
Abstract
Gambogic acid (GA), the main active component of gamboge, is well known for its marked antitumor effect in vitro and in vivo. The aim of this study was to assess the natural interaction between GA and chemotherapeutic agents, 5-fluorouracil (5-FU), oxaliplatin (Oxa), and docetaxel (Doc), which are widely used in gastric cancer treatment. This study also investigated the effect of GA on cell apoptosis and drug-associated gene expression for further mechanism research. Synergistic interaction on human gastric cancer BGC-823 cells and MKN-28 cells was evaluated using the combination index (CI) method. The double staining method with Annexin-V-FITC and PI was employed to distinguish the apoptotic cells from others. Expression of drug-associated genes, that is, thymidylate synthase (TS), excision repair cross-complementing (ERCC1), BRCA1, tau, and β-tubulin III, was measured by real-time quantitative RT-PCR. This study found that GA had a synergistic effect on the cytotoxity of chemotherapeutic agents against both cell lines. The combination of GA and chemotherapeutic agents could also induce apoptosis in a synergistic manner. The mRNA levels of TS, ERCC1, BRCA1, tau, and β-tubulin III were suppressed at 0.009, 0.075, 0.140, 0.267, and 0.624-fold, respectively, when cells were exposed to GA at the concentration of 0.25 μM. These data suggest that GA has a promising role in enhancing the efficacy of 5-FU, Oxa, and Doc in the treatment of gastric cancer. The potential mechanism would be their synergistic effects on apoptosis induction and the downregulation of chemotherapeutic agent-associated genes.
Introduction
Gastric cancer is one of the most common serious gastroenterological cancers. It is estimated that about 21,130 new cases have been diagnosed with a predictable 10,620 deaths in the United States in 2009. 1 According to the statistics, the 5-year relative survival rate of gastric cancer patients is 24.3%. The survival rates decrease remarkably along with tumor progression. For instance, 5-year survival rates are 61.1% and 3.4%, respectively, for patients with local and distant disease. 1,2 As recommended by NCCN, platinums, 5-fuorouracil (5-FU), and taxanes are considered as the first-line chemotherapy for the treatment of gastric cancer. However, the response rate of single drug chemotherapy remains lower than 20% and the chemotherapeutics sometimes lead to severe toxicity at their therapeutic dose. To achieve higher antitumor efficacy, delay the emergence of resistance and reduce the side-effect, a constant effort has been made in searching for novel chemotherapy in the field of cancer therapy.
Recent advances in the research of traditional Chinese medicine shed light on the discovery of novel adjunct to chemotherapy. Gambogic acid (GA, C38H44O8), the principal active component of gamboge and the resin from various Garcinia species, including G. morella and G. hanburyi, has been used as detoxification, homeostasis, anti-inflammatory, and parasiticide medicines for thousands of years. 3 Recent studies reported that GA activated apoptosis pathway in many cancer cell lines 4 –8 and inhibited human hepatoma tumor growth in SMMC-7721 9 as well as in mouse tumors such as Heps and S180 in vivo. 10 Induction of apoptosis and downregulation of telomerase activity 11 are possible mechanisms for the anticancer effect of GA. Recently, a differential apoptosis induction of GA was demonstrated on cancer cells and normal cells, suggesting that GA might be an effective anticancer drug candidate with low toxicity to normal cells. 12 A previous work indicated that GA is able to restore docetaxel (Doc) sensitivity in gastric cancer cell lines by inhibiting the expression of survivin. 13 The aforementioned findings showed the potential of GA to be a novel adjunct reagent to chemotherapy.
This preclinical study was therefore undertaken to investigate whether combination of GA and a series of chemotherapeutic agents resulted in a marked synergistic anticancer activity against gastric cancer cells. In this study, widely used anticancer drugs with different mechanisms were tested, including 5-FU, Doc, and oxaliplatin (Oxa). To elucidate further the mechanisms possibly involved in this action, the apoptosis induced by GA and chemotherapeutic agents was also investigated separately and in combination. The influence of GA on the expression of chemotherapeutic agent-associated genes was also examined.
Materials and Methods
Reagents
Doc, 5-FU, and Oxa were obtained from Jiangsu Hengrui Medicine Company. GA (99% purity) was supplied by the School of Pharmacy at China Pharmaceutical University. MTT (3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide) was purchased from Sigma Chemical Company. All other chemicals used were of the highest pure grade available.
Cell lines and cell culture
Two human gastric cancer cell lines, including poorly differentiated BGC-823 and well-differentiated MKN-28, were obtained from the Shanghai Institute of Cell Biology. Cells were cultured in RPMI 1640 medium (Gibco BRL) with 10% fetal bovine serum and 100 U/mL penicillin–streptomycin at 37°C in a water-saturated atmosphere with 5% CO2.
Cytotoxicity of GA and chemotherapeutic agents
Tumor cells, growing in log phase, were seeded at 2×103 cells per well into 96-well plates and allowed to attach overnight. Medium in each well was replaced with fresh medium or medium with various concentrations of drugs in at least five replicate wells and left for contact for 72 hours. The medium with drug was decanted and the IC50 doses of each drug were determined by MTT assay described later. The IC50 was defined as the concentration required for 50% inhibition of cell growth. Each experiment was allocated 10 wells containing drug-free medium for the control and performed on at least three separated occasions.
Determination of synergism and antagonism
Drugs were added either concomitantly or sequentially with six different concentrations of the single agents and six different concentrations of both agents at their fixed ratio based on their respective individual IC50 values for 72 hours. The fractional inhibition of cell proliferation was calculated by comparison to the control. Dose–response curves were obtained for each drug and for multiple dilutions of a fixed-ratio combination of two drugs.
Median effect analysis using the combination index (CI) method of Chou and Talalay 14 was employed to determine the nature of the interaction observed between GA and chemotherapeutic agents. The CI is defined by the following equation: CI=(D)1/(Dx)1+(D)2/(Dx)2+α(D)1(D)2/(Dx)1(Dx)2, in which (Dx)1 and (Dx)2 are the concentrations for D 1 (GA) and D 2 (chemotherapeutic agent) alone that gives x% inhibition, whereas (D)1 and (D)2 in the numerators are the concentrations of GA and another drug that produce the identical level of effect in combination. α=0 when the drugs are mutually exclusive (i.e., with similar modes of action), but α=1 when they are mutually nonexclusive (i.e., with independent modes of action). CIs>1 indicate antagonism, CIs<1 indicate synergy, and CIs=1 indicate additivity. Each CI ratio represented here is the mean value derived from at least three independent experiments.
Cytotoxicity assay
The in vitro drug-induced cytotoxic effects were measured by the MTT reduction assay. 15 After treatment, 1/10 volume of MTT was added to each well, and the plate was further incubated at 37°C for another 4 hours. Two hundred (200) microliters of dimethyl sulfoxide was added to each well to solubilize the MTT–formazan product after removal of the medium. Absorbance at 570 nm was measured with a multiwell spectrophotometer (BioTek). Growth inhibition was calculated as a percentage of the untreated controls, which were not exposed to drugs.
Apoptosis assay
Cells were cultured in a 60-mm Petri disk and allowed to grow to 75%–80% confluence. They were exposed to GA and anticancer drugs added singly or in combination for 48 hours. Then they were collected and incubated with Annexin-V-FITC (Bender Medsystems) and PI for determining surface exposure of phosphatidyl serine in apoptotic cells and necrotic cells. Analyses were performed with a FACScan flow cytometer (Becton Dickinson).
Quantitative RT-PCR
Total RNA was extracted from BGC-823 cells after 48 hours incubation with or without GA by using Trizol reagent (Invitrogen) and following the manufacturer's instructions. cDNA of 1 μg total RNA was synthesized using random primers and Primescript reverse transcriptase (Takara). Quantitative PCRs for indicated genes were carried out using SYBR green qPCR kit (Takara) by a fluorescent temperature cycler (Mx3000P Real Time PCR System; Stratagene). Primers sequences were as follows: thymidylate synthase (TS): 5′-ACCTGAATCACATCGAGCCA-3′ and 5′-TTGGATGCGGATTGTACCCT-3′; excision repair cross-complementing (ERCC1): 5′-GGGAATTTGGCGACGTAATTC-3′ and 5′-GCGGAGGCTGAGGAACAG-3′; BRCA1: 5′- GGCTATCCTCTCAGAGTGACATTTTA-3′ and 5′-GCTTTATCAGGTTATGTTGCATGGT-3′; Tau: 5′-TCACTTTTACAGCAACAGTCAGTG-3′ and 5′-TGCCATGTTGAGCAGGACTA-3′; β-tubulin III: 5′-GCGAGATGTACGAAGACGAC-3′ and 5′-TTTAGACACTGCTGGCTTCG-3′; β-actin: 5′-GCGAGAAGATGACCCAGATC-3′ and 5′-GGATAGCAACGCCTGGATAG-3′. The PCR conditions were 95°C for 10 seconds, followed by 45 cycles at 95°C for 5 seconds and 60°C for 20 seconds with a single fluorescence measurement. For the dissociation curve, reactions were incubated at 95°C for 10 seconds and ramped up from 55°C to 95°C with a heating rate of 0.1°C/sec and continuous fluorescence measurement.
Relative gene expression quantifications were calculated according to the comparative Ct method using β-actin as an internal standard. The melting curve of a product is sequence specific and can be used to distinguish nonspecific from specific PCR products. Gene expression was analyzed with the Stratagene analysis software and quantified by the 2−ΔΔCt method. 16
Statistical evaluation
Data were analyzed using SPSS 15.0 software and were expressed as mean±standard deviation (SD). Statistical analysis was performed using the Student's t-test. Values of p<0.05 were considered to be statistically significant.
Results
Cytotoxic effects of GA and chemotherapeutics on BGC-823 and MKN-28 cells
The cytotoxic effects of GA and other chemotherapeutics on cancer cells were evaluated by MTT assay. As expected, GA and three chemotherapeutics alone showed dose-dependent cell inhibition effects against both cell lines. Table 1 indicates the IC50s of GA and other chemotherapeutics against the two cancer cell lines. It is obvious that the four anticancer agents displayed antiproliferation effect in vitro. GA exhibited the strongest cytotoxity with the lowest IC50 on MKN-28 cells, with a value of 1.662±0.112 μM among the four agents. 5-Fu showed the weakest cell inhibition effect, with IC50 being 332.807±97.366 μM for BGC-823 cell line. The IC50 concentrations were then used to generate fixed ratios for subsequent combination studies and for the calculation of CIs.
IC50
Synergistic effects of GA and chemotherapeutics on gastric cancer cells
To explore whether GA could enhance the cytotoxicity of the chemotherapeutic agents used in the treatment of gastric cancer, the antiproliferation effects of 48-hour treatment with GA, 5-FU, Oxa, and Doc singly and in combination were examined. The combination ratios were designed to approximate the IC50 ratios of the individual component compounds, so that the contribution of antiproliferative effect for each compound in the combinations is roughly the same. 17
Both Figures 1A and 2A show the dose–response curves for BGC-823 and MKN-28 cell lines exposed to GA and chemotherapeutic agents separately and in combination. An apparent further cell survival decrease was observed in the combination group. To fully evaluate the nature of the interaction between GA and chemotherapeutic agents, the combination of drugs was analyzed using media-effect analysis, which resolves the degree of synergy, additivity, or antagonism at various levels of cell death.
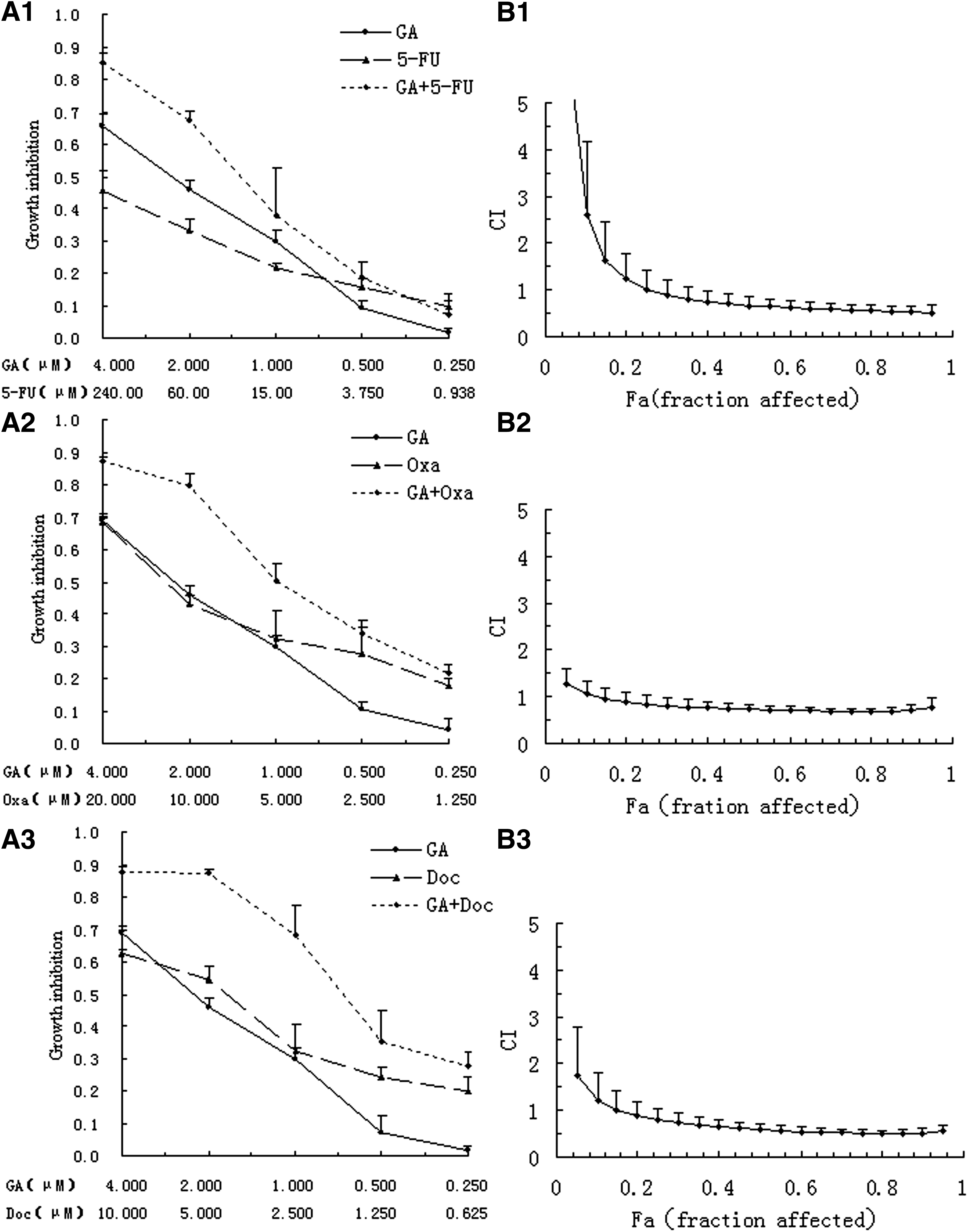
Analysis of synergy between gambogic acid (GA) and 5-fluorouracil (5-FU)/oxaliplatin (Oxa)/docetaxel (Doc) on BGC-823 cells.
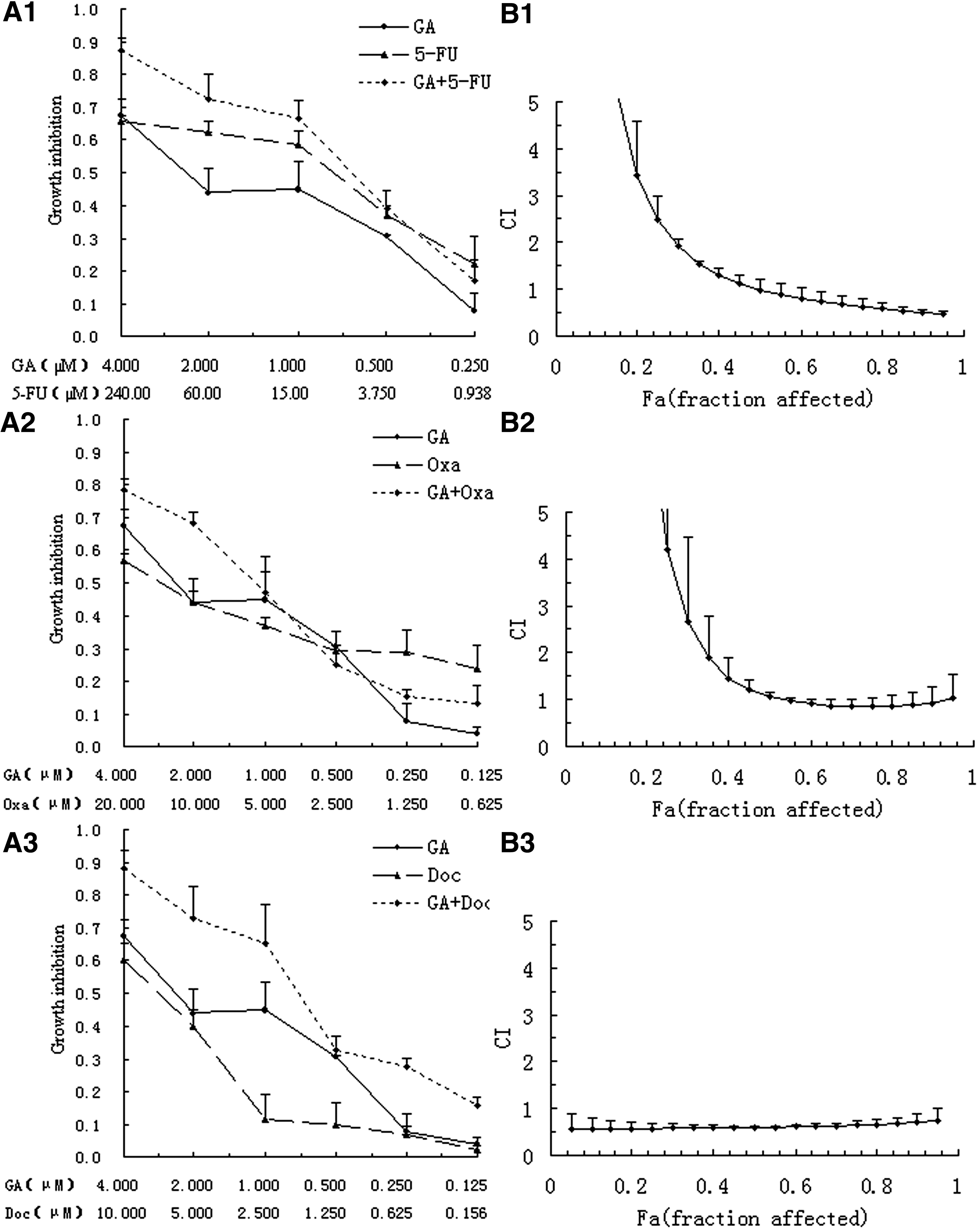
Analysis of synergy between GA and 5-FU/Oxa/Doc on MKN-28 cells.
Both Figures 1B and 2B illustrate media-effect analysis result with CIs on different fractional cell growth inhibition (FA) of BGC-823 and MKN-28, respectively. As for the combination of GA and anticancer drugs, the combination of two drugs generated more cell death than either of the drugs used separately in different range for different drugs and cells. In BGC-823 cells, media-effect analysis showed that the CI values were below 1 when FA reached 0.35 in the 5-FU group, and 0.05 in either the Oxa or DOC group, indicating a synergistic antiproliferative effect of GA and anticancer drugs against BGC-823 at this range. As for MKN-28 cells, the CI values were below 1 across almost the entire dose inhibition range when the GA combined with Doc, and CI values were below 1 when FA reached 0.5 in both the 5-FU and Oxa groups, which also indicated a more pronounced synergistic effect for the two drug combination.
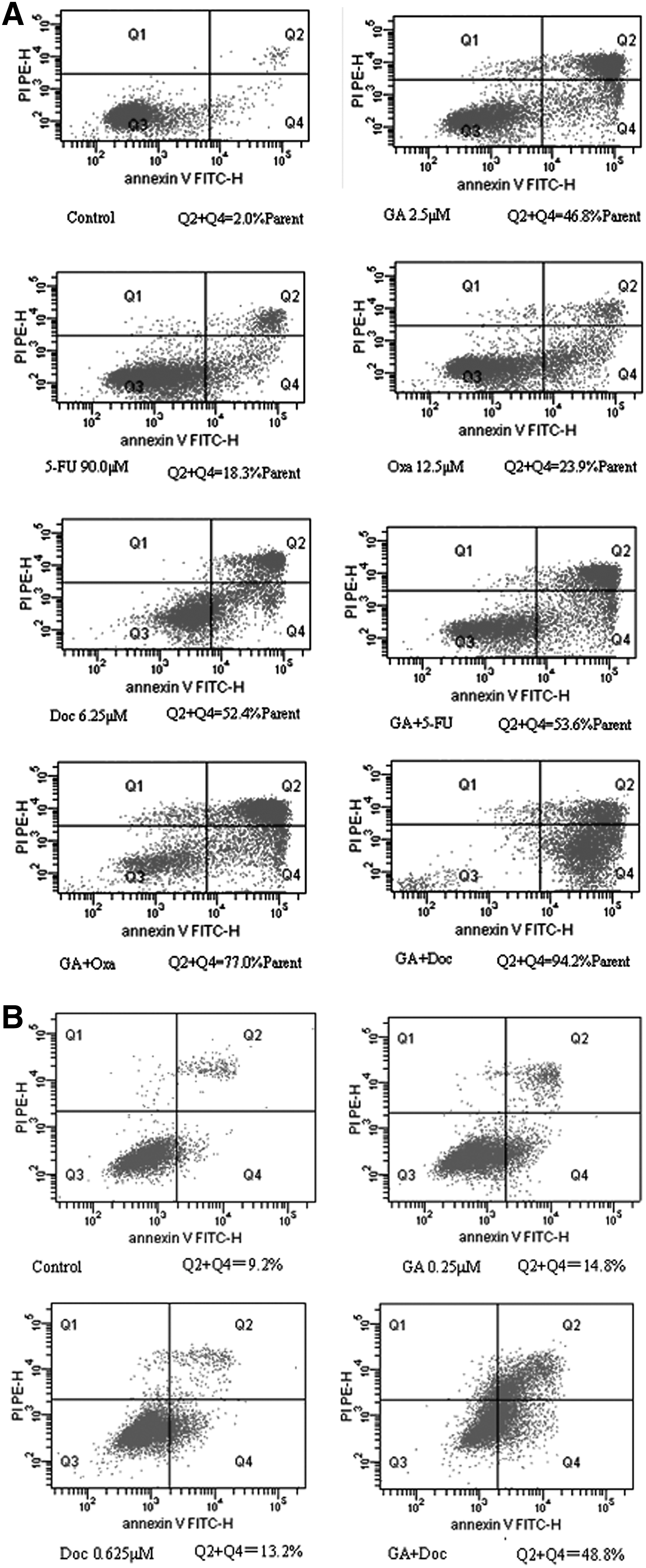
Apoptosis effects mediated by GA and chemotherapeutic agents
The apoptotic rates were analyzed using the Annexin-V-FITC and PI staining method 18 with different concentrations for each drug according to the previous results of MTT assay. Apoptosis was observed on BGC-823 cells with the rates of 46.8% for 2.5 μM GA, 18.3% for 90 μM 5-FU, 23.9% for 12.5 μM Oxa, and 52.4% μM for Doc, respectively. The combination of GA and chemotherapeutics led to moderate to apparent enhancement in the apoptotic rates for all the tree drugs (53.6% for GA+5-FU, 77% for GA+Oxa, and 94.2% for GA+Doc; Fig. 3A). A similar result was obtained on MKN-28 cells treated with GA and Doc (14.8% for GA, 13.2% for Doc, and 48.8% for combination; Fig. 3B).
GA modulates mRNA expression of chemotherapeutic agents-associated genes
To extend the observation made in two gastric cancer cell lines that the cytotoxicity of chemotherapeutic agents was potentiated in the presence of GA, the present study hypothesized that GA might modulate the expression of chemotherapeutic agents-associated genes, which was widely used to predict patients' outcome after chemotherapy. Five genes were selected, including TS, ERCC1, BRCA1, tau, and β-tubulin III for 5-FU, Oxa, and Doc, respectively. mRNA expression levels of these genes in BGC-823 cells treated with GA were assessed by quantitative RT-PCR. As shown in Figure 4, a dramatic change was observed in the expression of those genes. The mRNA levels of TS, ERCC1, BRCA1, tau, and β-tubulin III in BGC-823 cells were suppressed at 0.009 (p=0.0000), 0.075 (p=0.0000), 0.140 (p=0.0000), 0.267 (p=0.0001), and 0.624 (p=0.0001)-fold after 48 hours incubation with GA (0.25 μM). Although the mechanism of the interaction of GA and chemotherapeutic drugs was not clear, it is notable that GA could influence the expression of drug-related genes, which might increase the sensitivity to these agents.
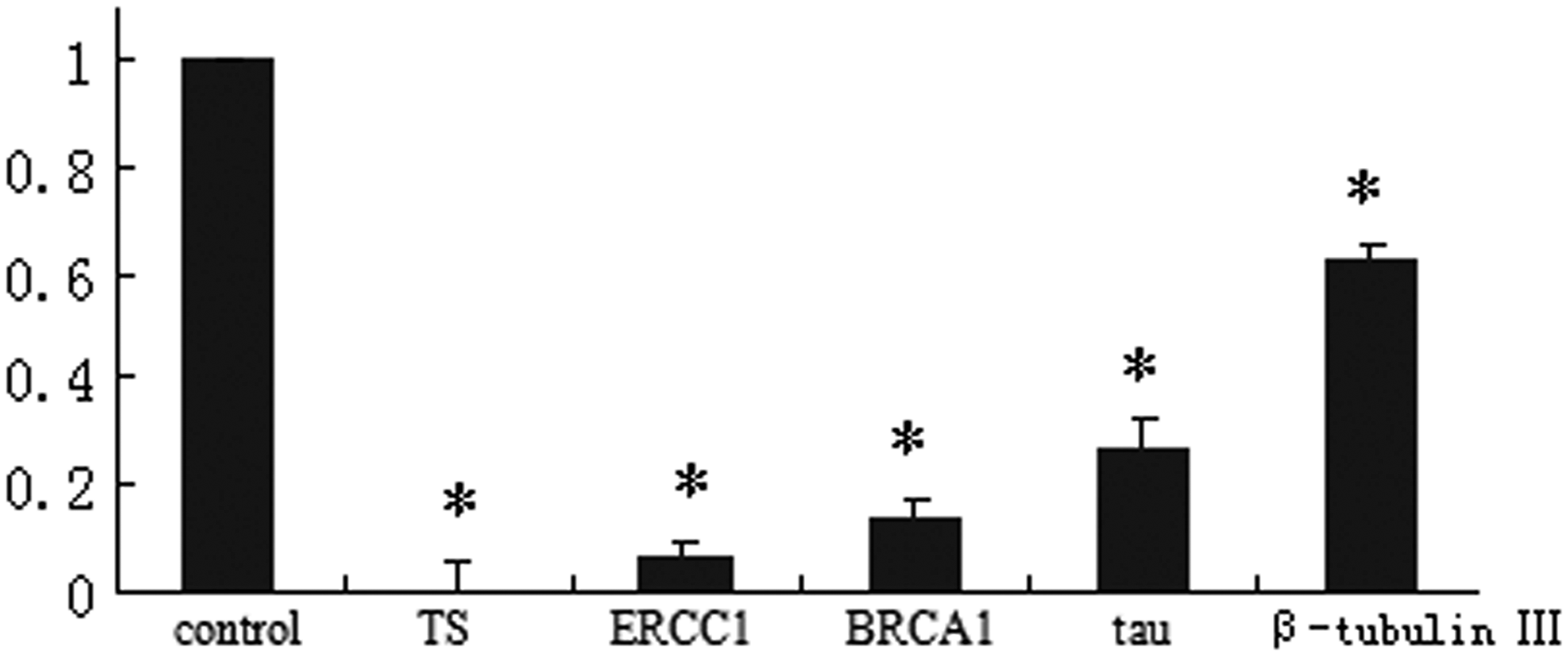
GA downregulated the drug-related gene expression. The mRNA expressions of drug-related genes, including thymidylate synthase (TS), excision repair cross-complementing (ERCC1), BRCA1, β-tubulin III, and tau in BGC-823 cells, were determined by quantitative RT-PCR after 48 hours of GA (0.25 μM) treatment. *p<0.01 compared with control.
Discussion
As recommended by NCCN, the first-line chemotherapy for gastric cancer is composed of the three aforementioned drugs. To overcome the probability of severe systemic toxicity or the low response rate of chemotherapy, more attention has been drawn on the discovery of small molecules that could synergistically enhance the anticancer effect of chemotherapeutics. Recently, plant-derived compounds attracted intensive interests for their potential antiproliferative properties and low toxicities. Moreover, several studies reported that these compounds, such as genistein, curcumin, and emodin, are capable to sensitize cancer cells to chemotherapeutic agents. 19 Previous studies have demonstrated the potential anticancer effect of GA in vitro and in vivo. 9,20 –23 GA, as reported by Yang et al., 12 has differential apoptotic induction on hepatoma cells and normal hepatocytes, which indicated that GA might be a kind of highly effective anticancer drug candidate with low toxicity to normal cells. In the present study, it is reported for the first time that the combined application of GA and three drugs (5-Fu, Doc, and Oxa) could exhibit synergistic and efficient antitumor effects on human gastric cancer cell lines, MKN-28 and BGC-823.
In the present report, GA revealed its potential to augment the cytotoxicity of commonly used chemotherapeutics in a synergistic manner. Flow cytometry analysis demonstrated that the synergistic proliferation inhibition effects by the treatment of GA and chemotherapeutics was possibly based on the apoptosis of cells. Cellular apoptosis has been reported to be the mechanism of antiproliferation effect of both classical chemotherapeutics (such as 5-Fu, Doc, and Oxa) and plant-derived GA. 24,25 Cell cycle arrest plays an important role in cell growth inhibition, and G2/M arrest commonly results in apoptosis. G2/M arrest on BGC-823 cells has been previously observed by the present study authors 13 and also by another research group. 8 Therefore, the present results probably implied that the enhanced cytotoxicities of GA and chemotherapeutics in combination might be based on a synergistic effect on apoptosis.
Previous studies have confirmed the association between drug sensitivities and mRNA levels of drug-related genes. As reported, TS is a key gene involved in 5-Fu metabolism. High TS expression level is reported to be associated resistance to 5-FU and poor clinical outcome. 26 Additionally, recent studies reported that tumor cells with high ERCC1 or BRCA1 expression are resistant to platinum-containing chemotherapy, 27,28 and mRNA expression levels of the two genes have been shown to be independent prognostic markers for patients treated with platinum-based chemotherapy. 28,29 Similarly, β-tubulin III and tau are closely related to taxanes resistance. 30,31 The present study demonstrated that the mRNA expression of 5-Fu-related genes (TS), Doc-related genes (β-tubulin III and tau), and Oxa-related genes (ERCC1 and BRCA1) in BGC-823 cells was decreased significantly after GA treatment in BGC-823 cell line. It could be supposed that the synergistic effect of GA and chemotherapeutics may be partly due to the downregulation of drug-related genes.
In conclusion, the present study demonstrated for the first time that GA could synergize with several chemotherapeutics against human gastric cancer cell lines. The underlying mechanisms may be related to the enhanced cell apoptosis and downregulation of drug-related genes. Collectively, as GA demonstrated potent antiproliferation effect and synergistic effect, it could be used as a potential drug sensitizer for the application of first-line chemotherapeutics. It is undoubtful, however, that the development of Traditional Chinese Medicine as drug sensitizers warrants more intensive research to evaluate the feasibility and advantages of clinical applications.
Footnotes
Acknowledgments
This work was supported by the Nature Science Foundation of Jiangsu Province (BK2008064).
Disclosure Statement
No competing financial interests exist.