
Abstract
Efficient delivery of tumor-associated antigens to professional antigen-presenting cells is important for inducing a response in patients receiving cancer immunotherapy. Interferon-gamma (IFN-γ) is used by the immune system to combat viral and fungal infections by restricting cell proliferation and, in some cases, inducing apoptosis. Using IFN-γ to activate target tumor cells prior to antigen loading of dendritic cells (DCs) may enhance the beneficial qualities of whole-cell tumor vaccines. The incubation of melanoma cell cultures with IFN-γ resulted in an increase in the expression of major histocompatibility complex molecules and ICAM-1 but generally decreased the expression of melanoma-associated tumor antigens. Additionally, important immune-stimulating molecules (heat-shock proteins, high-mobility group box-1 protein, and calreticulin) were also present but differentially regulated by IFN-γ. Loading of DCs with IFN-γ-treated tumor cells resulted in a small but significant increase in the expression of CD83-positive DCs, indicating the initiation of DC maturation (p=0.019). IFN-γ treatment of melanoma cell lines prior to antigen loading of DCs may aid in antigen processing and presentation.
Introduction
Melanoma is one of the leading causes of cancer-related death in the United States and is frequently characterized by resistance to chemotherapy. 1 Because melanoma antigens that can be recognized by immune cells have been identified, melanoma is thought to be an ideal candidate for immunotherapy. Dendritic cell (DC)-based therapy modalities may be used to stimulate the immune system in melanoma patients if proper antigen sources can be identified and protocols for generating vaccines are optimized.
Downregulation of tumor-associated antigens and major histocompatibility (MHC) complexes are one of the means that tumor cells use to evade the immune system. Enhancing or increasing the expression of these molecules may boost the immune system when administered as a vaccine. Interferons (IFNs) are pleiotropic cytokines associated with immune response to invading pathogens 2 and have been also shown to regulate the expression of MHC molecules on melanoma cells, which has led to their use as adjuvants for immunotherapy. 3,4 In addition, IFNs can induce apoptosis in some cell types and may also modulate the expression of tumor-associated antigens. 5 –7 As IFNs are used by the immune system as a means to target cells infected by viruses, there may be an advantage to treat tumor cells with IFN prior to their use in the immunotherapy setting, especially in approaches involving antigen-presenting cells such as DCs.
For cell death to be considered immunogenic, certain proteins or factors need to be displayed or secreted by the dying cells, nominally called “DAMPs” or danger-associated molecular patterns. 8 As IFNs can induce cell death and play a role in the immune response to foreign invaders, perhaps tumor cells treated with IFNs will display these molecules as a positive signal to the immune system. An example of a DAMP is calreticulin, a molecule normally found in the endoplasmic reticulum; this molecule has been shown to be translocated to the surface of cells in response to anthracyclines and was found to be critical for immunity against challenge by live tumor cells in a mouse model of colon cancer. 9 Additionally, the high-mobility group box-1 protein (HMGB-1), also known as alarmin, has been shown to be released in response to infection and tissue damage and promotes the maturation of antigen-presenting cells. 10 Last, heat-shock proteins are known to be important regulators of the immune system and also associated with antigen-presenting cell maturation, 11 and although not shown to be regulated by IFNs, their presence may aid in chaperoning tumor-associated peptides to the MHCs on antigen-presenting cells. 12 As IFNs are known to induce cell stress as well as apoptosis, it may be useful to explore whether IFN pretreatment of tumor cells prior to antigen loading results in the induction of positive immune-regulating signals, which may aid in more efficient antigen processing and presentation.
In this study, the effects of treating autologous purified melanoma tumor cells, isolated from surgical specimens and grown as short-term cell lines, with IFN-gamma (IFN-γ) were characterized, and these cells were loaded onto autologous DCs. The combined tumor cells and DCs were then administered as a vaccine in a phase II clinical trial for metastatic melanoma patients along with the adjuvant granulocyte-macrophage colony-stimulating factor (GM-CSF). The results of that trial showed it to be safe and effective with a remarkable 5-year survival rate of more than 50%. 13
Materials and Methods
Autologous DC generation
DCs were generated by plastic adherence method as previously described. 14,15 Briefly, autologous apheresis product was subjected to ficoll-hypaque (GE Healthcare) density gradient separation. The resulting peripheral blood mononuclear cells were placed in antibiotic-free AIM-V medium (Invitrogen) supplemented with 1000 IU/mL each of IL-4 (CellGenix) and GM-CSF (Berlex) (DC medium) at 15×106 cells/mL in cell cultivation flasks (Corning-Costar). After 1 hour incubation, the nonadherent population was discarded and fresh DC medium was added to the flasks. The following morning, the nonadherent cells were discarded, the flasks were washed once with ambient temperature phosphate-buffered saline (PBS), and fresh DC medium was added. The flasks were then cultivated for 6 days at which time flow cytometry evaluation was performed to determine the percentage and phenotype of DCs generated by this approach (preload DCs).
Autologous tumor cell line generation
Pure tumor cells generated and characterized as previously reported 16 –18 were expanded to 200 million cells and then incubated with 1000 IU/mL of IFN-γ (InterMune) for 72 hours in 15% FBS/ECS in RPMI (complete medium), irradiated with 100 Gy from a cesium source, and cryopreserved as previously described. 19 The IFN-γ-treated and irradiated tumor cells were recovered from cryopreservation, washed 3×with PBS, and then added to the in vitro cultivated DCs and incubated for ∼24 hours. The antigen-loaded DCs were harvested by gentle scraping with a rubber policeman and cryopreserved at equal amounts in 9–11 aliquots. An aliquot of cells was obtained for flow cytometry evaluation, which represents the postloaded DCs.
Flow cytometry
Phenotypic characterization of the DC populations were performed using monoclonal antibodies against surface markers obtained from BD Pharmingen: anti-MHC class II conjugated to PerCp, anti-CD11c conjugated to APC, and anti-CD80, anti-CD83, and anti-CD86 conjugated to PE. Isotype controls were used to determine the percentage of positive cells. Flow cytometry of tumor cells was conducted using antibodies against MHC class I and II conjugated to FITC, annexin-V-PE, and 7-amino-actinomycin D (7
Immunoblotting
Cytoplasmic cell lysates were prepared with Mammalian Protein Extraction Reagent (Thermo Scientific) plus protease inhibitor cocktail (Roche) at 10,000 cells/μL on ice. Approximately 25 μL/lane of cell lysates was separated on 12.5% Tris-glycine gels, transferred to PVDF membrane, and probed with antibodies against the following: calreticulin (MBL), Hsp-60, Hsp-70, Hsp-90 (R&D Systems), HMBG-1 (Cell Signaling), ICAM-1 (Santa Cruz Biotech), Mel-4, Mart-1 (Signet), tyrosinase (Upstate), and GADPH (Calbiochem).
Immunohistochemistry
Expression of a panel of antigens by melanoma lines were determined using immunocytochemical procedure. Briefly, cells were cultured in 8-chamber culture slides (Thermo Fisher) in the presence or absence of 1000 IU/mL IFN-γ. After 72 hours, the cells were washed three times with 1× PBS and fixed in cold acetone. After blocking endogenous peroxidase, the cells were incubated with appropriate primary antibodies against the antigens listed. Immunohistochemistry was performed using biotinylated anitmouse or rabbit immunoglobulins, Super Sensitive enzyme-conjugated streptavidin labeling and horse radish peroxidase chromogen, and substrate kits (Biogenex). The reactivity of the following anti-human polyclonal or monoclonal antibodies was investigated with isotype-matched control antibody: S-100 and HMB-45 (Biogenex), Mel-2, Mel-5, Mart-1 (Signet), Tyrosinase, Mage-1 (Thermo Scientific), Melan-A, HLA class I, and HLA class II (Dako).
Statistical analysis
Student's t-test of two-tailed two samples of equal variance was used. Significant differences were determined by a p-value of ≤0.05.
Results
Cell death was differentially induced in the autologous melanoma tumor cell line in response to incubation with IFN-γ for 72 hours in complete medium. Trypan-blue dye exclusion assay performed on cells either treated or not with IFN-γ revealed a significant trend toward lower viability in the IFN-γ-treated cells (89.1%±6.8% vs. 84.9%±9.3%, p=0.014, n=47). Analysis of a sample of four autologous melanoma cell lines by flow cytometry for apoptosis induction (Fig. 1A) revealed that melanoma cells are differentially sensitive to the effects of IFN-γ-induced apoptosis, with some cells displaying more late apoptosis or “dead” populations (7-AAD+/Annexin-V+), whereas others displayed signs of early apoptosis or “dying” populations (7-AAD−/Annexin-V+). The resulting presence of apoptotic cells after IFN-γ treatment was associated with significant decreases in progression-free and overall survival, which was reported elsewhere. 20 In agreement with this published report, a log-rank test revealed a significant association with lower viability upon IFN-γ treatment of melanoma tumor cells and overall survival in patients under study (data not shown).
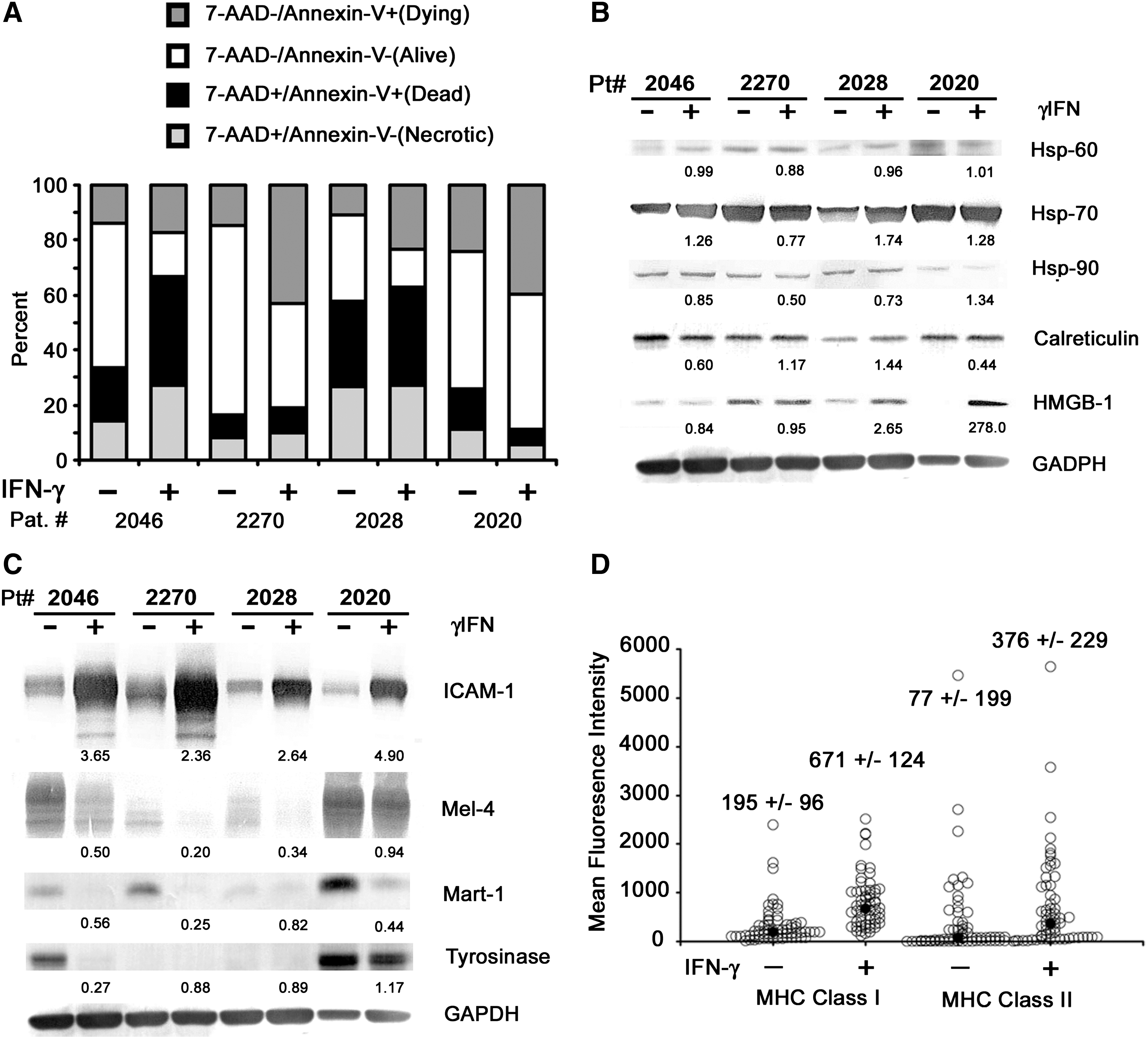
Characterization of interferon-gamma (IFN-γ)-treated autologous tumor cells used for loading dendritic cells (DCs). Autologous melanoma tumor cells were treated with or without 1000 IU/mL IFN-γ for 72 hours in 15% FBS/ECS in RPMI, harvested, irradiated with 100 Gy, and cryopreserved. The cells were then thawed in AIM-V and a sample was taken for flow cytometry and for preparation of cell lysates for immunoblotting prior to antigen loading of DCs.
Lysates from cells that were incubated in the presence or absence of IFN-γ were subjected to immunoblotting for a variety of molecules that may be important mediators of immunity (Fig. 1B). In the setting of melanoma cells treated with IFN-γ, heat-shock proteins appear to be differentially regulated but remain largely present in the cell preparations, especially in the case of hsp-70. The endoplasmic reticulum protein, calreticulin, and HMGB-1 appear to be upregulated in some cases upon treatment with IFN-γ (Fig. 1B). In contrast, common melanoma antigens (mel-4, Mart-1, and tyrosinase) generally appear to be downregulated by IFN-γ, whereas ICAM-1, a lymphocyte adhesion molecule associated with sensitivity to lymphocyte-mediated cytotoxicity, 21 is significantly upregulated (Fig. 1C). Indeed, IFN-γ-treated melanoma tumor cells were found to be more sensitive to cytotoxic T lymphocyte (CTL) activity (data not shown) in agreement with previously published results. 22 Additionally, immunohistochemistry of a panel of melanoma-associated antigens revealed that IFN-γ results in the downregulation of antigen expression in many of the antigens examined (Table 1), confirming observation reported elsewhere. 3
n=27 samples.
IFN-γ, interferon-gamma.
As previously reported, 5 the use of IFN-γ results in the upregulation of the MHCs class I and class II. As shown in Figure 1D, the treatment of autologous melanoma cells with IFN-γ resulted in the near universal and significant upregulation of MHC class I (p=2.8×10−8), with a median fold induction of 2.91±1.13 (95% CI). Additionally, the mean fluorescence intensity of MHC class II was also significantly higher but less so (p=0.039) with a median induction of 4.23±2.66 (95% CI). The level of MHC class II molecules on the surface of the autologous melanoma cells was generally lower than that of the MHC class I molecules, but in 70% of the cases the induction was greater than twofold in response to IFN-γ treatment for the MHC class II molecules because of the low initial level of MHC class II expression. The presence of these molecules on the tumor cells during loading of antigens onto DCs may provide an opportunity for “cross dressing” MHC complexes onto antigen-presenting cells. 23,24
A set of four representative autologous melanoma cell lines were incubated with IFN-γ and loaded in equal amounts onto DCs, which were then assayed by flow cytometry for the expression of CD80, CD83, CD86, and MHC class II. The results indicated that a small but appreciable increase in the percentage of positive population of DCs expressing CD83 was seen upon the loading of the IFN-γ-treated melanoma cells (Fig. 2). Additionally, more unprocessed tumor cells are noted in the CD86 dot plot (upper left quadrant), which resulted in a discernible reduction in the percentage of CD86-positive population, indicating that IFN-γ-untreated tumor cells were still present. This effect is most likely due to the induction of apoptosis by IFN-γ, as apoptotic cells are more likely to be phagocytosed by DCs as previously reported. 20
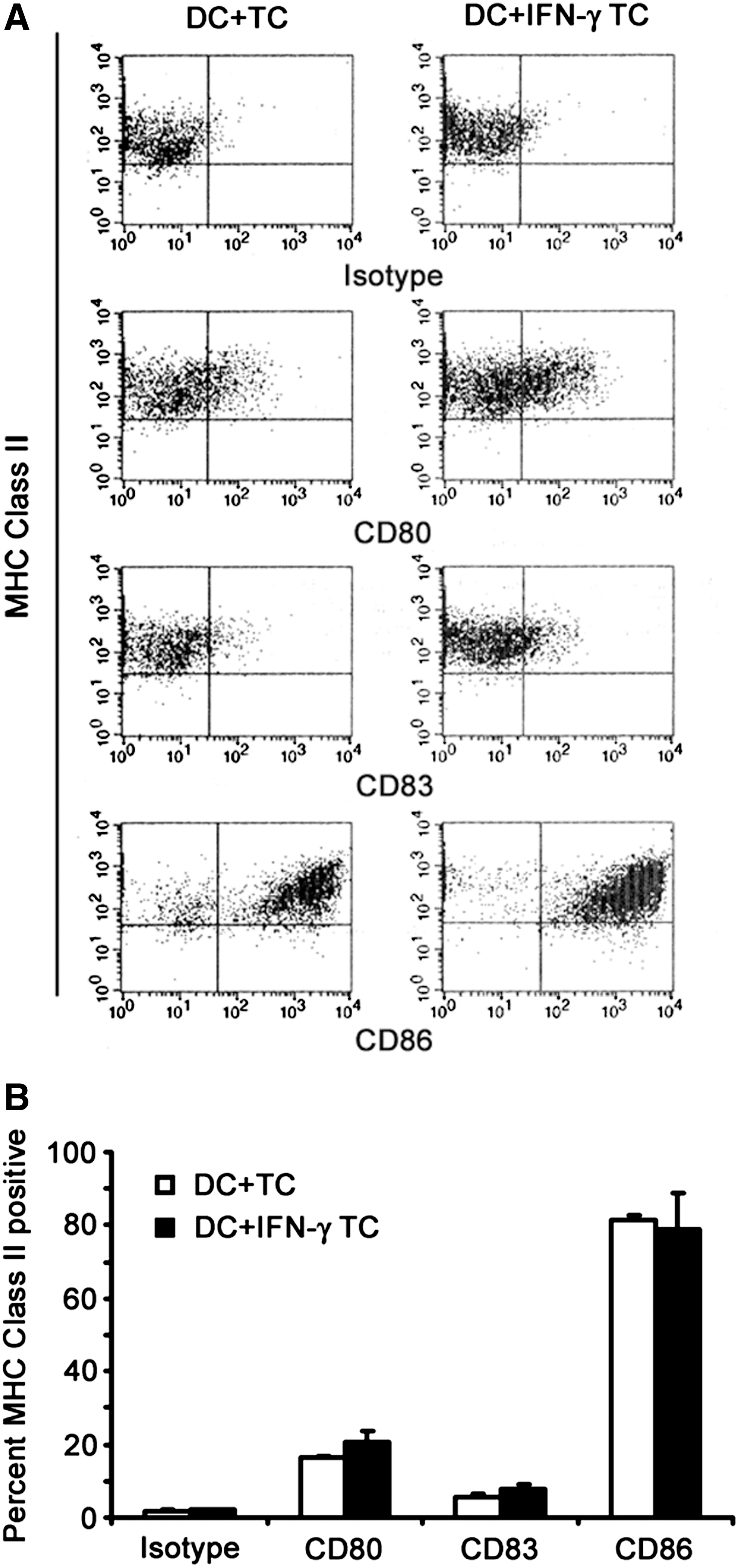
Phenotype of DCs loaded with autologous melanoma cell lines treated with or without IFN-γ. A set of four autologous melanoma cell lines were treated with or without 1000 IU/mL of IFN-γ for 72 hours, irradiated, and cryopreserved. The cells were then thawed in AIM-V and combined with autologous DCs for approximately 24 hours prior to harvest and assaying by flow cytometry for the expression of CD80, CD83, CD86, and MHC class II
As shown in Figure 3, a sample of preloaded DCs showed that they expressed CD80 (39.0%±16.2%), CD83 (7.1%±6.9%), and CD86 (73.6%±19.5%) and were MHC class II positive with a viability of 96.2%±5.0%. The loaded DCs had a significantly higher percentage of CD83 (9.4%±7.1%, p=0.019) with a significantly higher mean fluorescence intensity (172.9±79.0, p=0.0009), indicating that loading the DCs with irradiated, IFN-γ-treated tumor cells induces maturation in some DCs (Fig. 3B).
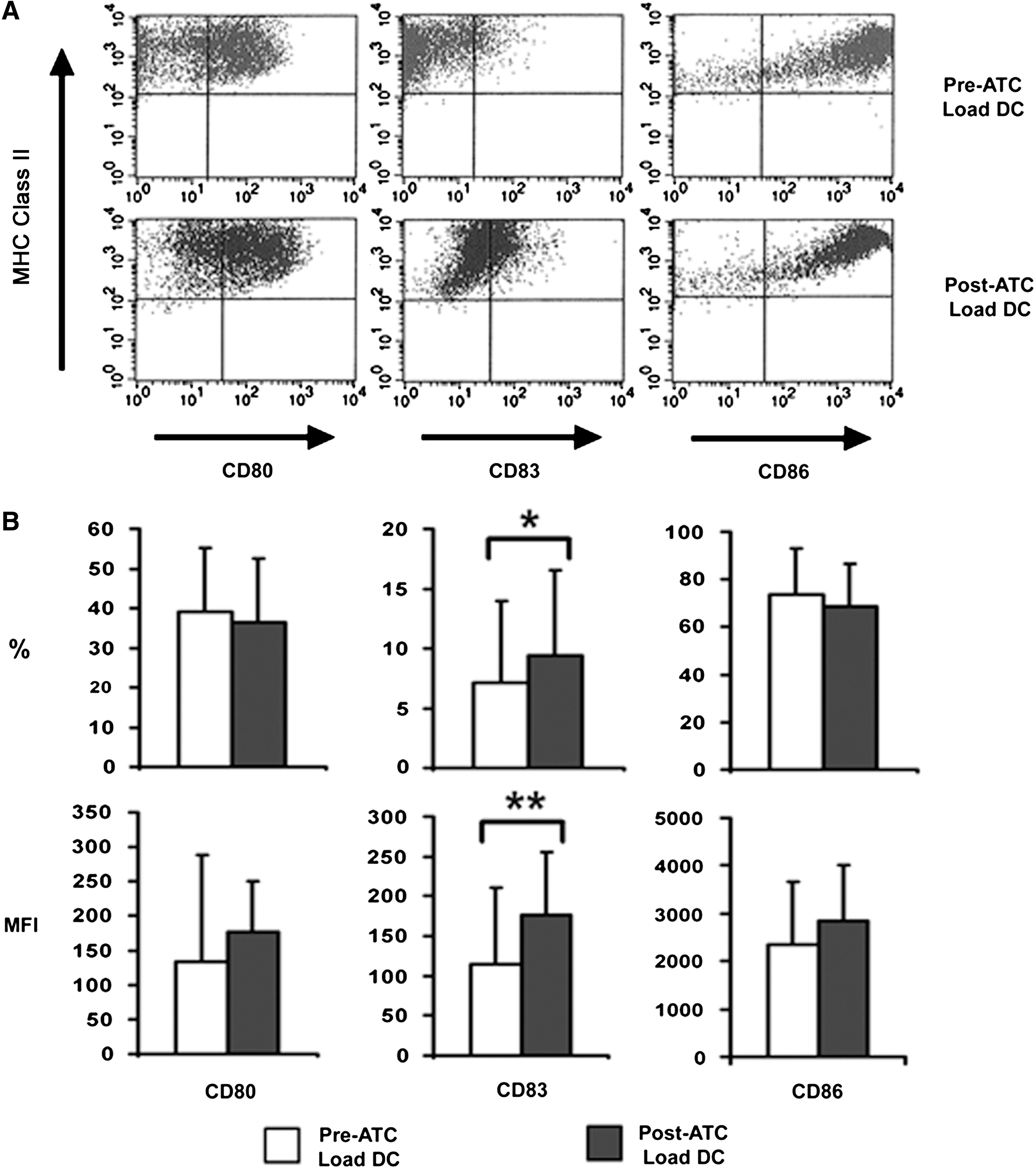
Phenotype of DCs used for dose preparation. Samples of DCs prior to loading (Pre-ATC Load DC, n=53) and after loading (Post-ATC Load DC, n=65) with IFN-γ-treated, irradiated autologous tumor cells were accessed by flow cytometry for the expression of CD80, CD83, CD86, and MHC class II. FACS Caliber beads were used to set the initial flow cytometer instrument settings, which were then held constant throughout the collection of data
Discussion
Improving the efficacy of DC-based cancer immunotherapy continues to be challenging, but optimization of protocols for antigen loading, maturation, and administration routes is currently being evaluated for the most effective methods to induce lasting tumor immunity. 25 Generation of a DC vaccine employing purified autologous tumor cells as the antigen source has been shown to be feasible and perhaps preferable by virtue of containing a patient-specific repertoire of tumor-associated antigens. 18,26 The difficulty and expense of generating purified autologous cell lines may be outweighed by the benefits of having the full complement of tumor-associated antigens in the patient-specific product. Some clinical trials are using unpurified autologous bulk tumors with some success; however, it is not known whether contaminating fibroblasts and/or necrotic tissue affect the immunotherapeutic product. 27 Further, the possibility of tumor stem cell-associated antigens being present in the purified cell lines may be a contributing factor in the relative effectiveness of autologous vaccine therapy. 28
IFN-γ treatment also resulted in a dramatic upregulation of major histocompatibility molecules prior to target cell loading onto DCs. The increased expression of MHC molecules may be a significant benefit, because it was recently demonstrated that the presence of MHC class II molecules is important for response to vaccine therapy. 29 However, in the trial reported here, no correlation could be drawn between overall survival and absolute values of MHC class I or class II expression (data not shown).
In addition, the presence of important immune-regulating molecules present in the phagocytosed material, such as calreticulin, HMGB-1, and heat-shock proteins, may provide a more bona fide maturation signal as opposed to the commonly used cytokine cocktails. Although the DCs present in this vaccine preparation could be considered immature, there is a significant trend toward maturation, which may be associated with the phagocytosis of late-stage apoptotic cells. 30 Additionally, the use of apoptotic cells has been correlated with the generation of DCs that were more effective at stimulating lymphocyte IFN-γ secretion versus DCs loaded with either tumor cell lysates or necrotic cells, suggesting that DCs loaded with apoptotic cells may be more potent in vivo. 31 However, recent published data demonstrated that a resistance to the proapoptotic effects of IFN-γ was associated with a better clinical outcome. 20
Although IL-12 secretion by mature DCs can lead to robust CTL activity, there has been some controversy as to whether ex vivo maturation leads to lasting tumor immunity. 32 The risk of induction of regulatory T cells, which can suppress antigen-specific CTLs, by immature DCs has also been shown to occur with cytokine-matured DCs. 33,34 A reevaluation of the sequence of signaling events that leads to maturation is being investigated to improve DC maturation protocols. 25 Thus, the use of irradiated whole tumor cells as the antigen source in this study, without the necessity of ex vivo cytokine maturation, may be a more preferable method of DC immunotherapy, because the evidence presented here indicates that the DCs have begun the process of maturation. Upon injection, these “maturing” DCs may complete the process of maturation by secreting chemokines, which will attract licensing, antigen-specific CD40L-expressing CD4+ T cells. 35 Indeed, serum chemokines, such as CCL17/TARC, produced by DCs in response to the adjuvant GM-CSF have been associated with better progression-free survival rates. 36
Activation of lymphocytes by DCs requires the expression of costimulatory molecules such as CD80 and CD86. 37 As a marker of maturation, CD83 is expressed on mature DCs and is thought to correspond to DCs that can induce a more potent immune response. 38 Although this represents only a tiny fraction of all the cells in the vaccine preparation, there is no reason to believe that the number of mature DCs alone in any one vaccine regimen leads to a more beneficial response. 39 The use of cytokine-matured DCs remains controversial, so perhaps DCs that are matured in this fashion retain some of the qualities that may be found in more native-born matured DCs because of the signals emanating from the target cells.
Conclusions
The complexity of the immune response and the difficulty in analyzing the results may be the result of the heterogeneity of the patient population. However, further investigation into the correlative factors that indicate which patient subpopulations will benefit from this type of therapy as opposed to more traditional chemotherapeutic methods should yield a more refined approach to the treatment of cancer.
Footnotes
Acknowledgments
The authors thank Sarah Tillman and Andrea Beatty for their assistance in generating the tumor and DC lines, Karen Spencer for her assistance in DC flow cytometry, and Dr. Senthamil R. Selvan for his assistance in evaluating the immunohistochemistry slides. Funding for this work was provided by the Hoag Hospital Foundation.
Disclosure Statement
The authors of this article have no conflicts of interest to disclose.