
Abstract
The melanoma differentiation-associated gene-7/interleukin-24 gene (mda-7/IL-24) is a novel tumor-suppressor/cytokine gene that exhibits potent tumor-suppressive activity without damaging normal cells. To enhance the antitumor effect, an mda-7/IL-24 mutant, RGD-mda-7, which includes the cell adhesive sequence 164Arg-165Gly-166Asp (RGD motif), was constructed and evaluated for bioactivity. RGD peptide binds to integrins αVβ3 and αVβ5, which are selectively expressed in tumor neovasculature and in the surface of some tumor cells. The wtmda-7/IL-24 and RGD-mda-7 were expressed in Escherichia coli and then purified and renatured. The immunostimulatory activity of RGD-mda-7 was assayed by stimulating peripheral blood mononuclear cells. The results suggested that the abilities of RGD-mda-7 to induce IL-6, TNF-α, and IFN-γ production were higher than wtmda-7/IL-24. Tumor targeting of RGD-mda-7 was assayed using cell adhesion experiments. The antitumor effect of the purified RGD-mda-7 on cell proliferation in vitro was evaluated using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) uptake, cell apoptosis by staining with fluorescent probes of FITC-annexin V and DAPI, and caspase-3 expression and activity. The in vitro results showed that RGD-mda-7 inhibited the proliferation of multiple tumor cell lines (Hela, ACHN, HepG2, and A549). Staining with fluorescent probes of FITC-annexin V and DAPI indicated that RGD-mda-7 could induce apoptosis more effectively in four tumor cell lines than wtmda-7/IL-24, but has no effect on normal cell line NHLF. Western blotting showed that treatment of tumor cells with RGD-mda-7 could activate apoptotic pathway by cleavage of caspase-3 as same as wtmda-7/IL-24. Further, RGD-mda-7 group showed a higher cleaved level of caspase-3, but not in NHLF cells. These results demonstrate that RGD-MDA-7 possesses more potent antitumor effects than wtmda-7/IL-24 and therefore merits further investigation in preclinical and clinical studies.
Introduction
Melanoma differentiation-associated gene-7 (mda-7) was identified and cloned using the differentiation induction subtraction hybridization approach after treating the HO-1 human melanoma cell line with IFN-β and mezerein, which resulted in growth arrest and terminal differentiation. 1 Based on structure (containing an interleukin [IL]-10 signature motif), secretion by cells (including subsets of T-cells), and location on chromosome 1q3 (in an area containing IL-10 family genes), mda-7 was redesignated as IL-24, belonging to the IL-10 cytokine family. 2 –4 The results of in vitro studies, in vivo animal studies, and a phase I clinical trial indicate that MDA-7/IL-24 has the ability to selectively induce apoptosis in numerous human cancer cell lines. 5 –11 In contrast, no significant growth-inhibitory effect occurred when this gene was transduced into normal human breast or prostate epithelial, endothelial, melanocyte, astrocyte, or fibroblast cells. 4,10,12 –14 Intriguingly, in addition to its direct apoptosis inducing properties, mda-7/IL-24 also shows antiangiogenic, radiosensitizing, immunostimulatory, and potent “bystander” antitumor activities. 3,15 –19
Although Ad.mda-7(INGN-241) has entered phase II clinical trials, 20 the major hindrance to its widespread use is to ensure effective delivery of the molecule to all cancer cells. 21 Thus, to further augment cancer-specific targeting and enhance the antitumor effect, an mda-7/IL-24 mutant, RGD-mda-7, was constructed and investigated.
The RGD (arginine–glycine–aspartic acid) peptide is an integrin-recognition motif found in many ligands. Integrins αVβ3 and αVβ5 serve as receptors for a variety of extracellular matrix proteins with the exposed RGD peptide sequence. 22 Integrins αVβ3 and αVβ5 is overexpressed on both multiple tumor cells (osteosarcomas, neuroblastomas, glioblastomas, melanomas, lung carcinomas, breast cancer, and so on) and the activated endothelial cells of tumors' neovasculature. 23 –32 The highly restricted expression of integrins αVβ3 and αVβ5 correlates well with tumor growth, progression, invasion, and metastasis. 22 When coupled to the anticancer drug, the RGD peptide could develop targeting and enhance the antitumor effect of the drug. 33 Moreover, RGD-containing peptides are able to directly induce apoptosis by triggering conformational changes that promote pro-caspase-3 autoprocessing and activation. 34
In the present study, the mda-7/IL-24 mutant RGD-mda-7 was constructed by inserting a glycine residue into the wtmda-7/IL-24 between Arg164 and Asp165 to form an RGD motif, and then the relevance of RGD-mda-7 in mediating cancer-selective cell killing and the ability to target tumor cells were assessed. Results indicate that RGD-mda-7 enhances wtmda-7/IL-24 antitumor effect in four tumor cell lines (Hela, ACHN, HepG2, and A549). Further, the present study documents that RGD-mda-7 induced apoptosis by activation of caspase-3.
Materials and Methods
Construction of expression vectors for mda-7/IL-24 and mutant RGD-MDA-7
The whole cDNA coding region of mda-7/IL-24 gene was kindly provided by Professor Liu (Xin-Yuan Liu, Institute of Biochemistry and Cell Biology, Chinese Academy of Sciences, Shanghai, China) and used as template for overlapping polymerase chain reaction (PCR). Four primers for PCR were designed as follows based on the mda-7/IL-24 sequence and the pET-28a(+)vector: P1: 5′-TGTG P2: 5′-ACTGTC
P3: 5′-ATCAGA
P4: 5′-TGTG
The P1 had Enterokinase restriction sites shown in italics. The P2 and P3 had the overlapping complementary 15-base sequence shown in italics, which resulted in the extra codon
Expression, purification, and renaturation of Wtmda-7/IL-24 and RGD-mda-7 proteins
The successfully constructed plasmid was used to transform Escherichia coli strain BL21(DE3) (resistant to 15 μg/mL kanamycin) competent cells. The positive clone cells were inoculated to overnight express instant TB medium (Novagen) containing 15 μg/mL kanamycin. The recombinant proteins were purified by His-tag affinity purification under denaturing conditions using BugBuster Ni-NTA His Bind Purification Kit (Novagen) and then renatured using Protein Refolding Kit (Novagen). All kits were used according to the manufacturer's instruction. Protein purity and identity were checked by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting.
IL-6, TNF-α, and IFN-γ production assay
Human peripheral blood mononuclear cells (PBMCs) were obtained from healthy volunteer donors by centrifugation over Ficoll–Hypaque density gradient separation. Monocytes were cultured in RPMI1640 medium containing 100 U/mL penicillin, 100 mg/L streptomycin, and 10% fetal bovine serum at 1×106 cells/mL, with the wtmda-7/IL-24 and RGD-mda-7 recombinated proteins added at the designated concentrations. The cells were incubated at 37°C in a 95% humidified 5% CO2 environment. At indicated times, the culture supernatants were harvested and the amount of IL-6, TNF-α, and IFN-γ was measured by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instruction (ZhongShan GoldenBridge Company).
Cell line and culture
The human lung cancer cell line A549 and normal human lung fibroblast cell line NHLF were purchased from Chinese Academy of Sciences Cell Bank. The human liver cancer cell line HepG-2 and uterine cervix cancer cell line HeLa were purchased from Tongji University. The human renal carcinoma cell line ACHN was available in the authors' laboratory. 35 All cell lines were maintained in RPMI 1640 medium supplemented with 10% fetal bovine serum (Gibco-BRL) and 1% K-glutamine/antimycotic solution. The cells were grown at 37°C in a humidified atmosphere containing 5% CO2.
Cell adhesion assay and adhesion inhibiting assay
Polyvinyl chloride microtiter plates (96 well) were coated, respectively, with RGD-mda-7 (8 μg/mL) and wtmda-7/IL-24 solutions (8 μg/mL) at 4°C overnight. After being washed with 0.9% sodium chloride, each well was filled with RPMI 1640 medium containing 1% bovine serum albumin for 2 hours at 37°C and washed again. A549 cells, HepG-2 cells, HeLa cells, and ACHN cells were detached using trypsin digestion, washed three times with 0.9% sodium chloride, resuspended in incomplete RPMI 1640 medium, and then added to RGD-mda-7– and wtmda-7/IL-24–coated plates with 2×104 cells/well in 100 μL. After incubation for 1 hour at 37°C in 5% CO2, unbound cells were removed by washing with incomplete RPMI 1640 medium. Adherent cells were fixed with a solution of 4% paraformaldehyde in phosphate-buffered saline (PBS; pH 7.2) and stained with 0.5% crystal violet. Absorbance from the plates was read on an ELX-800 spectrometer reader (Bio-Tek Instruments, Inc.) at 490 nm. The experiment was repeated more than four times. For adhesion inhibiting assay, the cells were stained with monoclonal antibodies directed against specific integrins. The antibodies that were used include LM609 (anti-avb3) and 15F11 (anti-avb5). Antibodies were incubated with cells in suspension for 30 minutes at an antibody concentration of 15 μg/mL. Then the assay was carried out in the same way as the adhesion assay.
Cell viability assay
Cells were plated in 96-well plates and treated with the RGD-mda-7 and wtmda-7/IL-24 proteins at the designated concentrations. At the indicated times, the medium was removed and fresh medium containing 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, 0.5 mg/mL, 100 μL) was added to each well. Cells were incubated at 37°C for 4 hours and then the medium was removed, and 150 μL solubilization solution (DMSO) was added and mixed thoroughly. Absorbance from the plates was read on an ELX-800 spectrometer reader (Bio-Tek Instruments, Inc.) at 490 nm.
Annexin V binding assays
Cells were analyzed for apoptosis, using the FITC-labeled annexin V (KeyGen Biotech Co., Ltd.). Briefly, cells were plated in 24-well plates, treated with RGD-mda-7 and wtmda-7/IL-24 proteins, and incubated at 37°C in humidified 5% CO2 for 48 hours. Cells were washed once with complete medium and PBS, added to 0.5 mL of binding buffer, stained with annexin V for 10 minutes at room temperature, and observed using a fluorescence microscope (Nikon; ECLIPSE-Ti).
DAPI staining
Cells were plated in 24-well plates, treated with the RGD-mda-7 and wtmda-7/IL-24 proteins, and incubated at 37°C in humidified 5% CO2 for 72 hours. Cells were washed twice with cold PBS (pH 7.2) and stained with 1 μg/mL DAPI (KeyGen Biotech Co., Ltd.) at 37°C for 30 minutes. Nuclear morphology was observed using a fluorescence microscope (Nikon; ECLIPSE-Ti).
Western blotting analysis
Cell lines were grown on 10-cm plates and protein extracts were prepared with RIPA buffer containing a cocktail of protease inhibitors. A measure of 50 mg of protein was applied to 15% SDS-PAGE and transferred to nitrocellulose membranes. The membranes were probed with polyclonal or monoclonal antibodies to mda-7/IL-24 and caspase-3.
Statistical analysis
Values were expressed as mean±SD, and statistical analysis of the results was carried out by one-way analysis of the variance followed by Duncan's new multiple range method or Newman–Keuls test. p-Values<0.05 were considered significant.
Results
Construction of mutant RGD-mda-7
By means of overlapping PCR, a Gly was inserted between Arg164 and Asp165 into wtmda-7/IL-24, and mutant RGD-mda-7 was created. The RGD-mda-7 was cloned into pET-28a(+) and transformed into DH5α competent cells. The expected mutation was confirmed by DNA sequencing (Fig. 1A, B).
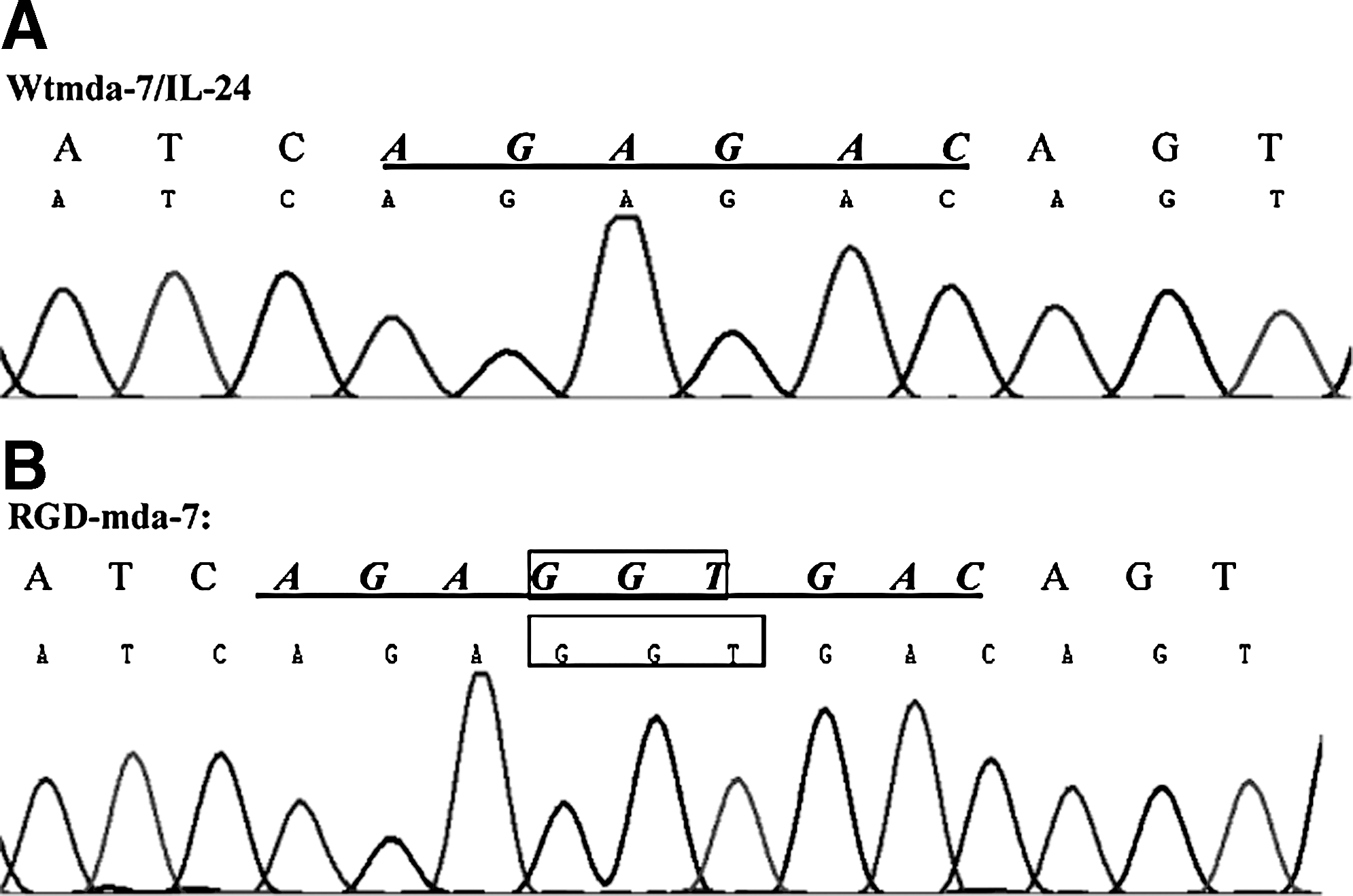
DNA sequence alignment of wtmda-7/IL-24
Expression, purification, and renaturation of the recombinant wild-type and mutant proteins
The transformed BL21(DE3) cells were inoculated to overnight express instant TB medium at 37°C for 16 hours. After induction, the cells were dissolved in SDS-PAGE sample buffer and subjected to SDS-PAGE. The result showed that the recombinant wtmda-7/IL-24 and RGD-mda-7 proteins were efficiently expressed, with the yield accounting for 30% of total bacterial proteins. SDS-PAGE analysis also showed that the recombinant proteins were mainly in the precipitation of cell lysate, indicating that the recombinant proteins predominantly existed in the inclusion bodies. After washing with 2 M urea, the recombinant proteins were purified under denaturing conditions with Nickel metal-affinity resin columns for single-step purifications of His-tagged in wtmda-7/IL-24 and RGD-mda-7. The purified proteins were renatured by step-by-step dilution using protein refolding kit. The purity of purified proteins was about 95% (Fig. 2A, B). Western blot analysis confirmed that RGD-mda-7 was recognized by anti-mda-7/IL-24 monoclonal antibody (Fig. 2C).
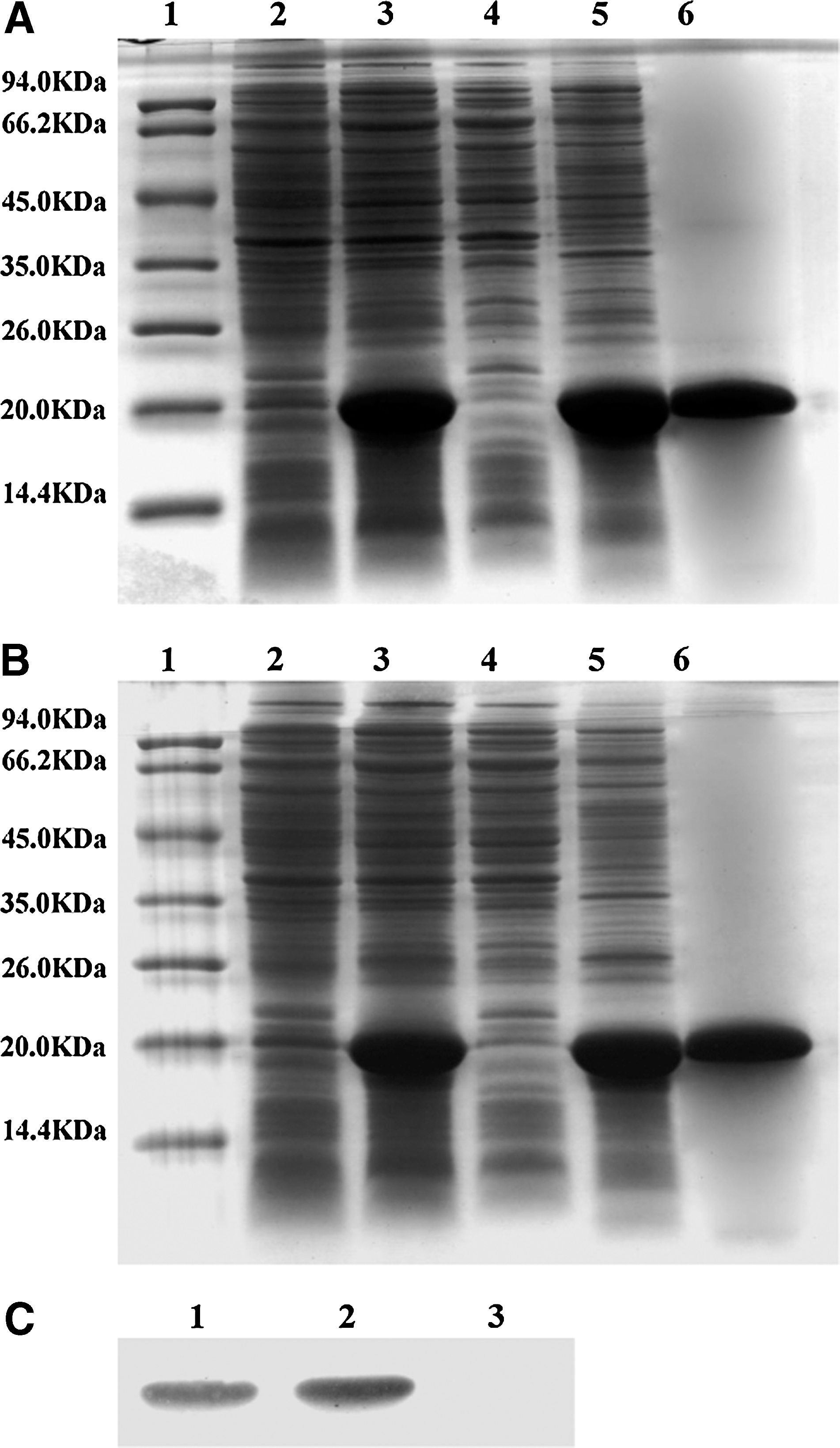
Expression, purification, and identification of RGD-mda-7 and wtmda-7/IL-24.
IL-6, TNF-α, and IFN-γ production assay
To study the bioactivity of RGD-mda-7, the induction of IL-6, TNF-α, and IFN-γ from PBMCs by mutant protein was assayed. Optimum levels of wtmda-7/IL-24 and RGD-mda-7 to induce maximal IL-6, TNF-α, and IFN-γ secretion were determined in a dose–response curve. Moreover, as shown in Figure 3, the abilities of inducing IL-6, TNF-α, and IFN-γ production by RGD-mda-7 were higher than wtmda-7/IL-24, suggesting that the immunostimulatory activity of purified RGD-mda-7 was more effective than that of wtmda-7/IL-24.
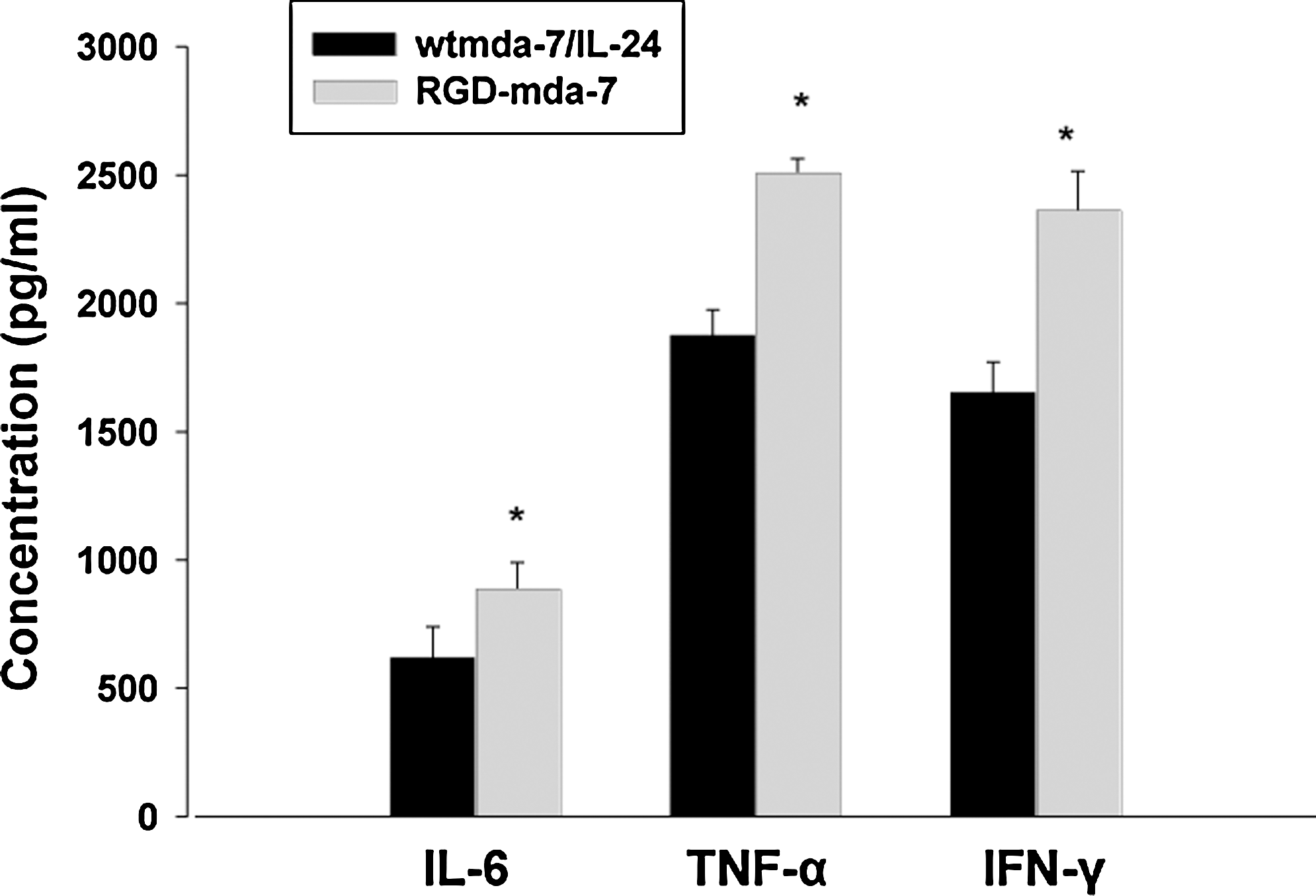
Effect of RGD-mda-7 on IL-6, TNF-α, and IFN-γ inflammatory cytokine secretion from PBMCs. Two (2) milliliters per well (1×106 cells/mL) PBMCs were plated in a 24-well plate and cultured with 0.2 μg/mL wtmda-7/IL-24 and RGD-mda-7. Supernatants were harvested at 48 hours and analyzed for cytokine content by enzyme-linked immunosorbent assay according to the manufacturer's instructions. Data were expressed as mean±SD from six independent groups (n=6). *p<0.05 versus wild-type mda-7/IL-24. PBMCs, peripheral blood mononuclear cells.
RGD motif targets mda-7/IL-24 to cancer cells
To assess whether the RGD domain of RGD-mda-7 is functional and accessible to integrins αVβ3 and αVβ5, cell adhesion promoted by RGD-mda-7 was compared with that promoted by wtmda-7/IL-24. Crystal violet staining showed adhesion and spreading of A549 cells, HepG-2 cells, HeLa cells, and ACHN cells on plates coated with RGD-mda-7, but not on plates coated with wtmda-7/IL-24 (Fig. 4). These results indicate that RGD domain of RGD-mda-7 was properly folded and able to interact with adhesion receptors integrins αVβ3 and αVβ5 in the cell membrane.
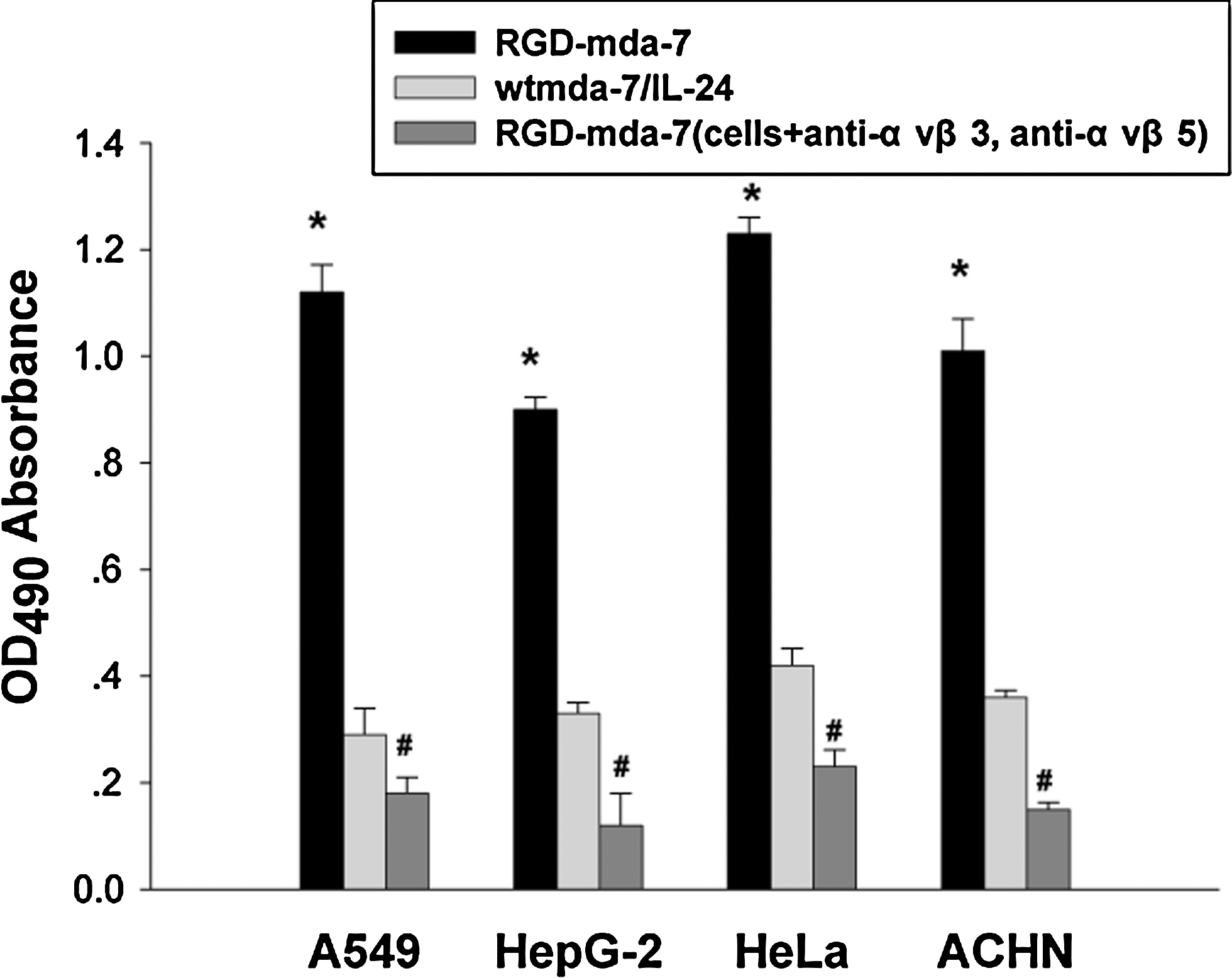
The binding specificity of RGD-mda-7 to cancer cells. Microtiter wells were coated with RGD-mda-7 (8 μg/mL) or wtmda-7/IL-24 and seeded with the tumor cells, which were incubated with antibodies anti-αVβ3 and anti-αVβ5. Absorbance was read at 490 nm. Data were expressed as mean±SD from six independent groups (n=6). *p<0.05 versus wild-type mda-7/IL-24; # p<0.05 versus cells+anti-αvβ3/anti-αvβ5.
Potent cytopathic effect induced by RGD-mda-7
To assess the cytopathic effect of RGD-mda-7, MTT assay was also performed to examine cell viability. Cells (HepG2, A549, HeLa, and ACHN) were plated in 96-well plates and treated with RGD-mda-7 compared with wtmda-7/IL-24 at different concentrations as indicated. As shown in Figure 5, treatment of HepG2, A549, HeLa, and ACHN cells with RGD-mda-7 resulted in a time- and dose-dependent CPE, and as was expected, RGD-mda-7 could kill tumor cells more powerfully than the wild mda-7/IL-24.

The effect of RGD-mda-7 on the growth of tumor cells.
RGD-mda-7 induces apoptosis in tumor cells but not in normal fibroblasts
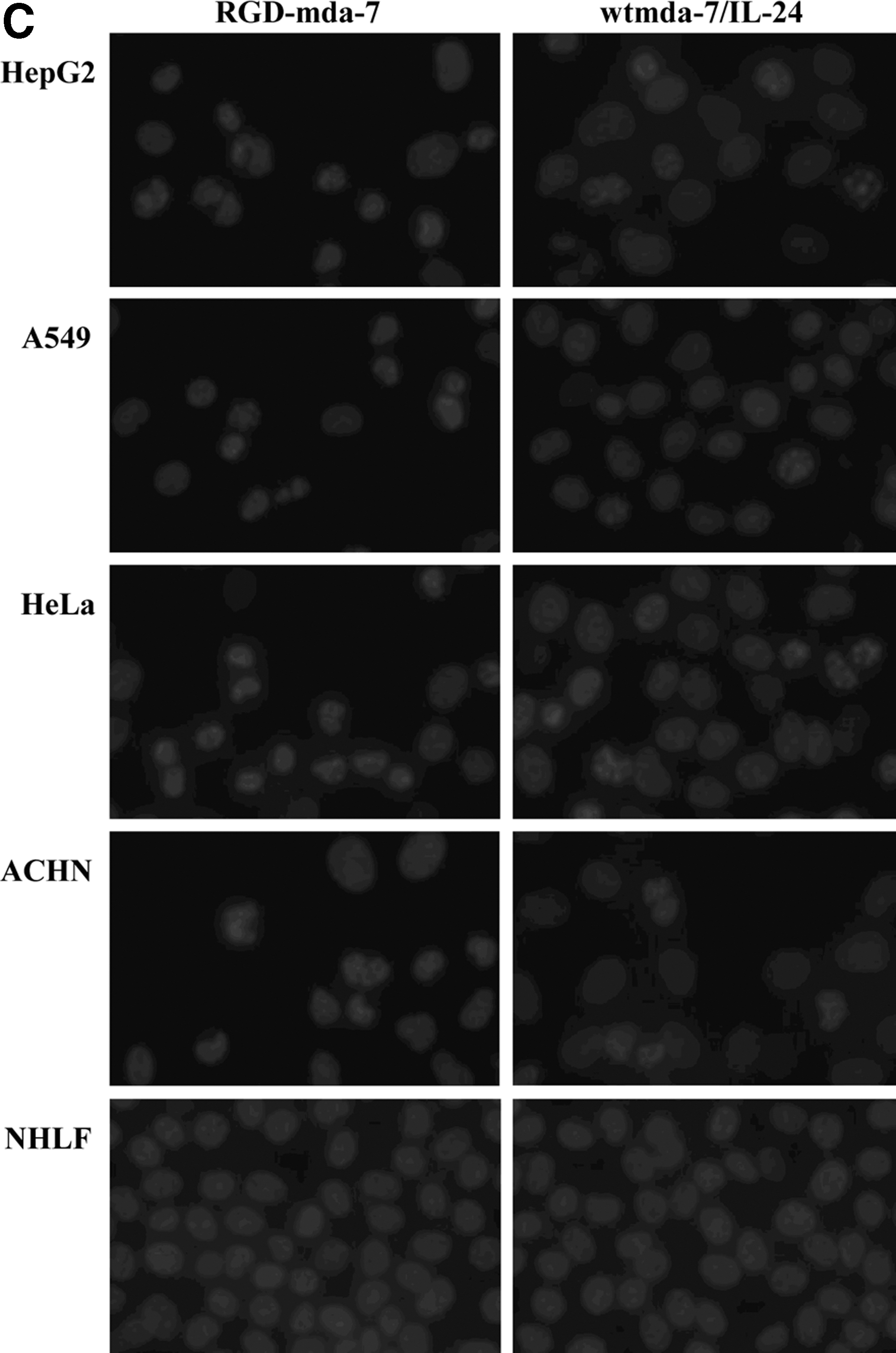
Following treatment of RGD-mda-7 or wtmda-7/IL-24, tumor cells (HepG2, A549, HeLa, and ACHN) and normal cells (NHLF) were analyzed for apoptotic change by annexin V assay and DAPI staining. After 48 hours, treatment of tumor cells with RGD-mda-7 resulted in a significant increase in the number of annexin V-positive cells compared with results for cells treated with wtmda-7/IL-24. In contrast, normal cells treated with either RGD-mda-7 or wtmda-7/IL-24 did not demonstrate any change in the number of annexin V-positive cells (Fig. 6A, B). To confirm these results further, DAPI staining was performed 72 hours after treatment. Tumor cells but not normal cells were observed to undergo apoptosis following treatment (Fig. 6C, D).
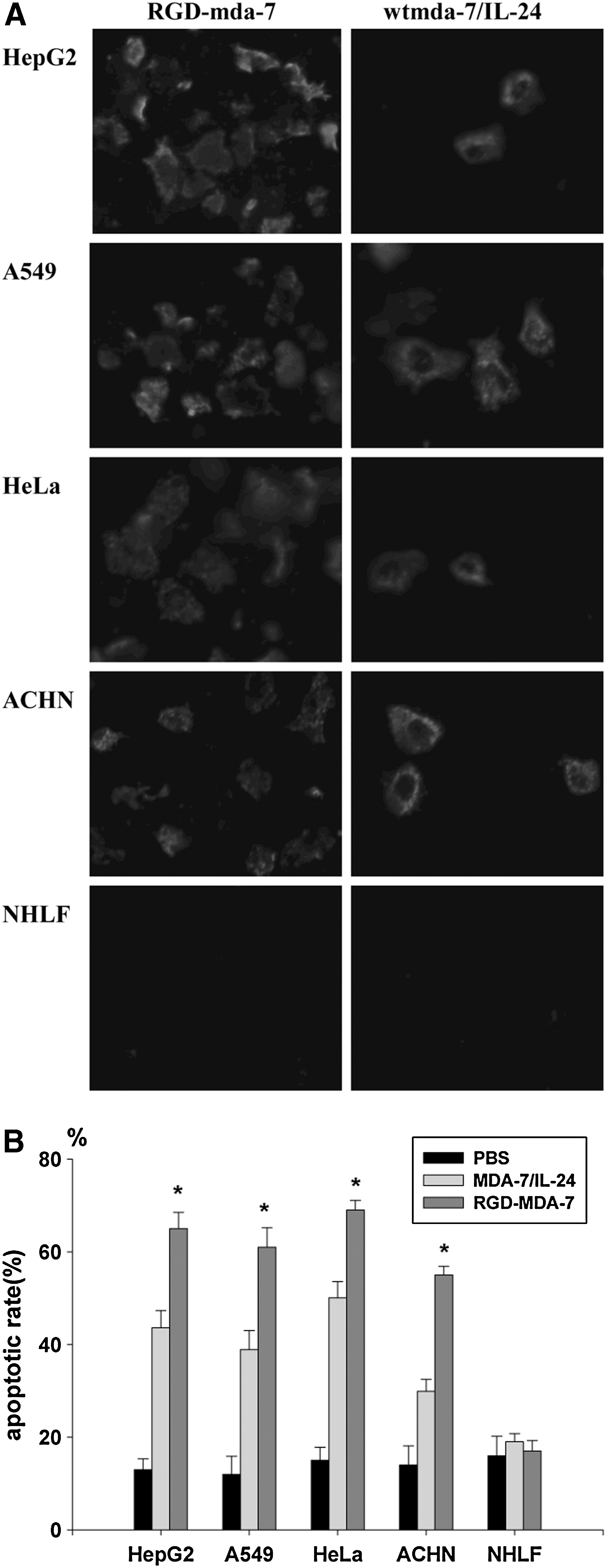
RGD-mda-7 induced apoptosis in tumor cells but not normal cells. Tumor cells (HepG2, A549, HeLa, and ACHN) and NHLF cells were treated with RGD-mda-7, wtmda-7/IL-24, and PBS, respectively, at a concentration of 8 μg/mL.

Activation and cleavage of caspase-3 following treatment with RGD-mda-7 and wtmda-7/IL-24
As mda-7 induces apoptosis in tumor cells, the activation of caspase-3 was studied by western blot analysis. These analyses demonstrated a higher cleaved level of caspase-3 in tumor cells treated with RGD-mda-7, whereas caspase-3 was not cleaved in NHLF cells (Fig. 7).
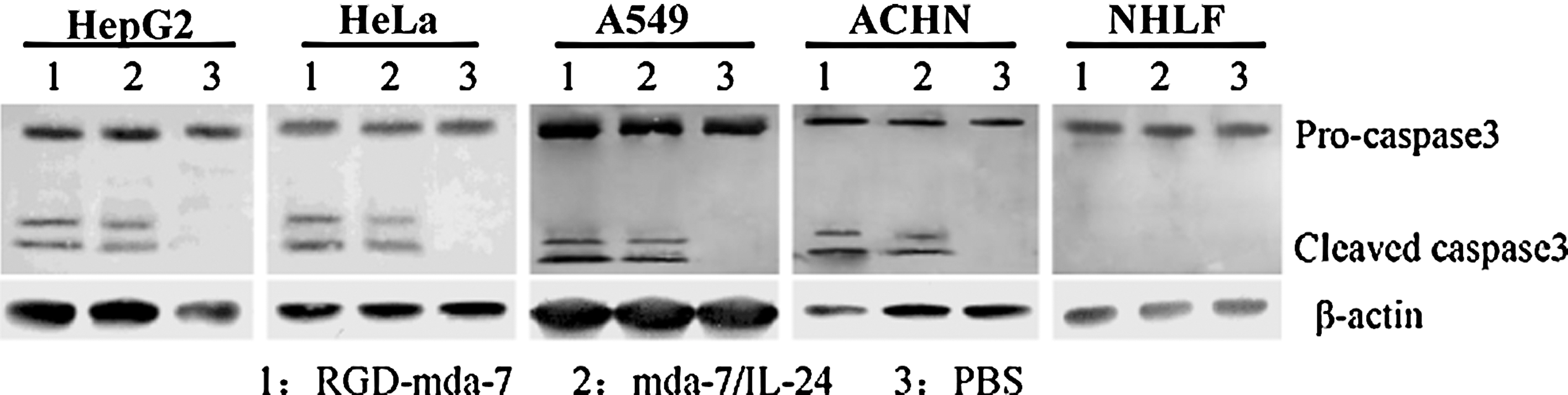
Mediation of apoptosis by RGD-mda-7 via caspase-3 activation. HepG2, A549, HeLa, ACHN, and NHLF cells were treated with RGD-mda-7 or wtmda-7/IL-24. After 48 hours, proteins were extracted from cells and separated by SDS-PAGE. Activation of caspase-3 was observed after treatment with RGD-mda-7 or wtmda-7/IL-24 in HepG2, A549, HeLa, and ACHN cells. No cleavage of caspase-3 was detectable in any PBS group. In NHLF cells, activation of caspase-3 was not seen.
Discussion
A recent phase I clinical trial indicates that mda-7/IL-24 is safe and induces significant clinical activity in patients with solid tumors. 36 These observations confirm the cancer therapeutic properties of this intriguing cytokine. The combination of direct cancer-specific apoptosis induction and indirect antitumor properties makes mda-7/IL-24 an ideal cancer therapeutic. The most important fail-safe for mda-7/IL-24 is its nontoxicity toward normal cells that might be exploited to target expression of mda-7/IL-24 to both cancer and normal cells, the latter serving as a reservoir for continuous secretion of mda-7/IL-24 protein, thereby exerting profound bystander antitumor effects. It is imperative that mda-7/IL-24 be delivered by a vector system that employs a universal entry system in the cells and is also shielded from immunologic clearance from the circulation.
In the present study, the mda-7/IL-24 mutant RGD-mda-7 was constructed and the relevance of RGD-mda-7 in mediating cancer-selective cell killing and the ability to target tumor cells were assessed. PCR mutagenesis has been used to insert a Gly between Arg164 and Asp165 of the wtmda-7/IL-24 polypeptide chain to form mutant RGD-mda-7. The recombinant proteins were efficiently expressed in E. coli strain BL21(DE3) and then purified and renatured.
Data indicate that mda-7/IL-24 protein functions as a pro-Th1 cytokine in PBMCs and induces secretion of IL-6, TNF-α, and IFN-γ. 2,3 To address the biological function of RGD-mda-7, its induction of IL-6, TNF-α, and IFN-γ secretion by PBMCs was examined. ELISA showed that RGD-mda-7 has the ability to induce IL-6, TNF-α, and IFN-γ production from PBMCs, which is more effective than that of wtmda-7/IL-24. It is well documented that the inserted Gly enhanced the immunostimulatory activity of wtmda-7/IL-24 protein.
Integrins αVβ3 and αVβ5 play a significant role in tumor progression, invasion, and metastasis and is a receptor for RGD tripeptide sequence. 22,32 To investigate whether the RGD domain of RGD-mda-7 is functional and accessible to integrins αVβ3 and αVβ5, cell adhesion enhanced by RGD-mda-7 was observed and compared with wtmda-7/IL-24. Figure 4 shows adhesion and spreading of A549 cells, HepG-2 cells, HeLa cells, and ACHN cells on plates coated with RGD-mda-7 but not on plates coated with wtmda-7/IL-24. These results indicate that RGD domain of RGD-mda-7 was properly folded and able to interact with integrins αVβ3 and αVβ5 and targets mda-7/IL-24 to cancer cells.
In the present study, significant cytopathic capacity was observed by MTT assay in all tumor cell lines treated with RGD-mda-7, compared with wtmda-7/IL-24. This cytopathic effect was time and dosage dependent. Apparently, RGD-mda-7 achieved antitumoral efficacy much more superior to wtmda-7/IL-24 by RGD sequence to interact with integrins αVβ3 and αVβ5 in the tumor cell membrane. These results suggest that the Gly inserted into the wtmda-7/IL-24 does not disrupt its biological function and selective killing effects in tumor cells, and that RGD-mda-7 inhibits the growth of tumor cells more effectively than wtmda-7/IL-24.
As shown in Figure 6, indicating RGD-mda-7 as a potent inducer of apoptosis in tumor cell lines, the nuclear morphology was further analyzed by DAPI staining. RGD-mda-7 elicited typical apoptotic morphological changes in tumor cells, including chromatin condensation and apoptotic body. In contrast, no significant change was found in NHLF cell control group. Moreover, statistical analysis showed that RGD-mda-7 had much stronger apoptosis-induced activity in tumor cells than wtmda-7/IL-24. A possible explanation for the difference is that RGD-mda-7 enhanced the antitumor potency of wtmda-7/IL-24.
A number of studies have sought to identify molecules involved in apoptotic signaling by mda-7/IL-24 treatment of tumor cells. Apoptotic mediators that are upregulated or activated by mda-7/IL-24 include caspases, p53, BAX, BAK, TRAIL, Fas, and DR4. 37 Signaling molecules implicated in mda-7/IL-24-mediated apoptosis include PKR, p38 MAPK, PI3K, JNK, and GSK-3. 37,38 It is evident, based on existing data, that the wide spectrum of tumor-specific apoptotic activity of mda-7/IL-24 involves multiple pathways. These molecules appear to be regulated only in tumor cells. This research showed that activation of caspase-3 was observed in tumor cells treated with RGD-mda-7 and wtmda-7/IL-24 but not in untreated control cells (Fig. 7). These results demonstrate that RGD-mda-7 induced apoptosis and activated caspase-3.
Conclusions
Gly was inserted into mda-7/IL-24 amino acid sequence to construct an RGD domain, which does not only disrupt mda-7/IL-24's biological function, but also enhance growth suppression and apoptosis in multiple human tumor cell lines, but not in normal cell line (NHLF). Although the antitumor mechanism RGD-mda-7 has to be further elucidated for future use, the preliminary data of this study indicate that RGD-mda-7 is an effective approach to enhance the antitumoral potency of mda-7/IL-24.
Footnotes
Acknowledgments
This project was supported by grants from the National Natural Science Foundation of China (No. 30873021), the Science and Technology Department of Jiangsu province (No. BK2007032), Health Departmental of Jiangsu province (No. H200724), and Program for New Century Excellent Talents in University (NCET-08-0700).
Disclosure Statement
No competing financial interests exist.