
Abstract
Objective:
The objective of the present study was to explore the role of the inhibitor of apoptosis protein (IAP) Livin in radioresistance in nonsmall cell lung cancer (NSCLC).
Methods:
Lung adenocarcinoma cell lines A549 and SPC-A1 were used for this study. Using the technique of molecular cloning and gene transfection, two Livin isoforms, Livinα and β, respectively, were expressed in A549 cells with the purpose of exploring the role of Livin in radiation resistance of A549 cells. Moreover, a Livin-specific gene-silencing system was developed using SPC-A1 cell line with the purpose of increasing radiosensitivity of SPC-A1 cells.
Results:
A549 cells were induced by radiation to express Livin isoforms, Livinα and β. A549 cells expressed Livin isoforms stably after gene transfection and the transfected cells demonstrated characteristics of antiradiation. However, Livin gene-silenced SPC-A1 cells exhibited remarkably enhanced radiation sensitivity.
Conclusion:
The IAP Livin is an important molecule in antiradiotherapy of NSCLC. Livin-specific gene silencing is likely to be an effective means to enhance radiation sensitivity of lung cancer.
Introduction
Livin, a novel inhibitor of apoptosis protein (IAP) family member, is very low or undetectable in most normal adult tissues, with the exception of the placenta. 1 Recently, Livin has been found to be highly expressed in most solid tumors, including melanoma, mammary cancer, cancer of the cervix, colon, bladder, stomach, renal, prostate, and esophageal cancers, leukemia, and lymphoma, where its expression is suggested to have clinical significance. 2 –6 Livin expression is significantly correlated with tumor stage, increasing with tumor progression. Both Livin expression rates and levels were higher in patients with bad prognosis. Livin inhibits apoptosis, and its overexpression renders malignant cells resistant to chemotherapy. Therefore, inhibitors of Livin could be useful adjuncts to chemotherapy in the treatment of malignancies. Several studies suggest that sequence-specific siRNA targeting Livin is capable of enhancing apoptosis of human cancer cells, such as breast cancer, malignant melanoma, hepatocellular carcinoma, and renal cell carcinoma. 7 –13 Some studies have reported Livin expression in nonsmall cell lung cancer (NSCLC). Tanabe 14 showed that the positivity rate for Livin mRNA expression in NSCLC was 76.3% (29 of 38). Knockdown of Livin inhibits cell growth and induces apoptosis in NSCLC cells, increasing cell chemosensitivity. 15
As already known, NSCLC usually shows radioresistance. Is there any relationship between radioresistance in NSCLC and IAP family member Livin? On the other hand, Livin has two isoforms, Livinα and β, two splicing variants that are almost identical, except for 54 bp. What is the physiological or pathological significance of the coexistence of the two isoforms? So far, whether Livin isoforms are involved in radioresistance of lung cancer cells is still a to-be-answered question. In the present study, experiments showed that A549 and SPC-A1, two lung adenocarcinoma cell lines, differ in Livin expression: Livin is not detected in A549, but both Livin isoforms are highly expressed in SPC-A1. In this study, eukaryotic expression vectors for Livin were constructed and then transfected into A549 cells to research on the role of Livin in radioresistance of A549 cells. Then, Livin expression was specifically downregulated in SPC-A1 cells to research on radiosensitizing effect on lung adenocarcinoma cells. This study provided experimental basis for research on mechanism of Livin's involvement in radioresistance of lung cancer cells.
Materials and Methods
Construction of recombinant plasmid pcDNA3.1-Livin
First, the software of Primer Premier 5.0 was used to design specific polymerase chain reaction (PCR) primers (Livinα-1 and β-1) for full-length cDNA of Livinα and β. Recognition sites for HindIII and XbaI (TaKaRa) were added at each 5′ end, respectively. Second, total RNA was isolated from SPC-A1 cells using TriPure isolation reagent (Roche). Full-length cDNA of Livinα and β was obtained by RT-PCR (TaKaRa RNA LA PCR™ Kit Ver.1.1; TaKaRa). PCRs using primers Livinα-1 and β-1 to get the whole Livin isoform cDNAs were run at annealing temperatures of 60°C for 30 cycles. Recombinant clones were validated by enzyme digestion and secondary PCRs using primers Livinα-2 and β-2 at annealing temperatures of 56°C for 30 cycles. Positive recombinant clones (pcDNA3.1-Livinα and pcDNA3.1-Livinβ) were validated again by sequencing. All the primers and length of PCR products were as follows: Livinα-1, 5′-cgc
Stable expression of Livin isoforms in A549 cells
Electroporation was performed for gene transfection. The cells were passaged at 1/10 dilution after 48 hours. G418 was added (800 μg/mL) for A549 cell screening. The culture medium was replaced at 2–3-day intervals until G418-resistant clones emerged (3 weeks after plating). Resistant cells were cloned by limiting dilution. The cells were kept under G418 selection. Morphology of A549 cells was assessed before and after transfection under inverted microscope and transmission electron microscope.
RT-PCR and northern blot
Livin isoform mRNA expression was detected by RT-PCR and northern blot in G418-resistant A549 cells. Primers of Livinα-2 and β-2 were used in RT-PCR. GAPDH (5′-accacagtccatgccatcac-3′, 5′-tccaccaccctgttgctgta-3′, 452 bp) was used as internal control. Total RNA from SPC-A1 cells was used as a positive control. Total RNA from A549 cells transfected with empty vector (pcDNA3.1) was used as a negative control. Thirty micrograms (30 μg) of total RNA was used in northern blot. The blot was hybridized at 68°C overnight in a hybridization solution (6×SSC + 0.5% SDS + 100 μg/mL denatured salmon sperm DNA) containing 32P-labeled DNA probe. Last, the membrane was washed and subjected to autoradiography at −70°C for 48 hours.
Immunofluorescence cytochemistry
Passaged cells were cultured on sterilized coverslips in a six-well plate at a density of 1×105 cells/mL. Immunofluorescence cytochemistry assay was performed at 1–2 days after the cells adhered to the coverslip. About 1:500 diluted rabbit anti-human Livin antibody (Alpha Diagnostic International, Inc.) was added as first antibody, incubated at 37°C for 60 minutes, and then incubated at 4°C overnight. A 1/500 diluted FITC-labeled secondary antibody (Beijing Zhongshan Biotechnology Co., Ltd.) was then added at a concentration of 50 μL per coverslip. The coverslip was sealed with 50% glycerin and fluorescence microscope was used for observation and photography.
Radiation-induced A549 cells apoptosis and Livin expression
A549 cells in log-phase growth before and after transfection were dissociated using 0.25% trypsin and subcultured for 24 hours in IMDM supplemented with 10% calf serum. A549 cells were radiated by 2 Gy 60Co before and after Livin isoforms transfection. After 48 hours, genomic DNA was isolated using phenol–chloroform method. Eight microliters (8 μL) of DNA products were electrophoresed on 2% agarose gels in 1×TAE buffer, stained with ethidium bromide, and visualized under UV light. Apoptotic ladder was observed. Otherwise, Livin isoforms in wild-type A549 cells were detected by RT-PCR at 24, 48, 72, and 96 hours after 2 Gy γ-ray radiation.
Radioresistance of Livin isoform-overexpressing A549 cells
A549 cells were divided into the following three groups: group A549/pcDNA3.1 (control group, A549 cells transfected with pcDNA3.1), group A549/Livinα (A549 cells transfected with pcDNA3.1-Livinα), and group A549/Livinβ (A549 cells transfected with pcDNA3.1-Livinβ). Attached A549 cells in exponential growth were subjected to digestion, resuspended in IMDM containing 10% calf serum to get single cell suspension, and seeded in 10-mL dishes at a cell density of 104 cells/10 mL. Then, A549 cells were irradiated with 60Co γ-rays at doses of 0, 2, 4, 6, 8, and 10 Gy and cultured for 2–3 weeks. The culture medium was replaced with a fresh one every 2 days. The number of clones containing >50 cells was counted under the microscope. The colony forming rate (%) and survival fraction (SF=colony forming ratio of radiation/colony forming ratio of nonradiation) were calculated. GRAPHPAD Prism 4.0 was used to draw radiation survival curve of A549 cells in different groups. D0, Dq, and N values were gained.
Gene silencing of Livin in SPC-A1 cells
Livin isoform-specific siRNA sequence screening was performed by selecting siRNA sequence in the common and different sequences of Livinα and Livinβ cDNAs. Livin isoform-specific siRNA oligonucleotide was synthesized and inserted into pSilencer2.0 vector (Ambion) and pAVU6+27 (kindly gifted from Dr. David Engelke, Michigan University) after being ligated with adaptors. The sequences of siRNA were as follows: Livinα, 5′-gtgcccaggagccaggaggggtc-3′; Livinβ, 5′-gtgcccaggagccaggagccagg-3′; Livinα+β, 5′-gacagtgccaagtg cctgc-3′, 5′-ggtgaggtgcttcttctgc-3′. At the same time, GAPDH siRNA (5′-gtggatattgttgccatca-3′) and nonhomologous sequence siRNA (5′-actaccgttgttataggtg-3′) afforded by pSilencer2.0 kit were used as positive and negative control, respectively. Molecular cloning was programmed as usual. After sequencing, positive recombinant plasmid was cotransfected with pIRES-EGFP at a 5:1 ratio into SPC-A1 cells. G418 selection and gene transfection by electroporation were performed as earlier. Resistant clones were picked up in 400 μg/mL G418 and cultured. Transfection effect was observed under the fluorescence microscope.
Semiquantitative RT-PCR and western blot
RT-PCR was programmed as described earlier. Grayscale value of every band was analyzed using a Gel-Doc 2000 gel analysis system (BioRad). The ratio of Livin/GAPDH (A Livin/GAPDH) of the bands in the same sample was calculated to quantilize mRNA expression of Livin isoform. In western blot, rabbit anti-human polyclonal anti-Livin antibody (Alpha Diagnostic International, Inc.) or goat anti-human polyclonal anti-GAPDH antibody (1/200; Santa Cruz) was added as first antibody, incubating for 60 minutes at room temperature. Horseradish peroxidase-labeled goat anti-rabbit (Beijing Zhongshan Biotechnology Co., Ltd.) or mouse anti-goat antibody (1/200) as second antibody was added to each sample, incubating for 60 minutes at room temperature. Then, protein bands were visualized by DAB. Grayscale value of every band was analyzed using a Mias image analysis system (Si Chuan University, China). The ratio of Livin/GAPDH (A Livin/GAPDH) of the bands in same sample was calculated to quantilize mRNA expression of Livin isoform.
Radiation sensitivity of Livin isoform-specific gene silencing in SPC-A1 cells
SPC-A1 cells were divided into four groups: Group SPC-A1/pSilencer2.0 (control, SPC-A1 transfected by pSilencer2.0-U6), Group SPC-A1/siLivinα (SPC-A1 transfected by pSilencer-Livinα), Group SPC-A1/siLivinβ (SPC-A1 transfected by pSilencer-Livinβ), and Group SPC-A1/siLivinα+β (SPC-A1 transfected by pSilencer-siLivinα+β). Attached SPC-A1 cells in exponential growth were subjected to digestion, resuspended in IMDM containing 10% calf serum to get a single-cell suspension, and seeded in 10-mL dishes at a cell density of 104 cells/10 mL. Then, SPC-A1 cells were irradiated with 60Co γ rays at doses of 0, 2, 4, 6, 8, and 10 Gy and cultured for 2–3 weeks. Colony formation, survival curve, and D0, Dq, and N values were gained as described earlier.
Statistical analysis
Each experiment was repeated three times. Data were expressed as the mean reading±standard deviation and analyzed using the software SPSS13.0. The significance of the differences between two means was evaluated by t-test. Difference was considered to be significant if the p-value was below 0.05.
Results
Stable expression of Livin isoforms in A549 cells
Eukaryotic expression vectors for Livin were constructed and then transfected into A549 cells by electroporation to research on the radiosensitizing effect of Livin on A549 cell growth. To ensure relatively high purity and expression rate of cells for the following experiment, monoclonal screening method is effective and reliable. In this study, limit dilution was performed for culture and amplification of monoclonal cells. Single clones in the test group were picked up for amplification, culture passaged, and then frozen for future use. After transfection, no special morphological changes of cells were observed under invert microscope. However, the cells seemed to be more polytypic and more microvillus under transmission electron microscope (Fig. 1A, B).
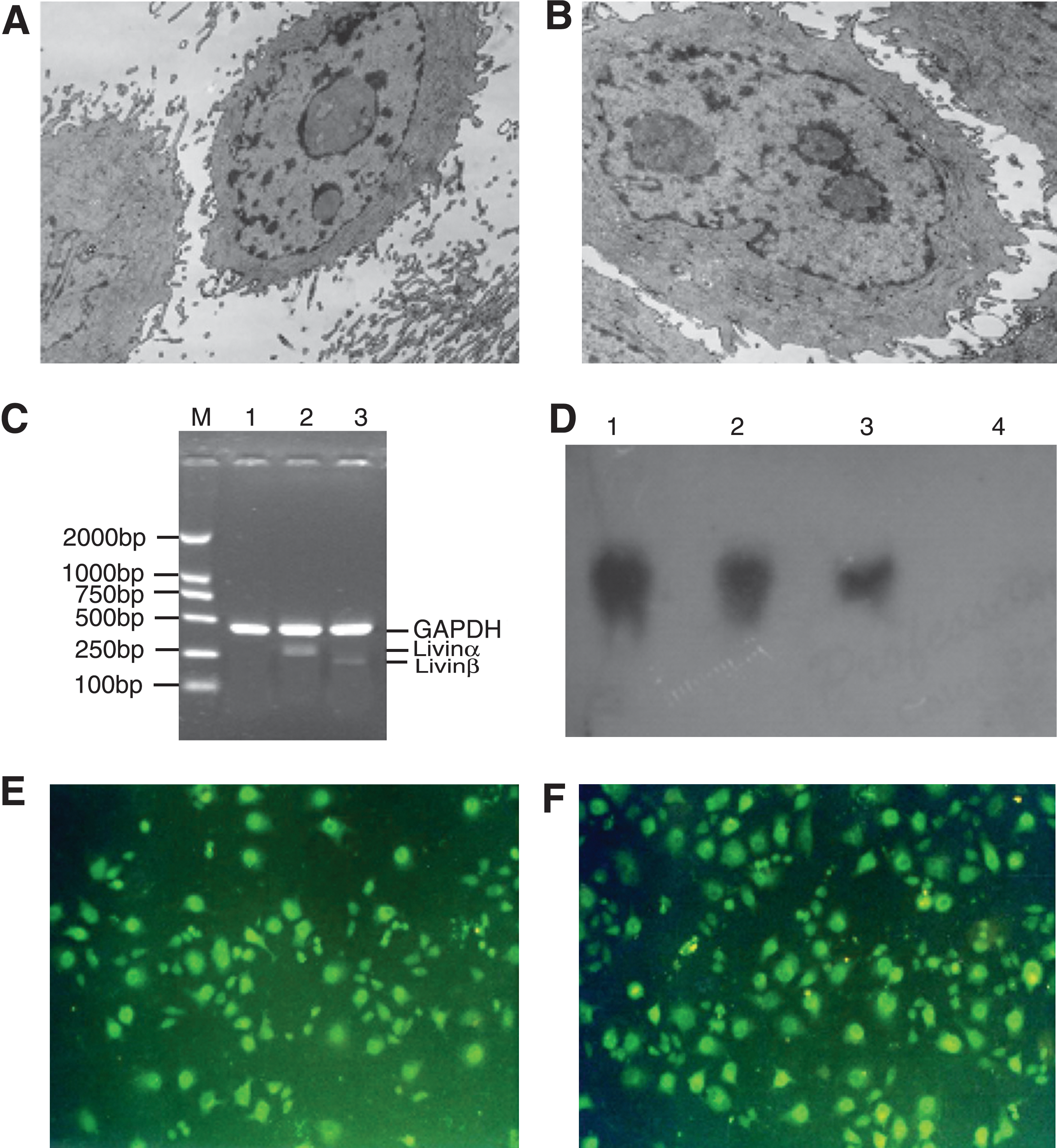
A549 cells before and after Livin gene transfection. After transfection, no special morphological change was observed under inverted microscope. However, more polytypes and more microvilli were observed under electron microscope
The amount of Livin mRNA was measured by RT-PCR and northern blot. Total RNA was extracted from transfected cells for RT-PCR. As a result, two fragments (275 and 221 bp, respectively) were successfully amplified in pcDNA3.1-Livin–transfected A549 cells but not in pcDNA3.1-transfected A549 cells, demonstrating a successful Livin expression in A549 cells (Fig. 1C). The transfected cells were collected to extract total RNA for northern blot. pcDNA3.1-Livin–transfected A549 cells, as well as positive control (SPC-A1), had positive results. However, pcDNA3.1-transfected A549 cells had negative results, further demonstrating the successful expression of Livin in A549 cells (Fig. 1D).
Livin protein expression was validated by immunofluorescence chemistry. Being activated by bluish violet light from the microscope, cells gave rise to yellowish green fluorescence under fluorescence microscope (Fig. 1E, F). So did SPC-A1 cells, whereas no fluorescence was observed in wild-type A549 cells (data not shown). Taken together, the data suggested that isoforms (Livinα and β) were expressed in A549 cells.
Radiation-induced apoptosis and Livin expression in A549 cells
At 48 hours after being irradiated at 2 Gy by 60Co γ-rays, A549 cell genome DNA was extracted. Electrophoresis showed apoptotic change of 180 bp DNA ladder in the control group (A549/pcDNA3.1), but no apoptotic change of DNA ladder in the A549/Livinα or A549/Livinβ group (Fig. 2A). Moreover, Livinα and β expression in irradiated A549 cells were detected. Livin isoforms began to appear at 24 hours after radiation, became higher at 48 and 72 hours, and also could be seen at 96 hours (Fig. 2B).
Radiation-induced apoptosis and Livin expression in A549 cells.
Specific gene silencing of Livin isoforms
Livin isoform-specific siRNA expression vectors were recombinated, namely pSilencer-Livinα, pSilencer-Livinβ, pSilencer-siLivinα+β, and pAVU6+27-siLivinα+β. The results of evaluation and sequencing showed that Livin isoform-specific siRNA expression vector was cloned successfully, and continuous siRNA expression in cells was achieved by gene transfection. The results of semiquantitative RT-PCR and western blot showed that the inhibition rate of Livin expression is 60%–80%, relating to different target. About 54 bp sequence difference between two isoforms makes correspondent siRNA gene knockdown highly efficient and specific (Table 1 and Fig. 3). A Livin/GAPDH showed grayscale value of one specific Livin isoform band relative to that of GAPDH band. Inhibition efficiency of pSilencer-siLivinα+β was stronger than that of pAVU6+27-siLivinα+β. So pSilencer-siLivinα+β was only chosen for the following study.

Livin expression in SPC-A1 cells before and after transfection. The results of semiquantitative RT-PCR
The results of semiquantitative RT-PCR and western Blot showed that inhibition rate of Livin expression is 60%–80%, relating to different target. About 54 bp sequence difference between two isoforms makes correspondent siRNA gene knockdown highly efficient and specific. A Livin/GAPDH showed grayscale value of one specific Livin isoform band relative to that of GAPDH band. Inhibition efficiency of pSilencer-siLivinα+β was stronger than that of pAVU6+27-siLivinα+β.
Compared with SPC-A1 or SPC-A1/pSilencer-Control, p<0.01.
PCR, polymerase chain reaction.
Antiradiation effect of A549 cells overexpressing Livin isoform
Radiation survival curves of A549 cells (Fig. 4A) included the control (A549/pcDNA3.1), A549/Livinα, and A549/Livinβ groups. The three groups had different D0, Dq, and N values. In the control (A549/pcDNA3.1), A549/Livinα, and A549/Livinβ groups, D0 values were 2.545, 2.703, and 2.632, Dq values were 1.956, 2.412, and 2.095, and N values were 3.062, 3.968, and 3.826 (Table 2). There were significant difference between the A549/Livinβ and A549/pcDNA3.1 groups (p<0.05) and very significant difference between the A549/Livinα and A549/pcDNA3.1 groups (p<0.01). D0, Dq, and N values increased at different degrees in A549 cells after Livin isoform was overexpressed. The results showed that Livin isoforms had similar tolerance to γ-rays at different degrees. Generally speaking, Livinα surpassed Livinβ in resisting γ-rays.
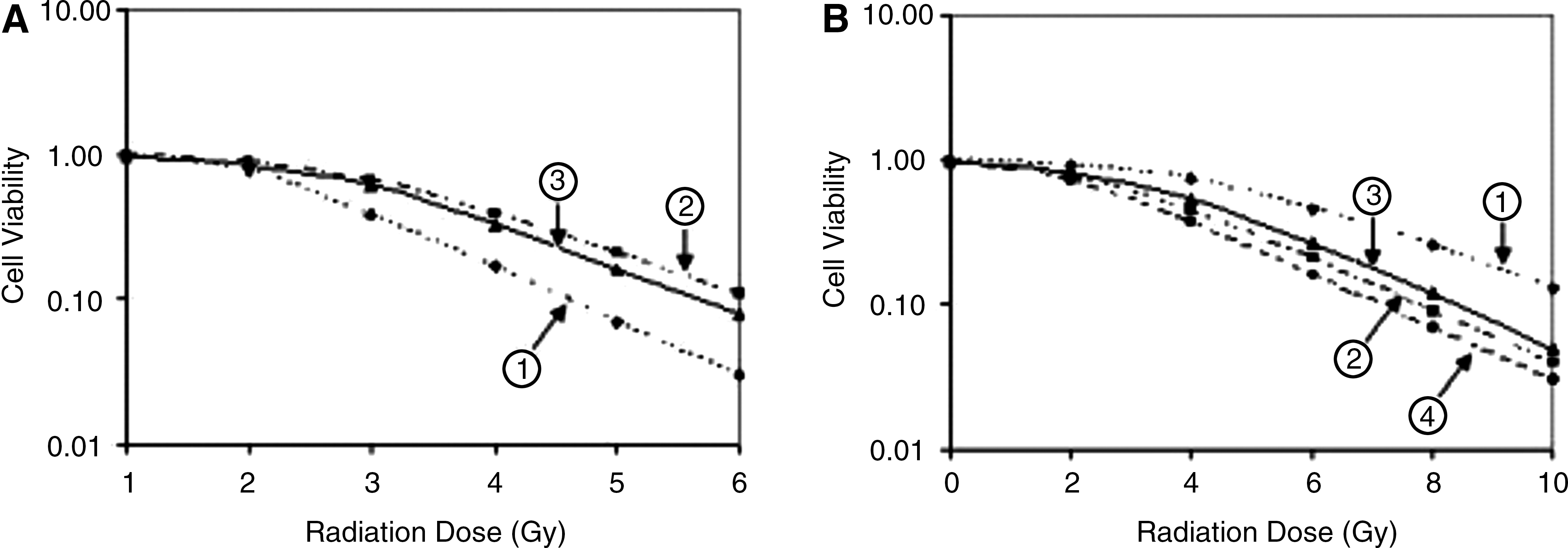
Effect of Livin isoforms on radioresistance of nonsmall cell lung cancer.
Comparing with A549/pcDNA3.1 or SPC-A1/pSilencer2.0, a p<0.01, b p<0.05.
D0, Dq, and N values increased at different degrees in A549 cells after Livin isoform overexpressed. Livin isoform had tolerance to γ-rays. D0, Dq, and N values decreased at different degree in SPC-A1 cells after Livin isoform knockdown. Livin isoform gene silencing had radiosensitization effect in γ-rays treatment.
Radiation sensitivity of Livin isoform-specific gene silencing in SPC-A1 cells
Radiation survival curves of SPC-A1 cells (Fig. 4B) included the control (SPC-A1/pSilencer2.0), SPC-A1/siLivinα, SPC-A1/siLivinβ, and SPC-A1/siLivinα+β groups. Four groups had different D0, Dq, and N values. In the control (SPC-A1/pSilencer2.0), SPC-A1/siLivinα, SPC-A1/siLivinβ, and SPC-A1/siLivinα+β groups, D0 values were 3.371, 2.486, 2.748, and 2.343, Dq values 2.916, 2.167, 2.512, and 1.875, and N values 3.021, 2.553, 2.832, and 2.431 (Table 2). Statistical analysis showed that, compared with the SPC-A1/pSilencer2.0 group, D0, Dq, and N values of the SPC-A1/siLivinα, SPC-A1/siLivinβ, and SPC-A1/siLivinα+β groups are significantly different (p<0.01, 0.05, and 0.01, respectively). D0, Dq, and N values decreased at different degree in SPC-A1 cells after Livin isoform knockdown. The results showed that Livin isoform gene silencing had similar (but different degree of) radiosensitization effect in γ-ray treatment. Knockdown of Livinα+β showed highest sensitivity to γ-rays.
Discussion
Research shows that damage of apoptotic mechanism in malignant tumor cells is closely related to the genesis, development, and recession of tumor, which is also an important reason for tumor chemo/radioresistance. IAP family is characterized by one or more repeats of a highly conserved ∼70 amino acid domain termed the baculoviral IAP repeat (BIR) and suppresses apoptosis triggered by a wide variety of stimuli, including viral infection, chemotherapeutic drugs, staurosporin, and growth factor withdrawal, and by components of the tumor necrosis factor-α (TNF-α)/Fas apoptotic signaling pathways. 16 –18 There are currently 5 human IAP family members, namely c-IAP1, c-IAP2, XIAP, NAIP, and Survivin. 19 –22 Inhibiting expression of IAP family member to induce apoptosis in tumor cells is a promising research direction in apoptosis-inducing tumor treatments, which may play an important role in reversing the drug resistance of tumor and be an adjunctive treatment after operation, chemotherapy, or radiotherapy so as to decrease tumor metastasis rate and recurrence rate.
Research on IAP family, especially research on relationship between Survivin expression in various tumor tissues and tissue typing, tumor typing, and tumor prognosis, has been a hot research field. 23 –25 Livin, a novel IAP family member, has a single BIR domain and a COOH-terminal RING finger domain. It shares similar structure (i.e., containing a single BIR domain) and similar tissue expression profile with Survivin. However, they differ not only in antiapoptotic function, but also in tissue expression profile: Livin, not Survivin, is a negative prognostic marker in early-stage superficial bladder cancer and may also be a marker for bladder cancer recurrence, demonstrating that, compared with other IVP family members, Livin has a different antiapoptosis mechanism.
To investigate Livin's effect on tumor progress and to search for new target for tumor treatment, it is important to research on Livin expression in different tumors and its biological function. Like other IAP members, Livin has been shown to interact with specific cysteine proteases, or caspases, which is required for the cleavage of certain proteins involved in the disassembly of the cell during apoptosis. But Livin has two isoforms (Livinα and β), which are different splice variants from the same gene. Research on Livin becomes a hot research field. Why does the cell express two proteins that have little difference in structure? What is the physiological or pathological significance of coexistence of the two isoforms? What is the difference between these two isoforms with regard to their antiapoptosis effect? What techniques can be used to effectively inhibit one isoform with the purpose of researching on function of the other isoform? Can either isoform be used as a molecular target for apoptosis-inducing tumor therapy? All questions are worth exploring.
There are a few reports of the relationship between Livin expression and lung carcinoma. Crnkovic-Mertens et al. 26 have observed that the targeted inhibition of Livin strongly sensitized NSCLC cells to different proapoptotic stimuli, such as UV irradiation or the chemotherapeutic drug etoposide. In addition, long-term silencing of Livin blocked the growth of NSCLC cells in colony formation assay. These effects of small interfering RNA were specific for Livin-expressing tumor cells. In a previous study, two lung carcinoma cell lines (A549 and SPC-A1) were used for RT-PCR assay. As a result, A549 cells do not express Livin variants. But after being irradiated, A549 cells expressed both Livin isoforms from 24 to 96 hours. Livin gene was transfected into A549 cells. Survival curves showed that D0, Dq, and N values increased. Livin isoforms had tolerance for γ-rays. SPC-A1 highly expressed two Livin variants. Moreover, when Livin genes were silenced in SPC-A1 cells, survival curves showed that D0, Dq, and N values decreased. Livin isoforms gene silencing enhanced cell sensitivity to γ-rays. The present study indicates that Livin is an important contributor to the radiation resistance of NSCLC cells and may serve as a novel molecular target for NSCLC therapy.
Livin has two isoforms, Livinα and β, which are different splicing variants from the same gene. Full-length cDNA of Livinα covers 897 bp, whereas that of Livinβ covers 843 bp. The only sequence difference between the isoforms is the 54 bp deletion of Livinβ in exon 6. However, they differ not only in antiapoptosis function, but in tissue expression profile. 27 Livinα, not Livinβ, could be a marker for poor prognosis in superficial bladder cancer. Overexpression of Livinα stimulated cell proliferation and inhibited chemical-induced apoptosis in bladder cancer cells. Livinα expression increases during the progression of bladder cancer. It may be used as a marker of early recurrence. 28,29 So far, differences between antiapoptosis functions of Livin isoforms remain unclear; the mechanism of Livin's involvement in antiapoptosis of lung cancer cells remains unknown. Whether Livin is involved in radiotherapy resistance of lung cancer cells is still a to-be-answered question. The relevant research may be a direction of this work. In the present study, Livin isoform-specific siRNA was efficiently and specifically expressed in SPC-A1 cells and could be a powerful tool for research on different functions of the two isoforms. Livinα, Livinβ, and Livinα+β can all serve as targets for radiation sensitivity-enhancing treatment of lung cancer (Livinα+β surpasses Livinα; Livinα is better than Livinβ). This study provides a basis for further exploring different antiapoptosis functions of Livin isoforms and also an experimental basis for exploring their role in radiotherapy resistance of lung cancer cells.
Crnkovic-Mertens 30 also used an isoform-specific RNA interference (RNAi) strategy to silence endogenous Livin expression in HeLa cells. The targeted inhibition of Livin beta, but not of Livin alpha, blocked the growth of HeLa cells in clonogenic survival assays. In addition, silencing of Livin beta, but not of Livin alpha, sensitized HeLa cells to different proapoptotic stimuli such as UV irradiation, TNF-α, or etoposide. However, the present study showed that both Livinα and β in A549 cells have similar radiation tolerance for γ-rays. Livinα surpasses Livinβ in resisting γ-rays. Gene silencing showed the same results in SPC-A1 cells. Livin isoform gene silencing had similar (but different degree of) radiosensitization effect in γ-rays treatment. Knockdown of Livinα in SPC-A1 cells showed remarkably higher sensitivity to γ-rays than knockdown of Livinβ. Knockdown of Livinα+β showed highest sensitivity to γ-rays. So, Livinα and Livinβ are likely to have different antiapoptosis roles in lung cancer cells to resist proapoptotic stimuli such as chemicaltherapy drugs or radiation.
This study demonstrated that Livin expression increases remarkably after radiotherapy and showed that besides killing tumor cells, radiotherapy also leads to Livin synthesis and expression in tumor cells as a result of apoptosis tolerance, which may partially contribute to radioresistance of lung cancer cells. Being useful in research on functions of isoforms with small sequence difference, RNAi can specifically inhibit expression of Livin isoforms and is thus an ideal technology in research on functions of two Livin isoforms. The next experimental stage should be to further test the radiosensitization of malignant cells in tumor-engrafted animals. Livin isoform could be a radiosensitizer in lung cancer radiotherapy.
Footnotes
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (to J.-G.S.; no. 30200282), the National High Technology R&D Program (to Z.-T.C.; no. 2008AA02Z104), and the Youth Innovation Foundation of Third Military Medical University (to R.-X.L.; no. 2007D172). The siRNA vector pAVU6+27 was kindly presented by Dr. David Engelke from Michigan University.
Disclosure Statement
No competing financial interests exist.