
Abstract
The mechanisms underlying tumor dormancy in human primary lesions and bone marrow metastases of nasopharyngeal carcinoma (NPC) are still not completely understood. The aim of this study was to determine differences in the fates of cultured primary NPC (P-NPC) cells, interferon-γ–transduced primary NPC (IFN-γ-P-NPC) cells, bone marrow metastatic NPC (BM-NPC), and IFN-γ–transduced BM-NPC (IFN-γ-BM-NPC) cells following xenotransplantation into these four groups of SCID mice through subcutaneous injection of 5×106 cells/site/animal (4 animals/group). The injected mice were monitored for tumor development at the sites of injection. In only the group injected with IFN-γ-P-NPC cells, the resulting nodules remained small throughout the 60-day observation period after injection, but gradually became palpably prickly. Histopathological examination revealed that these lesions invariably consisted of mostly structures of horny pearls and keratin bridges with occasional apoptotic and degenerative cells. In contrast, animals injected with nontransduced-P-NPC cells developed tumors progressively with occasional central necroses. In the two groups injected with IFN-γ-NPC-BM and NPC-BM cells, progressive growths of tumors were noted, with the latter being at slightly faster rates, whereas the xenografts of both groups showed a poorly differentiated phenotype with abundant vascularity. The study results highlight the high susceptibility of P-NPC but not BM-NPC following IFN-γ gene transfer to the induction of tumor dormancy, which is mediated via induced cell differentiation. Thus, induced cell differentiation could provide a new mechanism by which tumor dormancy is induced.
Introduction
The formation and progression of malignant tumors is a complex process, as it involves the interaction of tumor with its microenvironment. The state of tumor dormancy might occur at different stages of cancer development, from early stages of primary cancers to remnants of primary cancers, which could be “recurrences,” to dormant micrometastases in distant sites, which could, after a long period of tumor latency, be activated and evolved into clinical malignant entities. A number of mechanisms have so far been proposed to be involved in the process of tumor dormancy, including cell cycle arrest, hormonal dependence, inhibition of tumor angiogenesis, immune surveillance, and the influence of microenvironment. 1 –3 Accumulative evidence indicates that the concept of tumor dormancy is important to understand such discontinuous disease progress as found in some animal as well as human malignancies. As there are different mechanisms by which tumors do not progress with prolong latency periods, understanding the process of tumor dormancy is fundamental for both early tumor detection and for patient treatment and management.
Interferon-γ (IFN-γ) is known to induce enhanced expression of certain key immune regulatory molecules such as major histocompatibility complex class I and class II molecules on the surface of tumor cells. Through this process, it was hoped that improved antigen presentation to dendritic cells as well as to preexisting primed cytotoxic T lymphocytes (CTLs) for further expansion and therefore enhanced destruction of tumor cells could be achieved. 4 –6 One approach toward this end is to assure the continuous presence of IFN-γ in the tumor lesion by introducing the IFN-γ gene into the tumor cells constitutively. 4,6,7 When such IFN-γ gene-modified tumor cells are irradiated and injected back to the patient, it was thought that the resultant IFN-γ production at the tumor site should theoretically provide a permissive microenviornment favorable to trigger potent T-cell immune response, leading to the generation of tumor-specific CTLs, 4,6 although a contrary to this assertion has also been raised. 8
Nasopharyngeal carcinoma (NPC), which is strongly associated with Epstein-Barr virus as part of its link to tumorigenesis, is a tumor of epithelial origin with endemic distribution among certain ethnic groups and geographic regions. 9 –11 The highest incidence of the disease has been noted among southern Chinese, who live in Guangdong and Guangxi provinces as well as in certain parts of Taiwan. NPC patients with early stages (I and II) are currently treated with radiotherapy alone and have achieved a 5-year survival rate of greater than 75%. A 5-year survival rate of NPC patients with stage III or IV after radiotherapy vary considerably, ranging from 18% to 52%, despite improved results with combined use of radiotherapy and chemotherapy. NPC patients with metastatic disease diagnosed at the beginning of therapy almost all expire within 4 years. Therefore, understanding the differences between primary and metastatic NPCs is needed to devise more effective therapeutic strategies. Although most available NPC cell lines are of primary tumor origin, 12 –15 cell lines derived from metastatic sites are rarely available to be studied. 16 Following bone, lung, and liver, bone marrow has been reported as the fourth common metastatic site for NPC, with an incidence of 22%. 17 It is believed that bone marrow provides an enduring reservoir/sanctuary from which tumor cells could spread to other sites. 18,19 The biology of these bone marrow metastatic tumor cells has remained largely unknown. 20 –22 The invasive/metastatic phenotype of the tumor cells in bone marrow is reflected by their epithelial-to-mesenchymal transition and expression of several metastasis-associated molecules, which may be potential candidates for therapeutic targets. 18,23 –25 A number of NPC cell lines derived from bone marrow metastatic lesions collectively termed “BM-NPCs” have been established in this laboratory with some of their features described in comparison with those of NPC cell lines derived from primary sites termed “P-NPCs.” 8,16,26
Recently, it was noted that cultured P-NPCs and BM-NPCs could be distinguished from one another on the basis of their differential constitutive release of IFN-γ and interleukin (IL)-6 and so were between their response profiles to IFN-γ transduction in terms of the production of IFN-γ, IL-6, and IL-6Rα. 8 To extend this observation, this study describes the distinct differences in the fate of xenotumorigenicity in SCID mice between IFN-γ–transduced cultured P-NPC cells and nontransduced P-NPC cells and between IFN-γ–transduced cultured P-NPC and IFN-γ–transduced BM-NPC cells following subcutaneous injection of these tumor cell preparations. Surprisingly, only in the group of mice given IFN-γ–transduced P-NPC cells, the nodules at the injection sites were found to remain very small over a long period of observation. In contrast, animals in the groups of nontransduced P-NPC and IFN-γ–transduced and nontransduced BM-NPC cells progressively grew at different rates in the injected animals.
Materials and Methods
Cell lines and culture conditions
Human NPC cell lines used were NPC039 and NPC076, which were derived from NPC primary sites 14 and were gifts from Dr. C-T Lin, Department of Pathology, National Taiwan University Hospital, Taipei, Taiwan. The cell lines NPC-BM1, NPC-BM2, and NPC-BM3 derived from bone marrow metastatic lesions of three different NPC patients were established in this laboratory. 8,26 The comparative features of IFN-γ–transduced counterparts of the NPC039, NPC076, and NPC-BM1 cell lines have been previously described. 8 The culture medium used for these cell lines was RPMI-1640 (Gibco, Grand Island, NY) supplemented with 1 mM sodium pyruvate, 2 mM glutamine, 100 U/mL penicillin-G, 100 μg/mL streptomycin sulfate, and 10% fetal bovine serum (FBS). Cultures were maintained at 37°C in an incubator with 5% CO2, 95% air, and humidified atmosphere. In the routine maintenance of these cell lines, medium was changed every 3–4 days. When growth reached near confluence, cells were subcultured or, intermittently, were cryopreserved in RPMI-1640 medium containing 20% FBS and 10% dimethyl sulfoxide in liquid nitrogen.
Monoclonal antibodies
The monoclonal antibody (mAb; clone W6/32) to a monomorphic determinant of HLA class I molecules associated with β2-microglobulin was purchased from Dako Japan Co. Ltd. (Kyoto, Japan). Both mAb to HLA class II (clone DK22, specific for the β-chain of all DR loci, all DP loci, DQw1 but not with DQw3 locus) and mAb to ICAM-1 (CD54; clone 6.5B5) were purchased from DakoCytomation (Glostrup, Denmark).
Cytofluorometric analysis
Monodispersed cells harvested from monolayer cultures by trypsinization were distributed into test tubes (5×105 cells/tube) and centrifuged at 400 g to pellet the cells. Test antibody samples (5 μg/mL; 100 μL/tube) were added. Bound antibodies were detected by fluorescein isothiocyanate–conjugated polyclonal antibodies (goat anti-mouse IgG; Coulter Immunology, Hialeah, FL) by flow cytometry according to the procedures previously described. 8 Appropriate positive and negative control antibodies, such as anti-HLA class I (mAb W6/32), phosphate-buffered saline, and isotype-matched irrelevant mAbs were included in each experiment.
SCID mouse xenotransplantation experiments
Tumor cells at 5×106 per site were subcutaneously injected into the area above the hind leg of SCID female mice at about 8 weeks old. The injected animals were monitored for the tumor development at the injection sites and sacrificed at the end of experiments for histopathology. The volume of palpable tumor nodules was estimated according to the formula: volume (mm3)=0.4 a×b 2 , where a is the major tumor diameter and b is the minor diameter perpendicular to the major one. 27 The tumor masses resulting from injection with nontransduced and IFN-γ–transduced cells were aseptically removed and dissected for cell culture, and a portion of the tumor was fixed in formalin followed by embedding in paraffin. The tumor blocks were sectioned and stained with hematoxylin and eosin (H&E). The slides were then examined by light microscopy.
Results
Differential effects of transduction of IFN-γ gene into P-NPC and BM-NPC cell lines on the secretion of IL-6 and IL-6Rα
First, the secretion of IL-6 and IL-6Rα in the spent media of two P-NPC cell lines and one BM-NPC cell line harvested at the end of day 3 cultures was investigated. Both IFN-γ–transduced and nontransduced cells were included for each cell line tested. Levels of IL-6 significantly increased in both IFN-γ–transduced P-NPC cell lines (NPC039 and NPC076) up to >500 pg/mL, from marginal levels (∼40 pg/mL) secreted by their nontransduced counterparts (Fig. 1A). In contrast, a rather high level of IL-6 (1349.3 pg/mL) was constitutively secreted by the BM-NPC (NPC-BM1) cells, the level of which was increased only very slightly in the spent medium of IFN-γ–transduced counterparts.

Levels of IL-6
Using the same spent culture media, the levels of IL-6Rα secreted by the same panel of cell lines for IL-6 (Fig. 1B) were examined. High levels of this receptor (841.5 and 627.6 pg/mL) were detected in the spent media of NPC039 and NPC076 cells, respectively, whereas decreased IL-6Rα levels were found in the spent media of the IFN-γ–transduced P-NPC cell lines. As for BM-NPC cells, a rather low level of baseline IL-6Rα (204.3 pg/mL) was detected in the nontransduced cells, and essentially no increase in this receptor level (205.7 pg/mL) was found in the cultured supernatant of IFN-γ–transduced cells. These results are essentially similar to those previously observed. 8
Comparative in vitro growth characteristics between IFN-γ–transduced and nontransduced P-NPC cell lines and between IFN-γ–transduced and nontransduced BM-NPC cell lines
As illustrated in Figure 2, in both P-NPC (NPC039 and NPC076) cell lines grown as monolayers, cell numbers increased as a function of time up to day 3, when saturation was reached, and thereafter declined rather quickly, which coincided with a drop in their cell viability. In these two cell lines, which were successfully transduced with the IFN-γ gene, monolayer cells appeared to stay together tighter and longer, as evident by sustaining the degrees of saturation density as well as their cell viability after day 3 of culture. In contrast, the IFN-γ–transduced and nontransduced BM-NPC (NPC-BM1) cell lines grown as monolayers exhibited very similar growth pattern in that both maintained their saturated states firmly once they reached plateau at day 3, a phenomenon being quite different from that exhibited by nontransduced P-NPC cells.
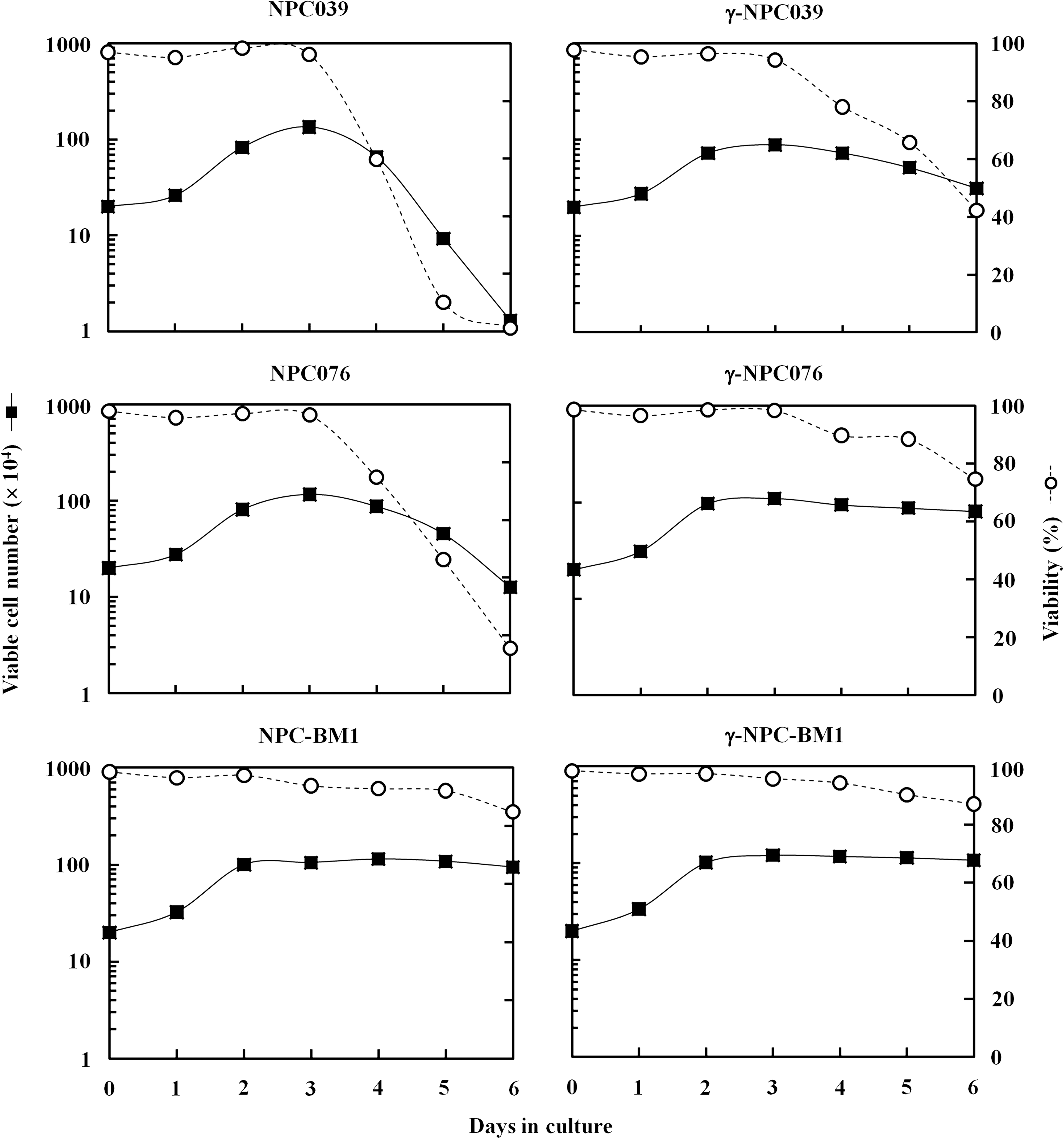
Growth curves of nontransduced and IFN-γ–transduced P-NPC (NPC039 and NPC076) and BM-NPC (NPC-BM1) cell lines. Cells were grown in culture dishes by complete medium for 6 days without change of medium. Triplicate cultures were set up for each time point. Results are expressed as viable cell number and viability. γ-NPC039, γ-NPC076, and γ-NPC-BM1 stand for IFN-γ–transduced NPC039, NPC076, and NPC-BM1 cells, respectively. P-NPCs, cultured cell lines derived from a primary tumor lesion; BM-NPCs, cell lines derived from bone marrow metastasis.
Tumorigenicity of IFN-γ–transduced and nontransduced P-NPC and BM-NPC cell lines into SCID mice
In xenotransplantation experiments, subcutaneous injection of four experimental groups of SCID mice with 5×106 cells of nontransduced and IFN-γ–transduced NPC076 and NPC-BM1 cells resulted in different incidences of tumor formation (Fig. 3A, B). Results showed that only the group receiving IFN-γ-NPC076 cells out of the four experimental groups did not develop measurable tumors and yet the small nodules (4/4) at the injection sites became palpable and remained small and prickle up to 60–63 days after tumor injection. These nodules consisted of mostly structures of horny pearls and keratin bridges with occasional apoptotic and degenerative cells. In contrast, other groups of animals injected with nontransduced-P-NPC cells developed tumors progressively. In groups of animals injected with either IFN-γ-NPC-BM1 cells or NPC-BM1 cells, progressive tumors clearly developed, with the latter being at slightly faster rates.
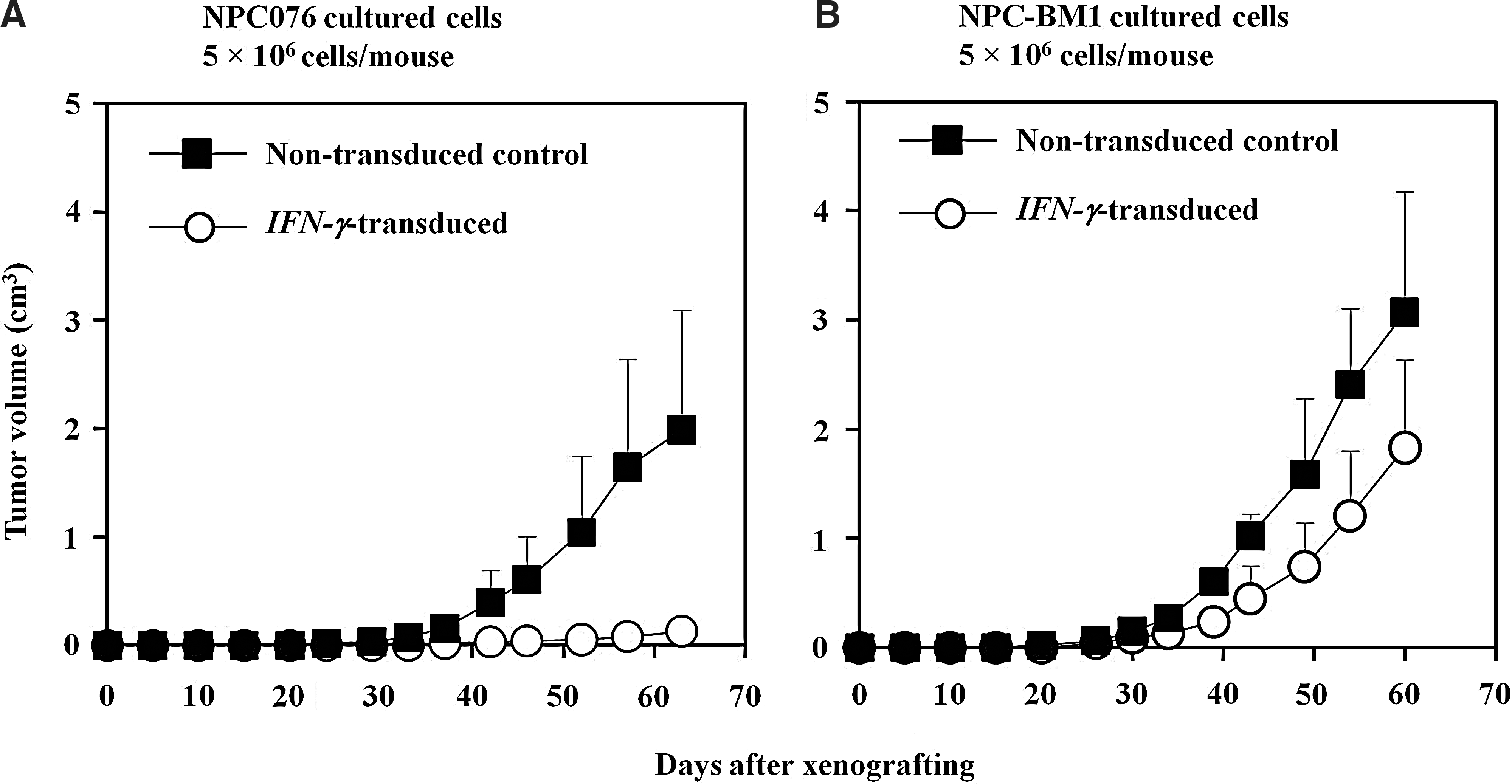
Growth curves of nontransduced and IFN-γ–transduced NPC076 cells
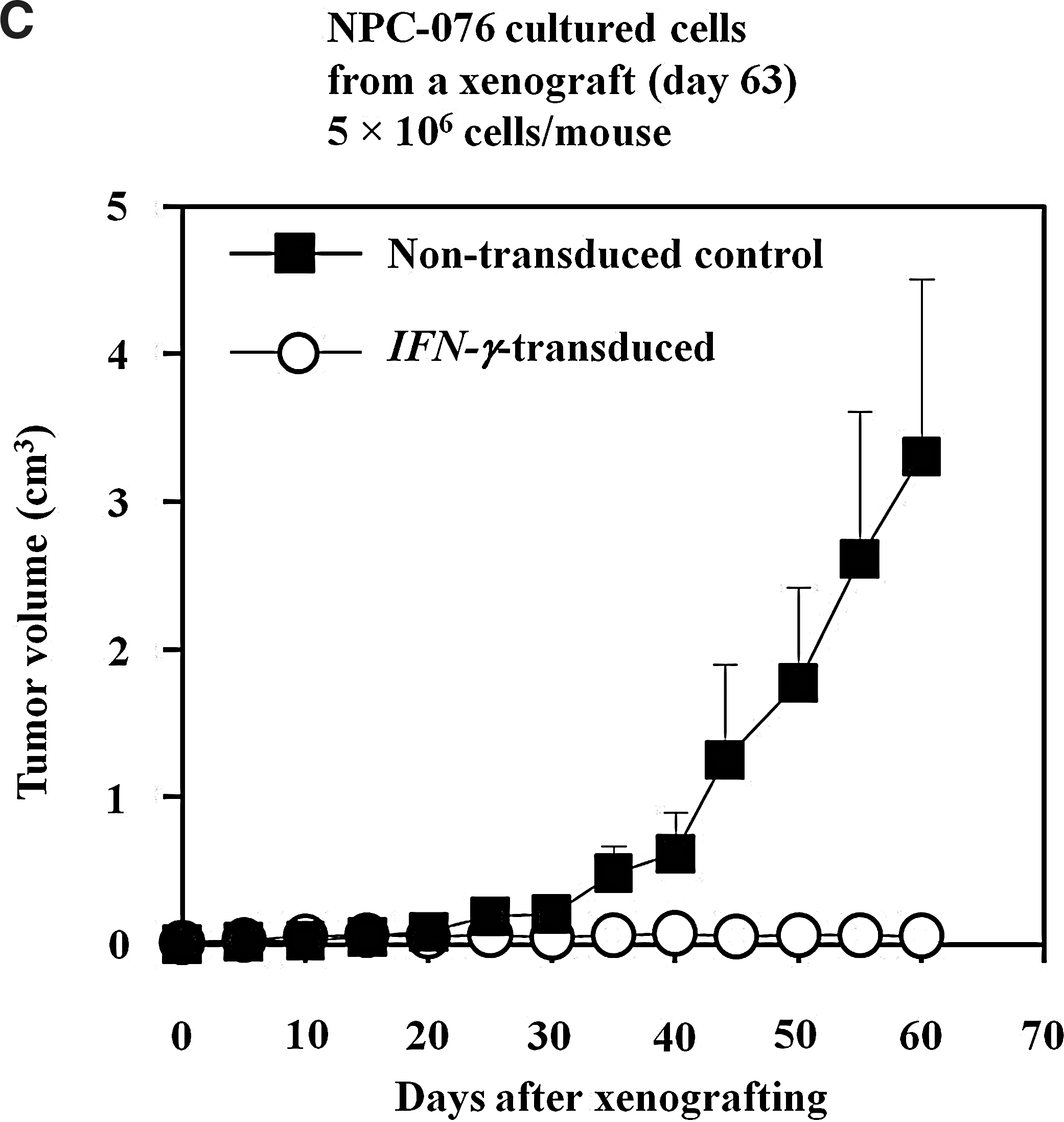
When dispersed cells prepared from one of the four xenografts resulting from IFN-γ-NPC076 and from nontransduced NPC076 cells were cultured in vitro, cultures from both groups grew very well and could be nicely passaged in culture as for before xenografting. To confirm the induction of tumor dormancy observed with IFN-γ-NPC076 cells as seen in Figure 3A, 5×106 cells of nontransduced NPC076 and IFN-γ-NPC076 cells were injected into 4 animals for each group (Fig. 3C). Very similar patterns of results were obtained, in which IFN-γ-NPC076 cells again failed to develop progressive growth of tumors with prickly palpable lesions, whereas the nontransduced counterpart developed tumors progressively at the injection sites in all 4 animals injected. Of note is that a shorter latent period of about 25 days and greater progressive growth with nontransduced NPC076 cells recovered from the xenograft (Fig. 3C), compared with nontransduced P-NPC cells never passaged in SCID mice before (latent period of ∼35 days and less steeper growth curve; Fig. 3A). This discrepancy was likely due to the selection of more invasive variants or enrichment of more cancer stem cells (CSCs) in the cultured cells reestablished from the xenograft.
Similar results of the successful induction of tumor dormancy were obtained with IFN-γ–transduced NPC039 cells, but not with their nontransduced counterparts in SCID mice (data not shown).
Gross description and histopathology of resulting tumor nodules
Examination of H&E-stained sections showed that in tumor nodules resulting from injection of IFN-γ–transduced NPC076 cells, the lesion was always small in size throughout the 60-day observation period and gradually became palpably prickly about 25 days postinjection. Histopathologically, the lesion was composed of structures of horny pearls and keratin bridges in most of central areas, accompanied by some hyperplastic epithelial cells in the periphery, which were partially surrounded by fibrotic tissues (Fig. 4A). In other areas, there were groups of epithelial cells with clear cytoplasms and densely stained pyconotic nuclei, representing degenerating cells (Fig. 4B). Some macrophages or lymphoid-liked cells were occasionally noted in the stromal areas. In the tumor lesions resulting from injection of nontransduced NPC076 cells, squamous cell-like morphology and lightly stained nuclei in areas of papillary tumor islands with occasional necrotic areas were seen (Fig. 4C). Muscular layers of the skin at the injection sites were found to be invaded by the tumor cells.

Representative histopathological micrographs of xenografts resulting from injecting IFN-γ–transduced NPC076
Tumor lesions resulting from the injection with IFN-γ–transduced and those with nontransduced NPC-BM1 cells were compared. The nontransduced lesions appeared larger in size, but shared their histopathological features very closely with their IFN-γ–transduced counterparts. As depicted in Figure 4D, tumor cells exhibited a more undifferentiated and invasive phenotype with rarely central necroses detected. Moreover, invasion of muscular layers with abundant vascularity was also evident in the xenografts resulting from both IFN-γ–transduced and nontransduced NPC-BM1 cells.
Discussion
This study described the differential susceptibility of IFN-γ–transduced P-NPC cell lines (NPC039 and NPC076) and BM-NPC cell line (NPC-BM1) to the induction of tumor dormancy in SCID mice following subcutaneous injection. The tumor dormancy was observed with only IFN-γ–transduced P-NPC cells, but not with nontransduced P-NPC and with IFN-γ–transduced BM-NPC or nontransduced BM-NPC cells injected. The chief characteristic of tumor dormancy observed in this study was the formation of horny pearls and keratin bridges, accompanied by some degenerative clear cells, apoptotic cells, and some viable cells in the tumor dormant lesions. The formation of horny pearls and keratin bridges, known to reflect a highly or terminally differentiated phenotype of epithelial cells, was clearly distinct from the histopathology of the progressively growing xenografts resulting from the injection with nontransduced P-NPC counterparts, or those growing xenografts from the injection with IFN-γ–transduced NPC-BM1 cells or nontransduced NPC-BM1 cells.
The monolayer cultures between P-NPC cells and IFN-γ–transduced P-NPC cells showed quite different growth patterns in that the former cells tended to detach from the bottom of the culture vessels, which coincided with the loss of cell viability once the cells reached confluence, whereas the latter became attached rather firmly throughout the 6-day culture. On the other hand, in nontransduced and IFN-γ–transduced BM-NPC cell cultures, monolayer cells were firmly attached to the bottom of the vessels throughout the log and stationary phases of the growth in both cases. The reason why there was no exhibition of a highly differentiated phenotype in cultured IFN-γ–transduced P-NPC cells is likely that IFN-γ released by the cells was not concentrated enough to induce the differentiation of monolayer cells in vitro, whereas the same cells grown as a three-dimensional mass in vivo were physically more compact, and thus, secreted IFN-γ could readily influence the tumor cells to transform themselves to a much highly differentiated phenotype.
It is of interest that formation of horny pearls and keratin bridges have long been observed with human lung squamous cell carcinoma A431 cells grown in semisolid gelatin containing complete growth medium in the presence of IFN-γ. In such a culture system, three-dimensional growth of tumor cells was allowed to form. 28 This effect was, however, not seen with the same cells grown under the similar culture condition without addition of IFN-γ or with the same cells grown in monolayer culture. These in vitro findings have been interpreted by the authors of these studies as being due to IFN-γ induction of a terminally differentiated phenotype of tumor epithelial cells. This phenotype in turn lost their growth and malignant potential in the three-dimensional architecture of cell growth. 28,29 These early in vitro observations are consistent with the present results from xenotransplantation experiments.
SCID mice were utilized to support the growth of human NPC cells as xenografts, as there are no functional T and B lymphocytes because of a recessive mutation on chromosome 16 in such animals. 30 The immunologic defect is known in the stem cell progenitor pool of T and B lymphocytes. 31 Nevertheless, the macrophages and NK cells are functionally intact in these animals. Because SCID mice will not usually reject xenografts of human tumors or human lymphocytes, the human T and/or B cells can repopulate themselves in SCID mice. SCID mice reconstituted with human effector cells such as lymphokine-activated killer cells, 32 cytokine-induced killer cells, 33 tumor-infiltrating lymphocytes, 34 and/or chimeric antibodies 35 may mimic “human immunocompetent” hosts. In the present study, all animals were sacrificed before they were 4 months old. In the present study, there were no mouse immunoglobulins detected in the sera of 40% randomly selected animals from those injected (data not shown). Thus, it was not likely to have the “leakiness” phenomenon in these animals tested.
The tumor dormancy referred to here is the tumor nodule resulting from the subcutaneous injection in SCID mice of IFN-γ–transduced P-NPC cells, such as NPC076 and NPC039 cells genetically modified to release IFN-γ and IL-6 in greater quantities in vitro. 8 The nodules at the sites of injection remained palpable but their small sizes sustained for the all observation period of nearly 3 months. Histopathologically, these tumor nodules consisted of the structures of horny pearls and keratin bridges as well as some areas of clear cell formation within a papillary-like epithelial tumor lesion, the clear cells signifying degenerative cells. However, not all IFN-γ–transduced P-NPC xenografts entered the differentiation pathway to become the terminally differentiated phenotype, as some of the cells remained viable and could be retrieved from the dormant lesion at the end of the observation period and because of the fact that the processed monodispersed cells could be reestablished as monolayer cultures. The phenotype of these retrieved cells was found to be not much different from that of the IFN-γ–transduced P-NPC cells prior to the first injection to SCID mice (results not shown). Further, in subsequent xenotransplantation experiments, all 4 SCID mice subcutaneously injected with 5×106 retrieved cells also failed to develop progressively growing tumors, and all became dormant tumors at the injection sites. Accordingly, it is reasonable to speculate that CSCs mostly, if not all, must have been inactivated and yet still sustained their minimal levels of stemness in the dormant lesions. The CSCs could be serially transplanted, indicating that it is a self-renewing subpopulation. If the concept of CSCs is right, 2 inactivation through the induction of a high degree of cellular differentiation by approaches such as IFN-γ gene transduction and/or eradication of CSCs by some inhibitors should be important treatment choices for cancer.
The reasons for IFN-γ–transduced BM-NPC cells not susceptible to the induction of tumor dormancy in the xenograft/SCID model used are not immediately clear. However, BM-NPC cells are relatively resistant to be modulated by exposure to IFN-γ protein or upon transduction with the IFN-γ gene (Fig. 1), 8 compared with P-NPC cells when similarly treated. The explanation for this is likely that a greater frequency of dysregulation of the IFN-γ signaling must have occurred along with the IL-6 signaling pathway (STAT-3) in a minority of cells within an NPC primary lesion and a greater number of cells in the metastatic lesion in the background of increasing genetic instability in metastatic tumors as the tumor progressed 26 and that IFN-γ resistance was likely to be associated with attenuation of suppressor of cytokine signaling (SOCS) gene induction and increased metastatic potential. 2,8,36 It has also been well documented that IFN-γ has a potent antiangiogenesis effect. 37,38 Therefore, in the dormant lesions resulting from injection with IFN-γ–transduced P-NPC cells, microvessels were hardly detected. Further, comments should be made on additional features distinguishing P-NPC and BM-NPC cells based on previously documented results as summarized in Table 1. These features include differences in the preferential expression of HLA-DR, greater resistance profile to drugs, such as genectin (G418) 8 and the EGFR inhibitor (cetuximab), 39 greater migration potential and Grp78 expression, 26 etc., by BM-NPC cells. Interestingly, the extent of radiosensitivity between BM-NPC and P-NPC cells seems to be rather similar, whereas the differential sensitivity to IL-6 activation followed by treatment with cetuximab leads to clearly different consequences. 39 Specifically, in sharp contrast to P-NPC cells showing moderate cetuximab-mediated growth inhibition with or without IL-6 pretreatment, BM-NPC cells were more resistance to cetuximab without IL-6 pretreatment and were stimulated with cetuximab following the anti-IL-6 mAb pretreatment. In addition, cell death induced by irradiation was significantly attenuated by IL-6 and augmented by IL-6 antibody. These results imply that blocking IL-6 signaling attenuates aggressive tumor behavior and at the same time can sensitize both BM-NPC and P-NPC cells to irradiation treatment, thus providing a novel and promising treatment approach.
Achieved through transduction of tumor cells with a retrovector carrying the IFN-γ gene.
P-NPCs, cultured cell lines derived from a primary tumor lesion; BM-NPCs, cell lines derived from bone marrow metastasis; Ab, antibody; Grp78, glucose-regulated protein 78; IL, interleukin.
The development of tumor dormancy as the result of subcutaneous injection of these cells in SCID mice is intriguing. It was possible that differentiation of tumor cells induced by either IFN-γ or IL-6, or both cytokines, might have synergistically overrided the angiogenic effects on tumor cell growth. However, the participation of IL-6 is less likely to play a role in tumor dormancy induction, as there are no reports available so far documenting the induction of normal epithelial cells or solid tumor cell differentiation by IL-6. On the other hand, IL-6 has been known to be involved in activation of tumor cells 40,41 or even CSC growth through an autocrine or paracrine fashion. The results of the present study suggest that a malignant state is potentially reversible to a highly differentiated phenotype from some malignancies at the earlier stage. It is indeed much easier to make P-NPC cells rather than BM-NPC cells to be susceptible to induction to a tumor dormancy state. Nevertheless, studies with a large panel of cell lines or cultures from primary or secondary sites of NPC are needed to definitely determine whether the susceptibility of tumor dormancy induction is indeed more restricted for primary NPCs than various metastatic ones including those from organs/tissues other than bone marrow. One should also bear in mind is that the induction of long-term tumor dormancy could be regarded as a form of cancer therapy in a clinical setting. 42,43
Currently, at least five mechanisms have been proposed for the induction of tumor dormancy, namely, cell cycle arrest, hormonal dependence, inhibition of tumor angiogenesis, immune surveillance, and the influence of microenvironment (Fig. 5). 1 –3 Based on results with the human NPC xenograft/SCID model, in this study, the authors propose “induced cellular differentiation” to be the sixth mechanism by which tumor dormancy can be induced through the IFN-γ gene transfer, although cell cycle arrest, 44 antiangiogenesis, 44 and/or stress signaling 45 could have also been involved in the process. From the therapeutic standpoint, it is unethical and not practical to directly infuse IFN-γ–transduced P-NPC cells to the autologous patients. Taking the advantage of the unique features of allogeneic mesenchymal stem cells (MSCs) in their ability to target primary and metastatic tumors and their relatively nonimmunogenic nature, 46,47 such allogeneic MSCs may be used as a novel, universal delivery vehicle for IFN-γ gene to induce tumor dormancy by infusing these cell preparations into preselected patients whose tumors have no detectable or low baseline levels of IFN-γ secretion and yet able to produce high levels of IFN-γ following IFN-γ transduction. This possibility has to be yet first tested in the human NPC-xenograft SCID mouse model used here for the proof of principle.
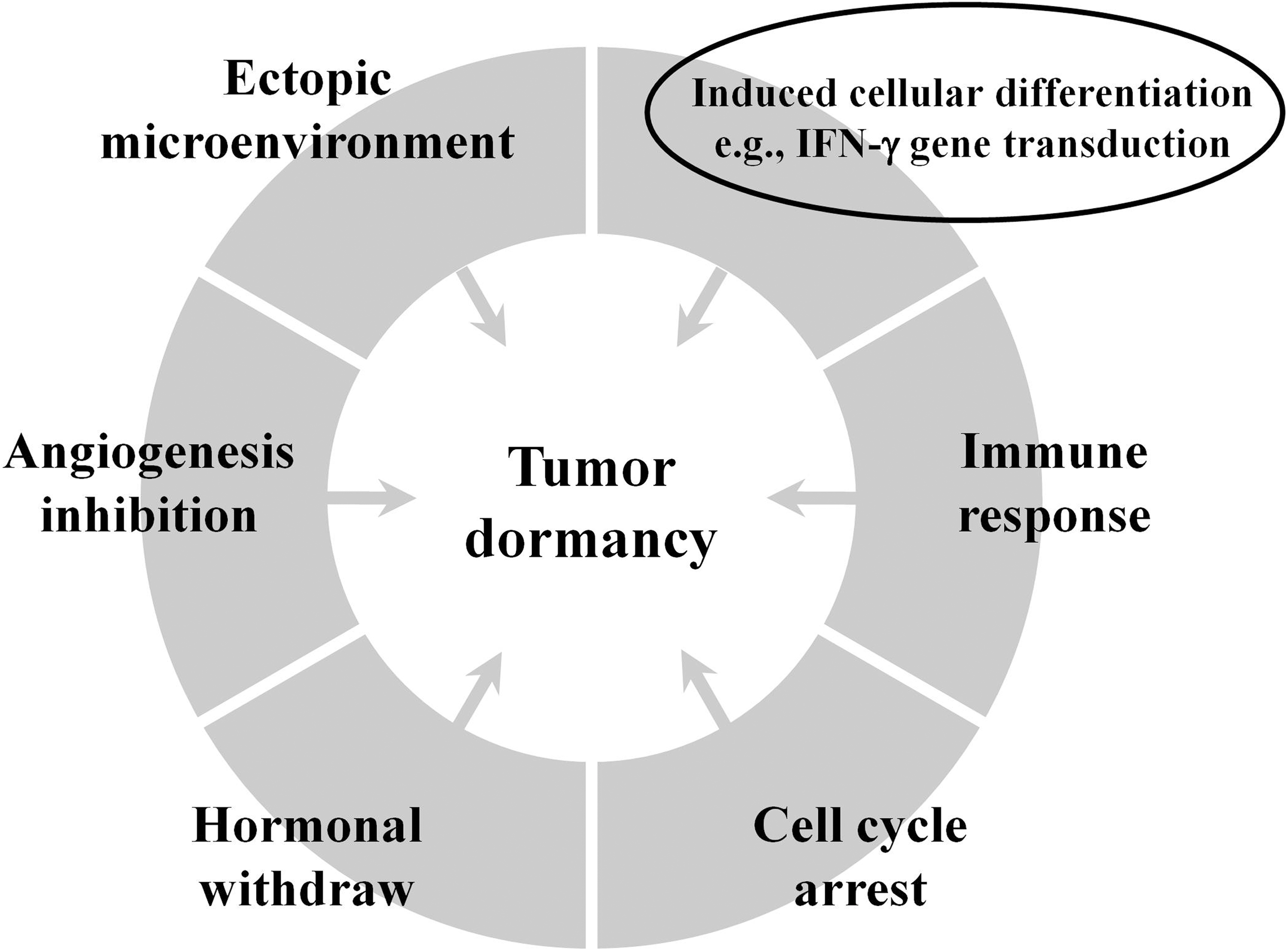
Proposed mechanisms of tumor dormancy. The one highlighted in circle is the mechanism derived from the present study, in which other stated mechanisms, such as cell cycle arrest and angiogenesis inhibition, may, however, also be indirectly or directly involved in the process. This figure was modified from one of the illustrations in Ref. 44
Conclusions
The authors believe that a delicate balance between the induction of a high degree of differentiation/apoptosis/antiangiogenesis, on one hand, and growth stimulation by cytokines/growth factors/proangiogenic or angiogenic factors, on the other hand, must have occurred to allow the tumor to be maintained in a state of dormancy. The resulting information from the present study may also lead to improved treatment strategies for NPC, as induction of tumor dormancy may be considered as one way to treat/control cancer growth 2 and as dormant tumor cells can be considered as therapeutic targets. 42,43 Finally, the present study provides a novel xenograft-SCID mouse model to be manipulated for the induction and even eradication of tumor cell dormancy in the future.
Footnotes
Acknowledgments
The authors express gratitude to Yung-Chi Shen and Ying-Ming Perng for providing technical assistance during the early part of this study and to Prof. Chin-Tarng Lin (Department of Pathology, National Taiwan University, Taipei, Taiwan) for providing the NPC039 and NPC076 cell lines. This work was supported in part by grants from the National Science Council, Taiwan (NSC86-2314-B1B2-091), and the Chang Gung Medical Research Fund (CMRP-624) to S.-K.L.
Disclosure Statement
The authors declare that they have no conflict of interest in relation to the publication of this manuscript.