
Abstract
The nucleotide sequence encoding amino acids 1–30 of endostatin (peptide 30, with amino acids 25–31 mutated from RGIRGAD to RGDRGD) was artificially synthesized and cloned into the plasmid pTYB2 and expressed in Escherichia coli (DE3). Peptide 30 was purified by chitin affinity chromatography followed by dithiothreitol removal by gel filtration and protein identification using Tricine–sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Similarly, peptide 27, corresponding to amino acids 1–27 of endostatin, was produced as control. The effects of peptide 30 on the metastatic potential of HepG2 cells were then investigated. Peptide 30 was found to effectively suppress the adhesion, invasion, and migration of HepG2 cells. Flow cytometry demonstrated that peptide 30 did not alter the expression of membrane integrin αvβ3, although immunofluorescent staining revealed that these integrins formed clusters on the cell surface. Therefore, the effects of peptide 30 on cancer cell invasion may involve the αvβ3 pathway. Moreover, peptide 30 inactivated metalloproteinase-2 (MMP-2) and MMP-9 and downregulated the expression of COX-2 (cyclooxygenase 2), MMP-2, and MMP-9 at both mRNA and protein levels. Peptide 30 also upregulated the expression of tissue inhibitor of metalloproteinase-1 and tissue inhibitor of metalloproteinase-2 at mRNA and protein levels. Lastly, an antibody against αvβ3 enhanced the biological effects of peptide 30.
Introduction
Endostatin is an endogenous angiogenesis inhibitor discovered by O'Reilly 1 in 1997. It is composed of 184 residues at the C-terminus of gelatin XVIII and is able to suppress cancer cell growth by inhibiting angiogenesis in tumors, 2 which has encouraged its clinical use.
In 2005, Tjin Tham Sjin et al. 3 found that amino acids 1–27 at the N-terminus of endostatin played an active role in suppressing angiogenesis. Two (2) years later, the authors' group modified engineered bacteria carrying the endostatin coding sequence by site-specific mutation. More specifically, the RGIRGAD sequence, corresponding to amino acids 25–31, was mutated to RGDRGD, producing an RGD-linked, 30-amino acid peptide (peptide 30). In vitro and in vivo experiments demonstrated that this peptide was more potent than endostatin in suppressing angiogenesis and cancer cell growth. 4
The present study evaluated the effects of peptide 30 on invasion by and migration of the hepatocarcinoma cell line HepG2. The present findings form a basis for the future clinical application of peptide 30.
Materials and Methods
Reagents
RPMI-1640 cell culture medium (Invitrogen), fetal calf serum (PAA), and trypsin (DIFCO) were purchased from Invitrogen, and matrigel (E1270) and Sephadex G-15 medium (Pharmacia) were purchased from Sigma. Fibronectin (FN) was obtained from Sigma.
WST-1 reagent was obtained from Roche. A chitin affinity chromatography column was purchased from New England Biolabs. Antibodies against α1, α2, α3, α5, β1, αv, β3, αvβ3, metalloproteinase-2 (MMP-2), tissue inhibitor of metalloproteinases-1 (TIMP-1), TIMP-2, COX-2, and β-actin and all secondary antibodies were purchased from Santa Cruz, and anti-MMP-9 was obtained from Cell Signaling Technology. Transwell cell culture chambers were purchased from Corning Costar. The total RNA extraction kit was purchased from Promega, and the DRR019ART-PCR kit was purchased from TaKaRa.
Cell culture
The human hepatocarcinoma cell line HepG2, purchased from the Shanghai Institute of Cell Biology, was cultured in RPMI-1640 culture medium supplemented with 10% fetal calf serum and incubated in 5% CO2 at 37°C. The cells were maintained on plates and subcultured after 0.25% trypsin digestion. All experiments were performed after the cells reached 70% confluence.
Extraction and purification of peptide 30 and peptide 27
Peptide 30 and peptide 27 were extracted and purified from the engineered bacteria pTYB2-T30 and pTYB2-T27 encoding peptide 30 and peptide 27, respectively, as described by Wang. 5 Dithiothreitol was removed using a Sephadex G-15 affinity column, and the purity of the peptides was confirmed by Tricine–sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE).
Effects of peptide 30 on growth and viability of HepG2 cells
HepG2 cells were adjusted to 1×104 cells/mL, and 100 μL of this cell suspension was added to each well of a 96-well plate. The cells were maintained for 24 hours before addition of peptide 30 or peptide 27 at 0, 20, 40, 60, 80, or 100 μg/mL. The cells were then grown for another 24 or 48 hours, and then 100 μL of serum-free medium and 10 μL of WST-1 (highly water-soluble tetrazolium salt, 2-(4-iodophenyl)-3-(4-nitrophenyl); Roche) reagent were added to the wells. Absorbance was determined using a plate reader (Bio-Rad) at 450 nm, with 630 nm as a reference.
Effects of peptide 30 on adhesion of HepG2 cells
As described by Sara et al. 6 and Chen et al., 7 96-well plates were coated with 2 μg of matrigel overnight at 4°C. Next, 20 μL of 2% BSA was added to each well for 1 hour, followed by washing with PBS. Meanwhile, HepG2 cells were treated with peptide 30 or peptide 27 at 0, 20, 40, 60, 80, or 100 μg/mL for 24 hours. The plates were then cultured at 37°C for 90 minutes. MTT (5 mg/mL) was added to each well at 20 μL per well, followed by incubation at 37°C for 4 hours. The reaction was terminated by adding DMSO to the medium. The spectrophotometer absorbance at 570 nm was measured.
Effects of peptide 30 on migration of HepG2 cells (scratch-wound healing assay)
HepG2 cells were seeded onto six-well plates without peptide treatment until the cells grew into a confluent single layer. The treatment groups received 50 μg/mL peptide 30 or peptide 27 for 24 hours, whereas the control group received no treatment. The culture medium was then discarded, and an identical line was scraped into the single layer of cells for each group, with three wells per treatment. 8 The width of the line was recorded and photographed at 0, 24, and 48 hours after wounding.
Effects of peptide 30 on invasion by HepG2 cells
As described by Liu et al., 9 the outer side of the polycarbonate membrane of a transwell was coated with 5 μg of fibronectin, and the membrane's inner side was coated with 5 μg of matrigel and air-dried to form a matrix layer. For treatment groups, the cells were treated with peptide 30 or peptide 27 for 24 hours before digestion with trypsin. The cell suspension was specifically dispensed into the transwell chambers, which were fit into the wells of 24-well plates, at 1×105 cells/100 μL. The wells of the plates also each contained 500 μL of RPMI-1640 medium supplemented with 20% FBS. After 24 hours of incubation, the cells on the lower side of the transwell were removed using a cotton stick. The transwell membrane was then stained with 0.1% crystal violet for 20 minutes. The membrane was photographed, and the number of cells that had migrated to the lower side of the membrane was counted under a microscope (TS100; Nikon) at 100× magnification.
Effects of integrins on invasion by HepG2 cells
The procedure for studying the effects of integrins was the same as that described above in “Effects of peptide 30 on invasion by HepG2 cells”. Cells treated with 50 μg/mL peptide 30 for 24 hours were digested with trypsin. Eight wells of a transwell chamber were then filled with 100 μL of cell suspension containing 2×105 cells. Antibodies against integrin α1, α2, α3, α5, β1, αv, β3, or αvβ3 were then added to the wells to a final concentration of 50 μg/mL. Cells treated only with antibody against peptide 30 or αvβ3 were included as controls.
Effects of peptide 30 on clustering of integrin αvβ3
HepG2 cells growing in log phase were digested with 0.25% trypsin and seeded onto 24-well plates at 5×105 cells per well for 24 hours. The treatment groups were then treated with 50 μg/mL peptide 30 or peptide 27, and the control group was treated with the same volume of PBS for 24 hours. Next, the cells were washed twice with PBS and fixed in 4% paraformaldehyde for 20 minutes at 4°C. After 30 minutes of blocking with 5% BSA, the cells were incubated with anti-αvβ3 (1:100) at 4°C overnight. Then, the cells were incubated with FITC-conjugated secondary antibody (1:500) at 37°C for 1 hour, followed by three PBS washes and fluorescent microscopy.
Flow cytometry assay of integrin αvβ3 expression after peptide treatment
This experiment followed the procedure described by Qin et al. 10 The cells were treated with 50 μg/mL peptide 30 or peptide 27 or left untreated for 24 hours. The cells were then digested with 0.25% trypsin and incubated with anti-αvβ3 at 37°C for 30 minutes. Then, the cells were dispensed at 1×105 cells/tube and incubated with FITC-conjugated secondary antibody at 37°C for 30 minutes. After further washing with PBS, the cells were detected by flow cytometry using a FACS Calibur (BD Biosciences).
Gelatin zymography assay for MMP-2 and MMP-9 activity after peptide 30 treatment
The same number of HepG2 cells (1.0×106 per well) were seeded onto six-well plates and cultured at 37°C overnight. The cells were treated with peptide 30 in serum-free RPMI-1640 at 0, 20, 40, 60, 80, or 100 μg/mL for 24 hours. The conditioned medium was collected and loaded onto a nonreducing 10% SDS-PAGE gel with 0.1% gelatin. 11 After electrophoresis, the gel was washed three times in 2.5% Triton X-100 at room temperature for 15 minutes. The gel was then incubated in gelatinase buffer at 37°C overnight, fixed, and stained in fixation buffer containing 0.25% Coomassie Brilliant Blue, followed by destaining.
Additional control, peptide 27-treated, and peptide 30-treated samples were also analyzed in parallel, with the control cells receiving 500 μL of serum-free RPMI-1640. The treatment groups received peptide 30 or peptide 27 at 50 μg/mL for 24 hours. The supernatants were then collected and analyzed by gelatin zymography.
Determination of expression of integrins αv and β3, COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 after peptide 30 treatment by RT-PCR
The total RNA was extracted using an extraction kit (Promega) after treatment of the HepG2 cells with 50 μg/mL peptide 30 or peptide 27 for 0, 12, 24, or 48 hours. The primers used in the PCR amplification are listed in Table 1, and the cycling conditions were as follows: predenaturing at 94°C for 2 minutes, followed by 35 cycles of denaturation for 30 seconds, annealing for 30 seconds, and extension at 72°C for 60 seconds. β-Actin, used as an internal control, was amplified for 30 cycles. The PCR products were visualized by electrophoresis in 1.5% agarose containing 0.5 μg/mL ethidium bromide under ultraviolet light.
MMP-2, metalloproteinase-2; TIMP-1, tissue inhibitor of metalloproteinase-1.
The HepG2 cells in another set of three treatment groups were administered 50 μg/mL peptide 30, 50 μg/mL αvβ3 antibody, or 50 μg/mL peptide 30 plus 50 μg/mL αvβ3 antibody, respectively, for 24 hours. Then, the mRNA levels of COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 were detected by RT-PCR as earlier.
Determination of expression of integrins αv and β3, COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 after peptide 30 treatment by western blotting
HepG2 cells were first treated with peptide 30 or peptide 27 at 50 μg/mL for 0, 12, 24, or 48 hours. The cells were lysed in 100 μL of RIPA buffer on ice for 1 hour. After the samples were centrifuged at 4°C at 16,000 rpm for 10 minutes, the supernatant was collected, and total protein concentration was measured using a nucleic acid and protein detector (DU640; Beckman). Fifty (50) micrograms of protein per sample was then used to perform 10%–15% SDS-PAGE. Following the transfer of the protein onto a PVDF membrane (Millipore) and incubation for 2 hours in 5% bovine serum albumin (BSA; Sigma), the membrane was incubated with primary antibodies against integrins αv, β3, COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 overnight. The next day, the membrane was washed and incubated with HRP-conjugated secondary antibody at 1:5000 for 2 hours. The membranes were developed by a standard enhanced chemiluminescence (ECL) method based on the manufacturer's protocol (Amersham).
The HepG2 cells in another set of three treatment groups were administered 50 μg/mL peptide 30, 50 μg/mL αvβ3 antibody, or 50 μg/mL peptide 30 plus 50 μg/mL αvβ3 antibody, respectively, for 24 hours. Then, the protein levels of COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 were detected by western blotting as earlier.
Statistical analysis
All results are expressed as the mean±standard deviation (SD). The data were analyzed by one-way analysis of variance (ANOVA), followed by either Dunnett's t-test for separate comparisons with the control group or Sidak's t-test for multiple-group comparisons. Differences were considered significant at p<0.05.
Results
Expression and purification of peptide 30
Peptide 30 and peptide 27 were purified by chitin affinity chromatography. These recombinant peptides resulted in a specific 3-kDa band visualized by Tricine–SDS-PAGE (Fig. 1).

Purified peptide 30 and peptide 27 visualization by Tris–Tricine–sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Lane 1: protein molecular weight marker; lane 2: recombinant peptide 30 (5 μg); lane 3: recombinant peptide 30 (2 μg); lane 4: recombinant peptide 27 (5 μg).
Peptide 30 inhibits proliferation of HepG2 cells
Peptide 30 or peptide 27 was added to the HepG2 cells' supernatant at various concentrations (0–100 μg/mL) for 24 or 48 hours. Cell growth was inhibited in a dose-dependent manner, with peptide 30 observed to be more active than peptide 27 (Fig. 2). The IC50 of peptide 30 was calculated based on a linear regression model to be 49.7 μg/mL. This value was used in subsequent studies.
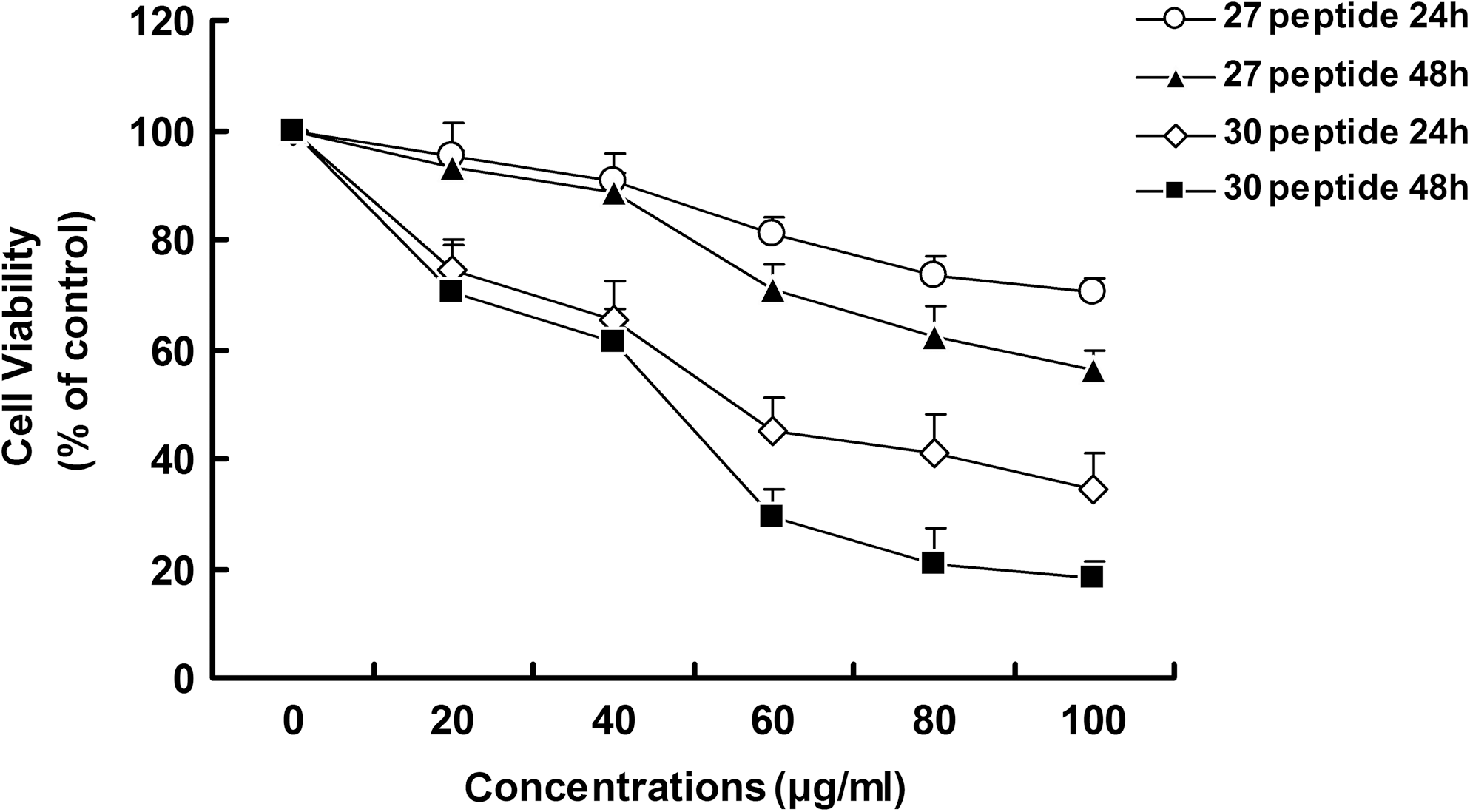
Effect of peptide 30 on HepG2 cell viability. HepG2 cells were seeded onto 96-well plates at 5×103 cells per well and treated with different concentrations (0–100 μg/mL) of peptide 30 or peptide 27 for 24 or 48 hours. Each concentration was repeated in five wells (n=5). Cell viability values, determined using the WST-1 assay, were expressed relative to the control group (0 μg/mL dose). Each result exhibited here is the mean of at least three independent experiments.
Peptide 30 suppresses adhesion of HepG2 cells to the extracellular matrix
In contrast to peptide 27, peptide 30 significantly suppressed the adhesion of HepG2 cells at a concentration of 40–100 μg/mL (p<0.05) (Fig. 3).
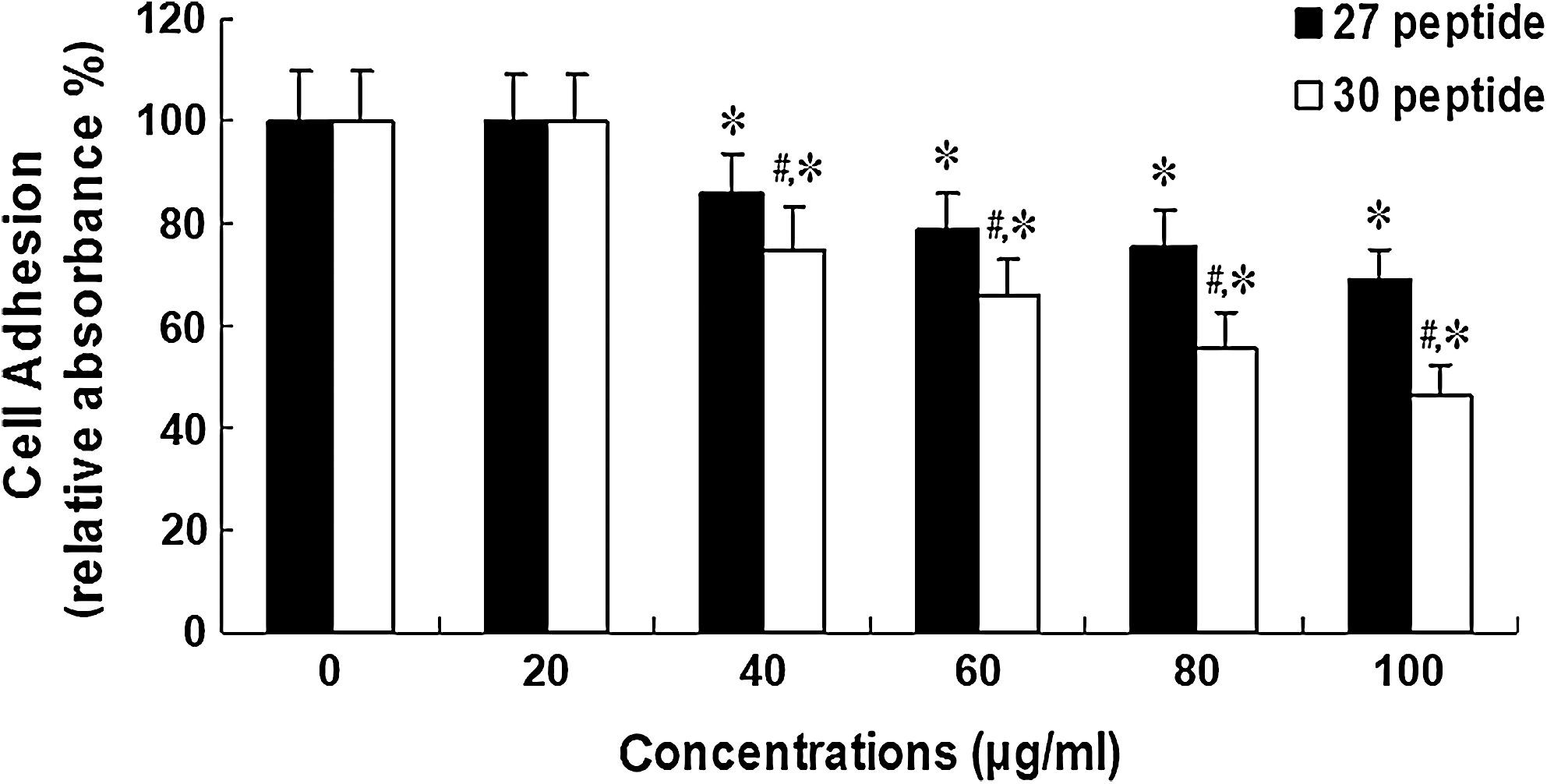
Concentration-dependent effects of peptide 30 on cell-matrix adhesion of HepG2 cells. HepG2 cells were treated with various concentrations (0–100 μg/mL) of peptide 30 or peptide 27 for 24 hours. Treated cells were then subjected to cell-matrix adhesion, as described in the Materials and Methods section. The experiments were repeated three times (n=3), and values are expressed as the mean±SD. *p<0.05, compared with the control group (0 μg/mL dose); # p<0.05, compared with the peptide 27-treated group.
Peptide 30 suppresses migration of HepG2 cells
A wound-healing assay was designed to assess the effects of peptide 30 on the migration of HepG2 cells. In the control group, the cells grew to confluence by 48 hours after wounding, whereas both peptide 27 and peptide 30 suppressed cell migration at 24 hours after wounding. At 48 hours after wounding, peptide 30 more significantly suppressed cell migration than peptide 27 (Fig. 4).
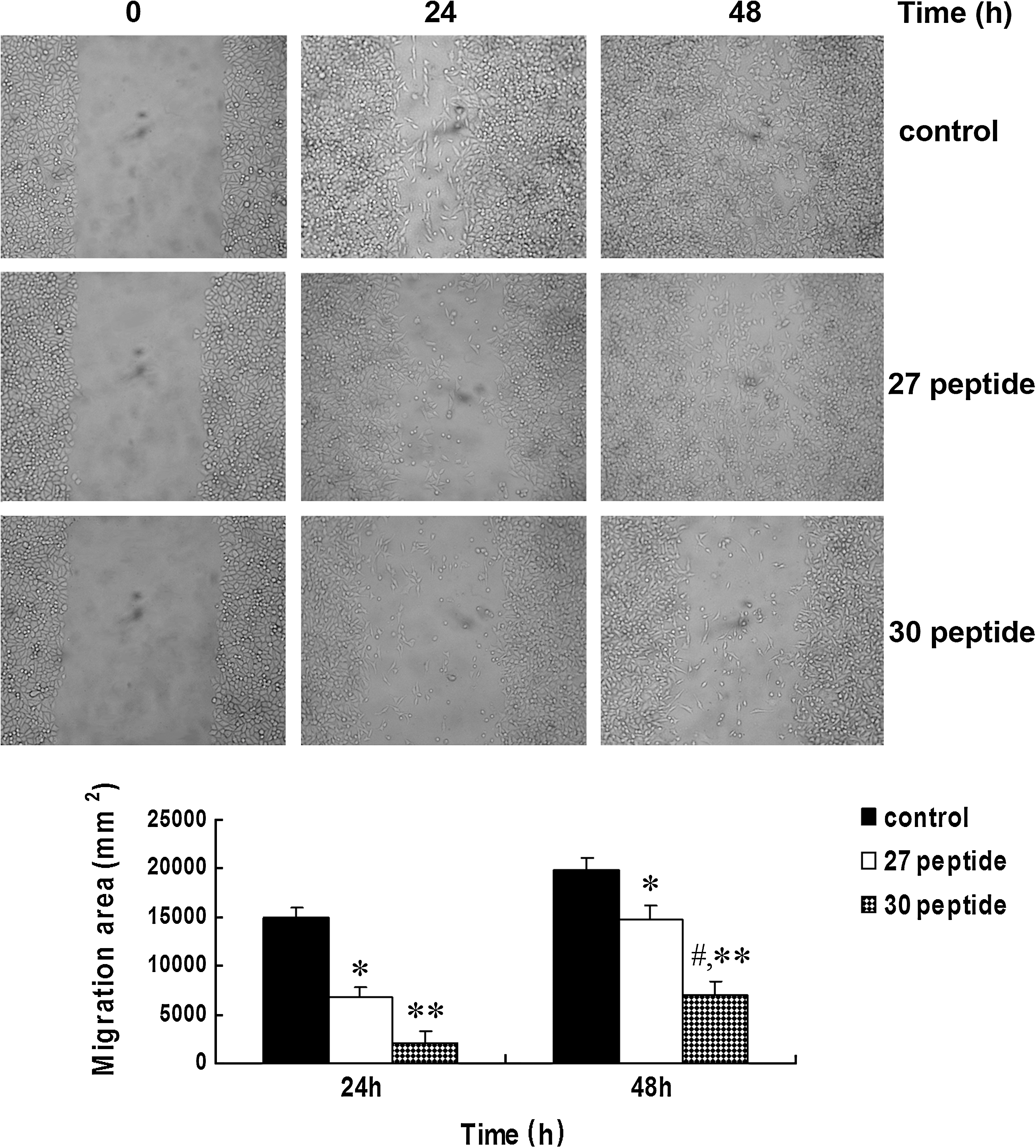
Effect of peptide 30 on HepG2 cell migration. The migration of HepG2 cells was measured by means of a wound-healing assay. HepG2 cells were treated with 50 μg/mL peptide 30 or peptide 27 for 0, 24, or 48 hours. A wound track was scored in each dish using a plastic scraper, followed by incubation for 48 hours. The data represent the mean±SD. *p<0.05 and **p<0.01, compared with the control group; # p<0.05, compared with the peptide 27-treated group. The experiments were performed in triplicate (n=3).
Peptide 30 suppresses invasion by HepG2 cells
Figure 5A shows the numbers of cells invading the other side of the transwell membrane: 205.6±4.6, 132.9±3.1, and 71.4±2.8 for the control, peptide 27-treated, and peptide 30-treated groups, respectively. To further explore the mechanism of peptide 30 suppression of HepG2 cell invasion, antibodies blocking integrin α1, α2, α3, α5, β1, αv, β3, or αvβ3 were added to the transwell chambers previously treated with peptide 30; the resultant numbers of migrating cells were 90.5±3.5, 86.8±4.6, 82.7±5.0, 79.2±3.8, 82.5±5.8, 39.99±4.5, 74.3±3.9, and 38.9±6.1, respectively. Antibodies against αv and αvβ3 further suppressed HepG2 cell invasion (Fig. 5B).
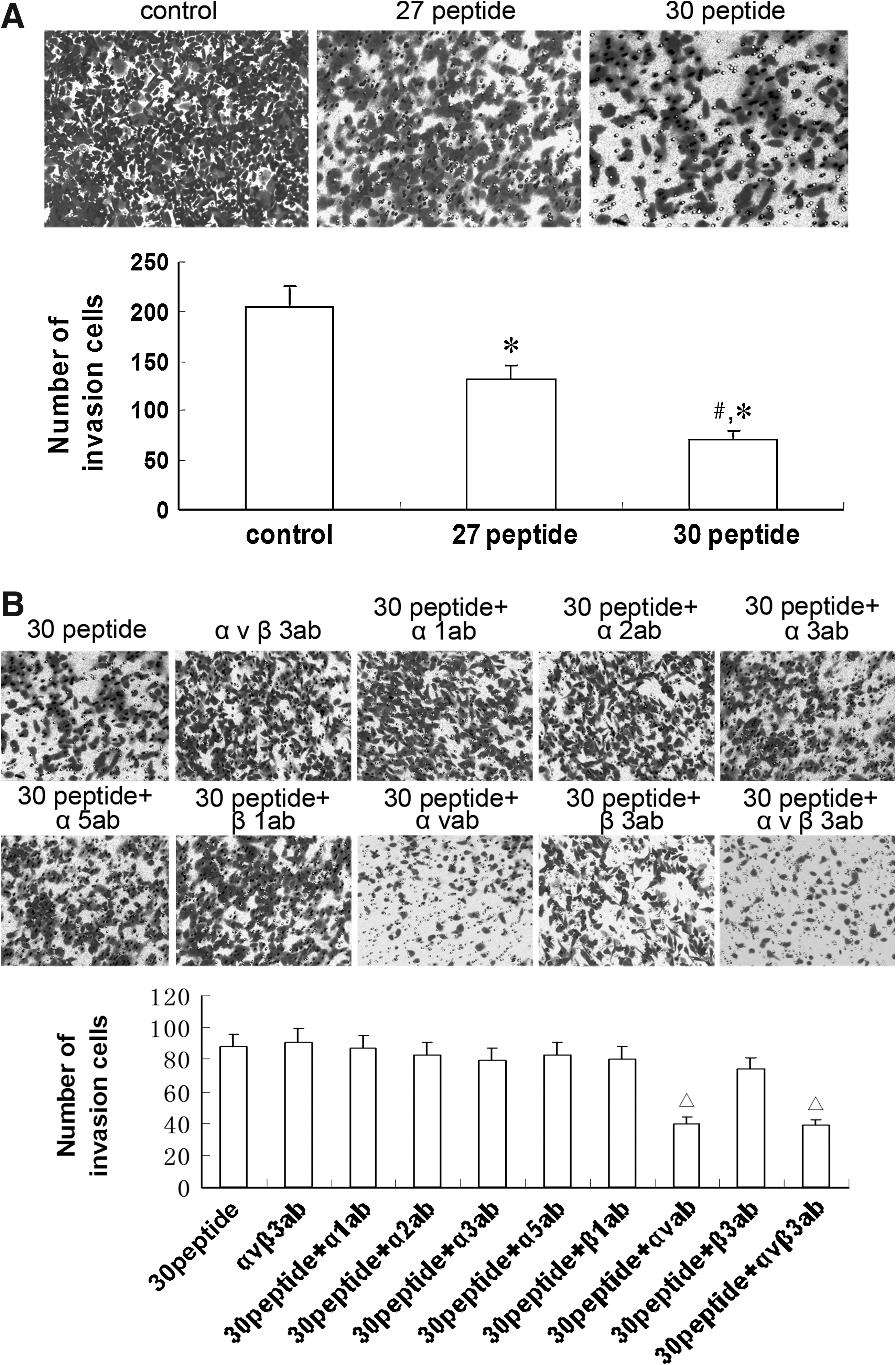
Effects of peptide 30 and integrins on invasion by HepG2 cells.
Peptide 30 causes clustering of integrin αvβ3
Compared with the control, the peptides, especially peptide 30, induced significant clustering of integrin αvβ3 on the HepG2 cell surface (Fig. 6).
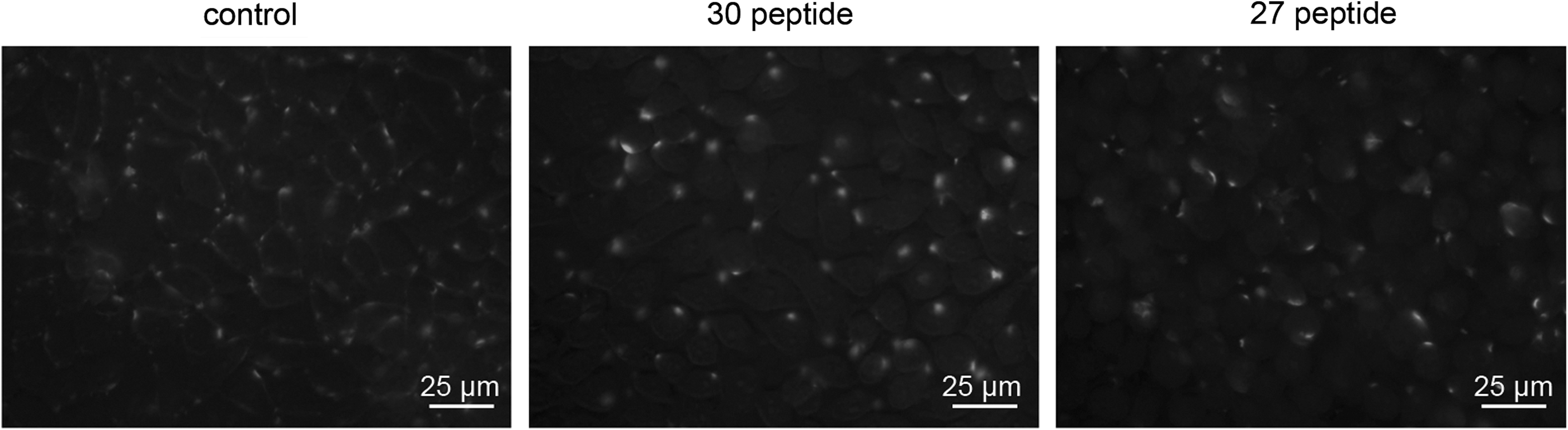
Peptide 30 increases integrin αvβ3 clustering on HepG2 cell surface. All photographs were taken under a fluorescent microscope at 400× magnification.
Peptide 30 does not change cell surface expression of integrin αvβ3
The expression of integrin avβ3 on the cell surface was not significantly changed by peptide 27 or peptide 30 treatment (Fig. 7).

Effect of peptide 30 on integrin αvβ3 cell surface expression levels, as measured by flow cytometry.
Peptide 30 suppresses MMP-2 and MMP-9 activity in HepG2 cells
MMP-2 and MMP-9 activity was analyzed 24 hours after peptide 30 treatment at 0, 20, 40, 60, 80, or 100 μg/mL. Peptide 30 inhibited MMP-2 and MMP-9 activity at concentrations over 40 μg/mL, with a gradual decrease in activity with increasing peptide concentration (Fig. 8A). Additionally, peptide 30 more powerfully suppressed MMP-2 and MMP-9 activity than peptide 27 when administered to HepG2 cells at 50 μg/mL for 24 hours (p<0.05) (Fig. 8B).
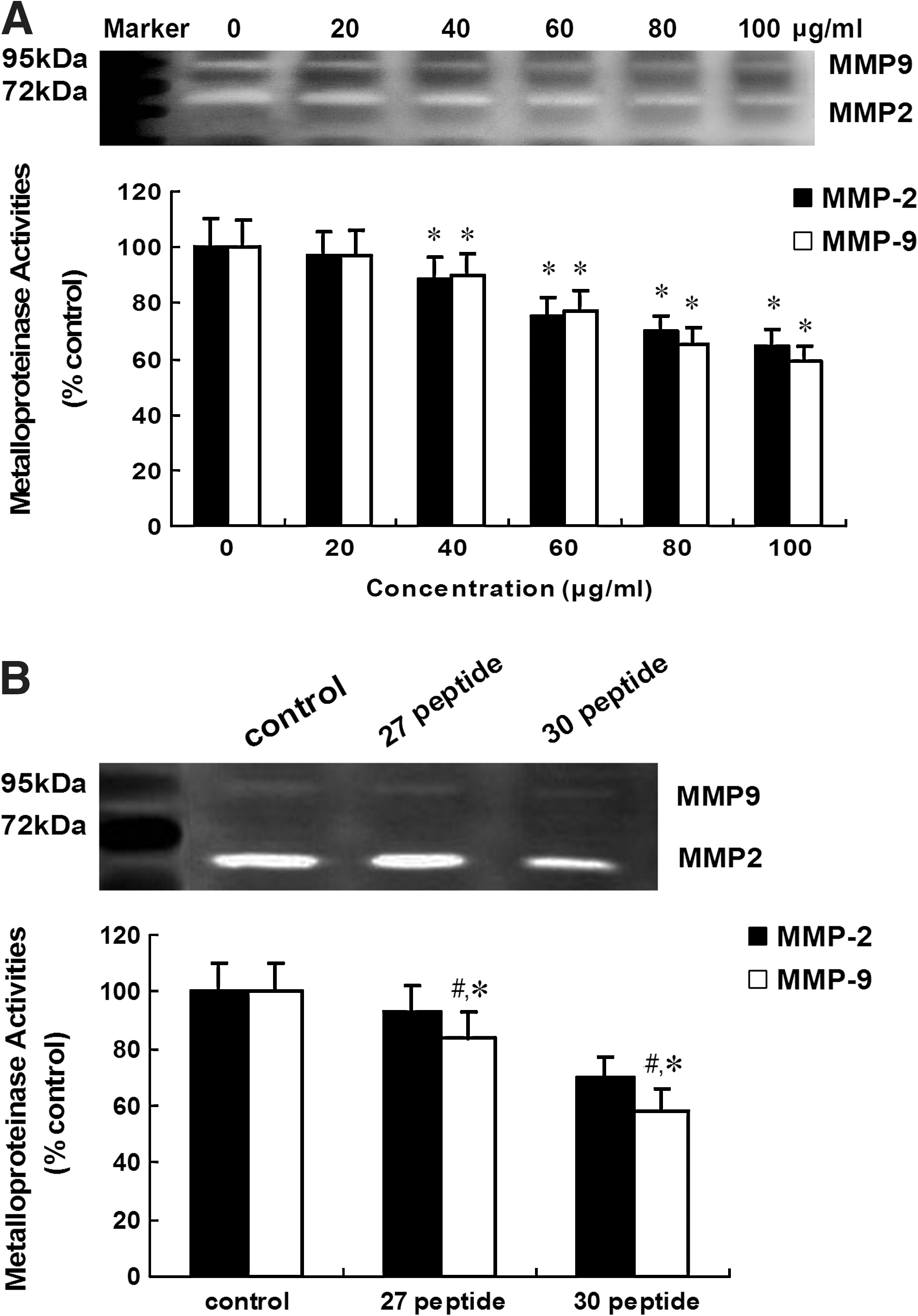
The effect of peptide 30 on cell gelatinase activities of HepG2 cells.
Peptide 30 affects COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 mRNA expression
As shown in Figure 9A–C, the mRNA expression of COX-2, MMP-2, and MMP-9 were downregulated by peptide 30 and peptide 27 treatment in a time-dependent manner. More specifically, the mRNA levels of COX-2, MMP-2, and MMP-9 were significantly decreased 24 hours after peptide 30 treatment, whereas the TIMP-1 and TIMP-2 mRNA levels were significantly upregulated (p<0.05). There was no significant change for integrins αv and β3 at the mRNA level (data not shown). Figure 9D demonstrates that combined peptide 30 and anti-αvβ3 treatment, compared with peptide 30 treatment alone, further downregulated COX-2, MMP-2, and MMP-9 and upregulated TIMP-1 and TIMP-2 at the mRNA level.
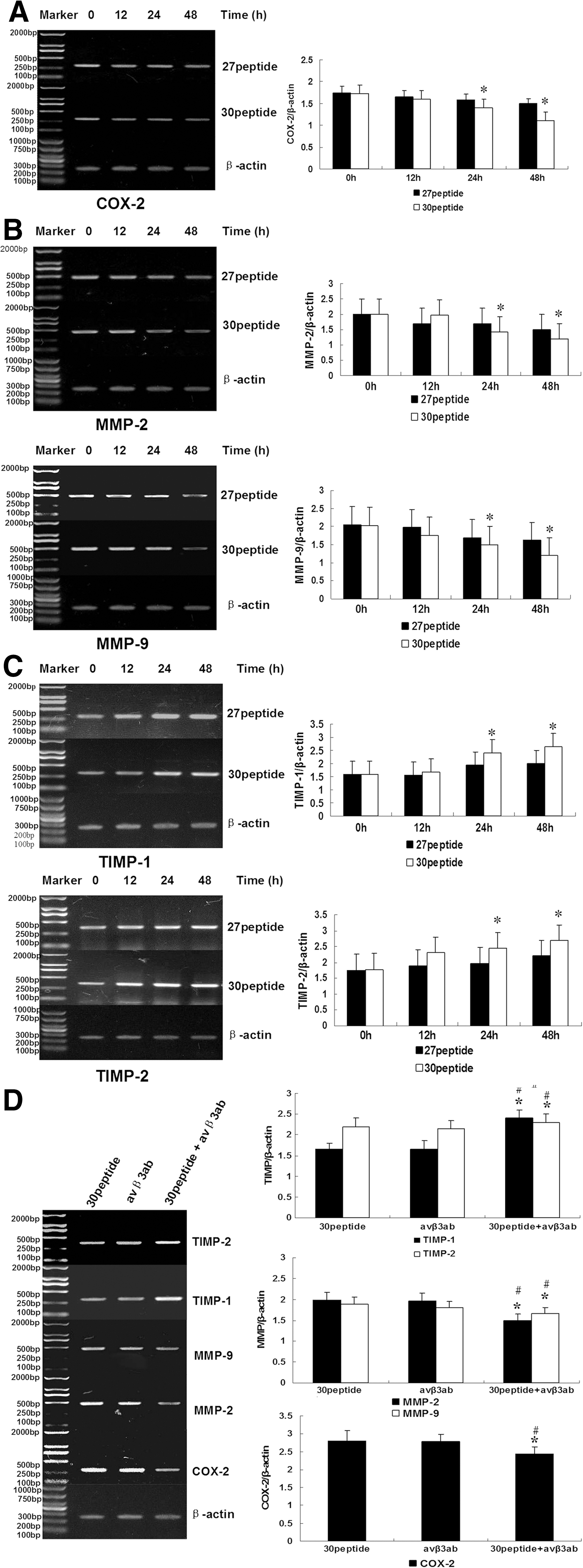
Peptide 30 affects COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 protein expression
Figure 10A–C shows that peptide 30 and peptide 27 downregulated COX-2 and its downstream targets MMP-2 and MMP-9 in a time-dependent manner. Compared with peptide 27, peptide 30 downregulated the expression of COX-2, MMP-2, and MMP-9 protein but upregulated TIMP-1 and TIMP-2 expression at 24 hours postpeptide treatment (p<0.05). In contrast, the protein expression of integrins αv and β3 was not significantly altered (data not shown). Compared with peptide 30 or αvβ3 antibody treatment alone, combined peptide 30 and αvβ3 antibody treatment significantly downregulated COX-2, MMP-2, and MMP-9 protein levels and upregulated TIMP-1 and TIMP-2 proteins (Fig. 10D).

Discussion
Invasion and metastasis are two key biological behaviors of malignancies that are at the root of failed surgery, chemotherapy, and radiation. They are also the most prevalent causes of cancer-related death. An ideal anticancer drug should not only suppress angiogenesis, inhibit cancer cell proliferation, and promote cancer cell apoptosis, but also potently suppress cancer cell invasion and metastasis. A previous study 3 indicated that the introduction of the RGDRGD sequence into endostatin-based peptide 30 preserved the protein's antiangiogenic activity in cancer tissue while suppressing cancer cell growth and inducing cancer cell apoptosis, enhancing the protein's antitumor potential. The present study focused on the specific effects of peptide 30 on cancer cell invasion and metastasis as well as its potential mechanisms of action, providing support for the peptide's clinical application.
Cancer metastasis is a complex process involving cell–cell and cell–extracellular matrix adhesion, extracellular matrix degradation, and cancer cell migration. 12,13 Adhesion to extracellular matrices and basement membranes is considered to be the initial step in the invasive process of metastatic tumor cells. 14 In vitro basal membrane invasion models imitate the in vivo invasion of cancer cells and reflect their invasion potential. 15,16
In the present study, based on an extracellular matrix adhesion assay following peptide treatment, peptide 30 more potently suppressed the adhesion of HepG2 cells to a matrigel matrix than peptide 27. This suppressive effect was dose dependent at peptide concentrations greater than 40 μg/mL. Additionally, in a matrigel-based invasion model, peptides at 50 μg/mL resulted in invading cell numbers of 132.9±3.1 and 71.4±2.8 for peptide 27- and peptide 30-treated cells, respectively, in contrast to 205.6±4.6 of the control cells. These results indicate that peptide 30 powerfully suppressed invasion by HepG2 cancer cells.
As early as 1863, Virchow 17 suggested that cancer cells are able to move, which was verified by Gey via a time-lapse video. From then on, the relationship between cell movement and metastatic potential has been the focus of research. To demonstrate the effects of peptide 30 on the random movement of HepG2 cells, a wound healing assay was performed in conjunction with peptide 30 or peptide 27 treatment at 50 μg/mL for 24 or 48 hours. The results demonstrate that these two peptides may suppress the random movement of cancer cells, with peptide 30 more actively inhibiting movement than peptide 27. Taken together, the data confirm the stronger inhibition of cancer cell adhesion, invasion, and migration by peptide 30 than by peptide 27.
Because the difference between peptide 30 and peptide 27 is only within the RGDRGD domain, it was supposed that the stronger ability of peptide 30 to suppress cancer cell adhesion, invasion, and migration was related to this domain. The RGD motif comprises the amino acids Arg-Gly-Asp and is frequently found in proteins of the extracellular matrix. RGD specifically improves the binding of extracellular matrix proteins to various types of integrins, relaying extracellular signals into the cells to induce either pathological or physiological reactions, including cancer cell adhesion, proliferation, metastasis, and apoptosis. 18
RGD-containing peptides can compete against extracellular matrix proteins for binding the integrins, resulting in the disintegrin effect. In the context of an invasion assay, antibodies against α1, α2, α3, α5, β1, αv, β3, or αvβ3 were added to the cell culture to screen the subtypes of integrin that bind to peptide 30. It was found that only those against αv and αvβ3 provided additional suppression of cancer cell invasion. RT-PCR and western blotting confirmed that anti-αvβ3 significantly enhanced peptide 30-induced regulation of COX-2, MMP-2, MMP-9, TIMP-1, and TIMP-2 expression. Flow cytometry analysis revealed that peptide 30 did not change the cell surface expression level of αvβ3, whereas immunofluorescent staining indicated clustering of αvβ3 on the cell surface. Therefore, it was hypothesized that peptide 30 impairs integrin function, rather than altering integrin expression levels, through RGD-mediated enhancement of selectivity and specificity of peptide 30 for the αvβ3 receptor, resulting in disintegrin effect and compromising the prometastatic effects of integrins.
It has been reported that inducible cyclooxygenase 2 (COX-2) is involved in the development, advancement, invasion, metastasis, differentiation, and prognosis of cancers. 19,20 Additionally, COX-2 has been positively associated with cancer malignancy. 21 Murphy et al. 22 have suggested that the integrin αvβ3 signaling pathway is involved in COX-2 regulation, whereas COX-2 may be closely incorporated with downstream matrix MMPs to regulate metastasis. 23 COX-2-regulated MMPs are secreted in a PG-dependent manner and exert effects on the adhesion and migration of hepatocarcinoma cells. 24,25 In the present study, RT-PCR and western blotting confirmed the downregulation of COX-2 and thus of downstream MMP-2 and MMP-9 by peptide 30. Moreover, TIMP-1 and TIMP-2, the natural inhibitors of MMP-2 and MMP-9, were upregulated. Anti-αvβ3 further decreased the levels of COX-2, MMP-2, and MMP-9 and increased the levels of TIMP-1 and TIMP-2. Finally, gelatin zymography revealed that, in a dose-dependent manner, peptide 30 suppressed the ability of cancer cells to degrade gelatin via MMPs. It was speculated that peptide 30 combined with αvβ3 through RGD and resulted in disintegrin effect. The blocking of αvβ3 at the cancer cell surface by αvβ3 antibodies synergistically compromised the integrin αvβ3 signaling, ultimately suppressing the invasion and metastasis of cancer cells.
In summary, RGD-modified endostatin peptide 30 may potently suppress the proliferation and metastasis of, as well as the angiogenesis and invasion by, the hepatocarcinoma cell line HepG2. Peptide 30 was more effective than peptide 27 in suppressing this invasion and metastasis. The mechanism of peptide 30 activity may involve the binding of the RGDRGD motif to integrin αvβ3, leading to downregulation of COX-2, MMP-2, and MMP-9 expression and upregulation of TIMP-1 and TIMP-2 and decreasing the ability of cancer cells to degrade the extracellular matrix. Therefore, peptide 30 may actively suppress the in vivo adhesion, invasion, and migration of cancer cells, encouraging its development as an adjuvant in the clinical treatment of cancer.
Footnotes
Acknowledgment
This work was supported by the Key Laboratory of Bio-pharmaceutical Engineering of Ministry of Education of China at Harbin Medical University,
Disclosure Statement
The authors declare that they have no conflict of interest.