cocultured with unirradiated cells using a Transwell system 9,17 to study the mitochondrial gene expression responses in both irradiated and bystander cells. At 0, 4, 8, 12, and 24 hours time points, the irradiated and bystander cells were harvested. The RNA isolated from the harvested cells was subjected to QRT-PCR analysis. The relative expression of 13 mitochondrial protein-coding RNAs (Table 1) including 7 members of NADH dehydrogenases and genes belonging to cytochrome c oxidase subunits, cytochrome b, and ATP synthases was examined. Differences and similarities in the relative gene expression among the irradiated and bystander cells were observed.
The expression analysis of NADH dehydrogenase 1 (MT-ND1), NADH dehydrogenase 2 (MT-ND2), NADH dehydrogenase 3 (MT-ND3), NADH dehydrogenase 4 (MT-ND4), NADH dehydrogenase 4L (MT-ND4L), NADH dehydrogenase 5 (MT-ND5), and NADH dehydrogenase 6 (MT-ND6) in irradiated and bystander cells during a 24-hour time period is shown in Figure 1. The expression of MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, and MT-ND6 was upregulated until it reached a maximum level at 24 hours in irradiated cells (Fig. 1). The MT-ND1 was only upregulated at the 24 hours time point (Fig. 1A). MT-ND3 was first downregulated at 8 hours postirradiation, followed by an upregulation in its expression levels until it reached the maximum expression at 24 hours (Fig. 1C).
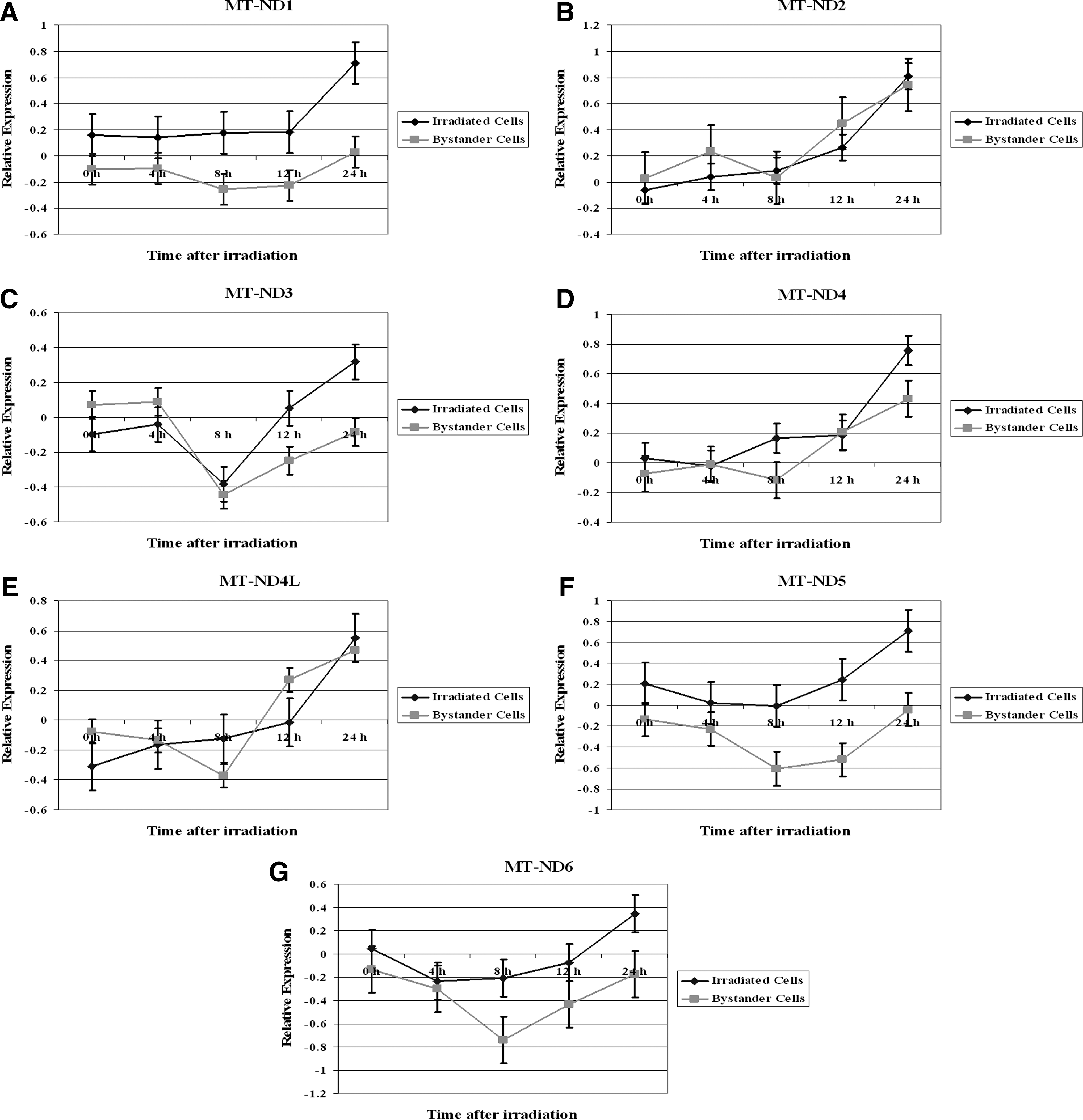
Relative expression of mitochondrial NADH dehydrogenases in TK6 and bystander cells. The cells were treated with 2 Gy X-rays dose and were cocultured with nonirradiated cells. The gene expression was assessed at 0, 4, 8, 12, and 24 hours postirradiation. Data are expressed as Log2.
The expression of seven members of mitochondrial NADH dehydrogenases in bystander cells revealed that MT-ND1, MT-ND5, and MT-ND6 remained downregulated throughout the 24-hour time period (Fig. 1A, 1F, and 1G, respectively). On the other hand, MT-ND2, MT-ND4, and MT-ND4L were upregualed in the bystander cells. The highest level of MT-ND2, MT-ND4, and MT-ND4L expression was seen at the 24-hour time period (Fig. 1B, 1D, and 1E, respectively).
There was no statistical difference in the expression of MT-ND2 (p=0.27), MT-ND3 (p=0.45), MT-ND4 (p=0.13), and MT-ND4L (p=0.68) between directly irradiated TK6 or bystander cells. In contrast, the expression of MT-ND1 (p=0.007), MT-ND5 (p=0.007), and MT-ND6 (p=0.02) was significantly different at most of the time points among the irradiated and bystander TK6 cells.
Next, the modulation of cytochrome c oxidase I (MT-CO1), cytochrome c oxidase II (MT-CO2), cytochrome c oxidase III (MT-CO3), cytochrome b (MT-CYB), ATP synthase 6 (MT-ATP6), and ATP synthase 8 (MT-ATP8) in irradiated TK6 and bystander cells (Fig. 2) was examined. In irradiated TK6 cells MT-CO1, MT-CO2, MT-ATP6, and MT-ATP8 were upregulated following the IR exposure (Fig. 2A, B, E, and F, respectively). The highest level of expression for these genes was observed at 24 hours after radiation treatment. An opposite effect was seen for the expression levels of MT-CYB (Fig. 2D). The MT-CYB was repressed after irradiation and the lowest expression was observed at 24 hours time point. MT-CO3 remained down regulated until the 12 hours time point, followed by an induction in its expression levels that reached a maximum value at 24 hours (Fig. 2C).
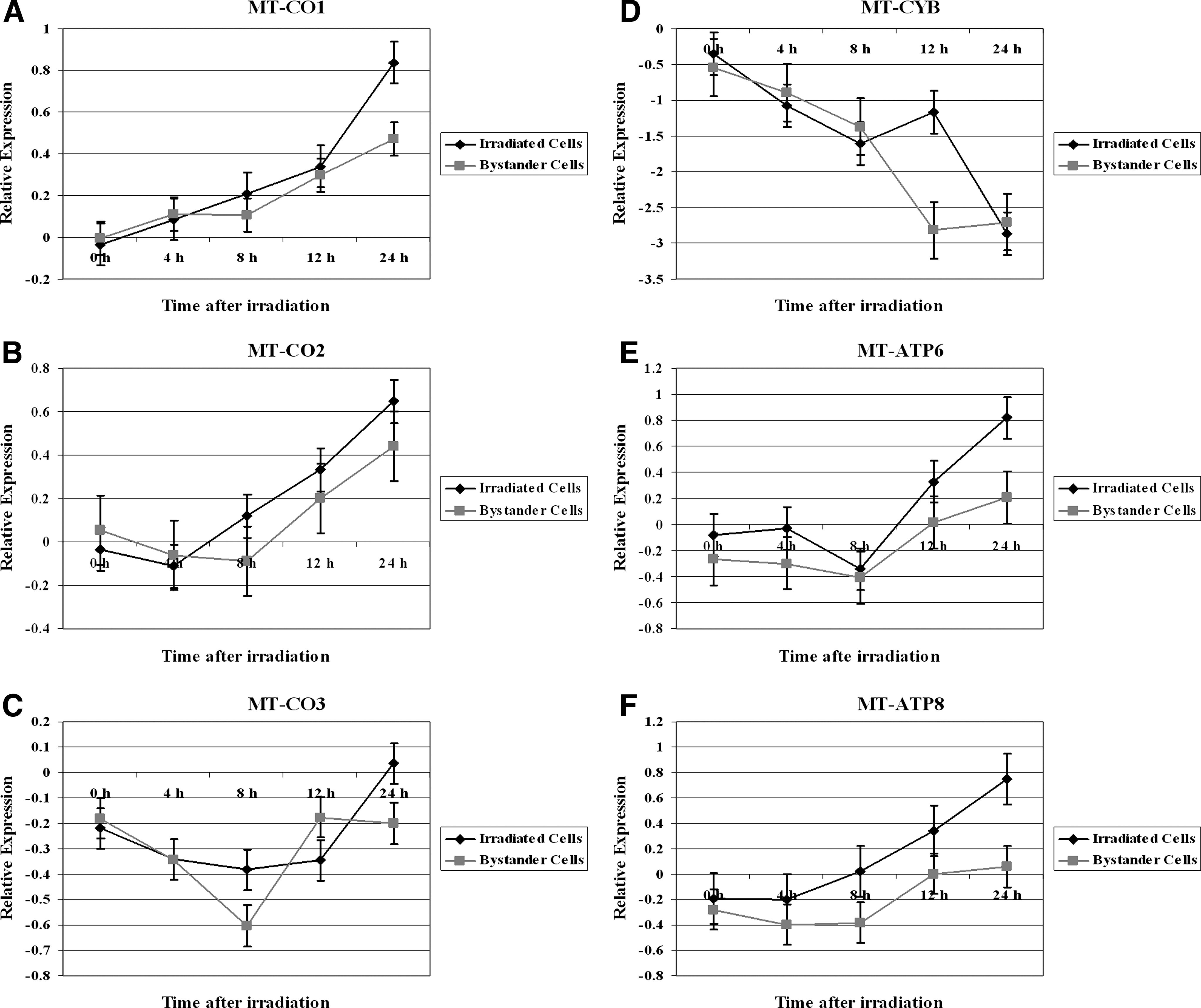
Relative expression of mitochondrial cytochrome c oxidase subunits, cytochrome b, and ATP synthases genes in 2 Gy irradiated TK6 and bystander cells. The gene expression was monitored at 0, 4, 8, 12, and 24 hours after radiation treatment. Data are shown as Log2 values.
The expression of MT-CO1, MT-CO2, and MT-ATP6 in bystander cells revealed that these genes were induced and their maximum induction was at the 24 hours time point (Fig. 2A, B, and E). MT-CO3 was first downregulated until 8 hours in bystander cells and its expression was upregulated after that time and plateau at 12 hours (Fig. 2C). The expression of MT-CYB was increasingly repressed and reached its lowest levels at the 12 hours time point (Fig. 2D). The expression of MT-ATP8 in bystander cells remained downregulated during the 24-hour time period following radiation.
The expression of MT-ATP6 (p=0.03) and MT-ATP8 (p=0.03) was significantly different among the irradiated and bystander TK6 cells. On the other hand, there was no statistical difference in the expression of MT-CO1 (p=0.28), MT-CO2 (p=0.26), MT-CO3 (p=0.55), and MT-CYB (p=0.52) between directly irradiated TK6 or bystander cells.
Discussion
Role of mitochondria in radiation-induced bystander effect
Exposure to IR causes increased mitochondria-dependent generation of ROS, with subsequent cell cycle checkpoint arrest, apoptosis, and stress-related responses. Free radicals are believed to play an active role in the bystander response. Cellular membranes and mitochondria have been identified as two distinct ROS producers. A complex interaction of direct irradiation and bystander signals leads to membrane-dependent amplification of cell responses. 22
The mitochondria play an important role in the regulation of radiation-induced bystander effects. The mitochondria-dependent NF-kappaB/inducible nitric oxide synthase (iNOS)/nitric oxide (NO) and NF-kappaB/cyclooxygenase-2/prostaglandin E2 signaling pathways are active in this process. 27 IR-induced stress response to mitochondrial dysfunction increases the number of mitochondria per cell. 28 In contrast, the cells, which lack mitochondria, are unable to produce a bystander signal, suggesting that an active mitochondrial function is required for these responses. 29 Inhibitors of NOS and mitochondrial calcium uptake provided evidence that NO and calcium signaling are part of the signaling cascade. 21 The cellular response to damage is complex and relies on simultaneous activation of a number of signaling networks. To elucidate the underlying mechanisms involved in the bystander effect, analyses of gene expression signatures in IR-induced oxidative stress are crucially important.
Mitochondrial gene expression in bystander cells
The regulatory mechanisms involved in the bystander response have not been elucidated. To provide insight into the pathways responding in bystanders, mitochondrial gene expression levels were measured. The bystander effect is known to occur in TK6 cells treated with 2 Gy dose of X-rays. 9 The focus of the present study was to determine whether the mitochondrial gene expression changes are involved in bystander cells. The data presented here show that alterations in the expression of mitochondrial genes are involved in the radiation-induced medium-mediated bystander effect.
mtDNA contains 13 genes that encode proteins of the OXPHOS complex that are involved in ATP generation. The relative expression of these 13 mitochondrial genes including 7 members of NADH dehydrogenases and genes belonging to cytochrome c oxidase subunits, cytochrome b, and ATP synthases was examined. Some of these genes have been shown to respond to IR treatment. There were significant differences in the relative expression of several mitochondrial NADH dehydrogenases among the directly irradiated and bystander cells. The NADH dehydrogenase subunit 4 level has been shown to increase after irradiation, with elevated expression persisting for at least 24 hours. 30 Nuclear genes related to the cell cycle or death (CDKN1A, GADD45A, NOTCH1, and BCL2L1) and cell communication (IL1B, TCF7, and ID1) were shown to be upregulated in irradiated cells, but not in bystander cells. 31 It has been previously reported that the radiation-induced nuclear gene expression profile in irradiated cells is different from unirradiated bystander cells. 32 The present results indicate that the changes in the expression of mitochondrial NADH dehydrogenases gene are induced in directly irradiated cells but not in the bystander cells. Mitochondria could be a target of signaling by IR, because it is known to be at the site of cellular oxidative stress. The differences in the modulation of mitochondrial NADH dehydrogenases might have a role in defining the responses of bystander cells after IR treatment.
The investigation of genes belonging to cytochrome c oxidase subunits and ATP synthases in irradiated TK6 cells indicated that most of these genes were upregulated in both irradiated TK6 and bystander cells. Previous studies have shown that MT-CO1 and MT-CO2 were induced after irradiation. 30 Cytochrome c is a pivotal protein that resides in mitochondria as component of mitochondria respiration and apoptosis initiator. Cytochrome c-deficient cells had attenuated ROS/NO and micronuclei induction to radiation-induced bystander signals, indicating that cytochrome c is essential for the bystander effect. 33 The proteins encoded by cytochrome c oxidase and other mitochondrial mRNAs characterized in this study are part of the mitochondrial respiratory chain, which produces ATP through the process of OXPHOS. These observations indicate that the mitochondrial genes regulating energy generation show similar responses in bystander cells. Similar nuclear gene expression patterns have been observed in irradiated and bystander cells. PTGS2, IL8, and BCL2A1 responded with an upregulation of expression in both bystander and directly irradiated cells. 34 No differences in the transcriptomes of cells grown in medium from X-irradiated cells or directly irradiated were seen. 4 The transcript profiles of cells grown in culture medium from irradiated cells concluded that X-irradiation and bystander effects induce similar changes of transcript profiles in most functional pathways. 35 Similar expression patterns of certain nuclear or mitochondrial genes in directly irradiated and bystander cells indicate common functions in the IR-induced pathways.
Mitochondria probably act as a sensor responding to the extracellular bystander signals. The inability to maintain functional mitochondrial metabolic capability as a response to the bystander signals can compromise cellular functions. These results imply that signaling between irradiated and bystander cells lead to mitochondrial transcriptional stress response in bystander cells. The disturbed mitochondrial gene expression levels in bystander cells may have a role in regulating the radiation response, leading to the induction of nontargeted cellular effects.
Footnotes
Acknowledgments
The authors thank Dr. Howard Liber (Colorado State University, Fort Collins, CO) for providing TK6 cells. The authors are thankful to the DNA analysis facility, University of Vermont, for assistance with the real-time PCR experiments. This work was supported by an endowment fund of the College of Nursing and Health Sciences, University of Vermont.
Disclosure Statement
The authors declare that no conflicts of interest exist.