
Abstract
Background:
Antibodies have been proved to be effective in cancer treatment. Peroxiredoxin I (Prx I) is a potential target for cancer radiotherapy. The aim of this article is to investigate the effect of a novel phage display single-chain variable fragment (scFv) antibody targeting Prx I on human lung carcinoma cell line A549 radiosensitivity and the underlying mechanisms.
Materials and
Results:
Prx I scFv incubation significantly increased A549 cell radiosensitivity, and this might be through enhanced intracellular ROS level and caspase 3 expression. In addition, protein expression of radiosensitivity-related proteins, RAD51 and γ-H2AX, was also modulated. The nude mouse xenograft model bearing A549 tumor treated with Prx I scFv also exibited enhanced radiosensitivity compared with the PBS group.
Conclusions:
These results suggested that Prx I scFv could become a new therapy candidate for lung cancer radiotherapy.
Introduction
Lung carcinoma is now one of the most common and lethal malignancy. A total of 219,440 new lung cancer cases and 159,390 deaths from lung cancer are evaluated to occur in the United States in 2009, accounting for 30% of total deaths from cancer. 1
Due to the high occurrence and mortality rates of lung cancer, the knowledge about lung cancer diagnosis and treatment has always been a concern. Since patients with lung cancer usually present no characteristic symptoms in early stage and are usually diagnosed at advanced stages, treatment of cancer remains a formidable challenge. Up to now, radiotherapy and chemotherapy are the main methods. Lung cancer is resistant to radiotherapy, resulting in ineffectiveness of radiation treatment, so it is of vital clinical significance to explore new therapies to enhance radiosensitivity of lung carcinoma. Ionizing radiation (IR) could evoke a series of biochemical events inside the cell, including DNA damage and repair, apoptosis, cell cycle control, signal transduction, and oxidative stress response. Among these effects, reactive oxygen species (ROS) induced by IR is crucial for cell survival. 2,3 Carcinoma cells have its antioxidant enzymatic systems that could eliminate the production of activated oxygen species, among which is the recently characterized peroxiredoxins (Prxs).
Prx I is a member of the Prxs family and has been shown to be upregulated in a wide variety of cancer cell lines, including lung adenocarcinoma. 4 –6 Upregulation of Prx I could enhance the oxidative resistance of carcinoma cells and was thought to be one of the most important reasons for chemotherapy resistance. 5 Recently, downregulation of Prx I expression was found to have some relations with enhanced radiosensitivity of cancer cells; decreased Prx I expression by siRNA strategy enhanced the IR-induced cell death in vitro and enhanced the IR effects on tumor xenografts. 7
Several strategies such as RNA interference, antibody, and antisense have been developed to knockdown gene expression or decrease protein expression, which is crucial for cancer cell survival or metastasis. Among these strategies, antibodies have been proved to be effective in clinical cancer treatment through the specific conjunction of antibody and antigen. 8 Phage display technology is an attractive method that avoids the deficiencies of hybridoma technology, generating antibodies from both immune and nonimmune sources. 9 A novel phage display single-chain variable fragment (scFv) antibody targeting Prx I of lung adenocarcinoma has been successfully produced in our laboratory and has been demonstrated to have the potential as a new candidate for lung cancer therapy. 10
In the present study, we investigated the effect of Prx I scFv on A549 cell radiosensitivity and explored the underlying mechanisms, aiming to provide foundations for searching new method for increasing lung cancer radiosensitivity.
Materials and Methods
Cell culture
The human lung carcinoma cell line A549 was cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with glutamine (4 mM), penicillin (100 i.u./mL), streptomycin (100 μg/mL), and 10% fetal bovine serum (Hangzhou Sijiqing Co. Ltd.) in a humidified incubator with 5% CO2. Cells were passaged every 4 days; cells from passages 3–20 were used in these experiments.
Flow cytometric analysis for cell cycle, cell death, and ROS production
Flow cytometry was performed on a FACScan flow cytometer equipped with an argon laser at 488 nm excitation (Becton Dickinson). To detect A549 cells apoptosis rate, cells were processed with different treatments, then washed twice with PBS, and resuspended in 200 μL of binding buffer (10 mM HEPES, pH 7.4, 140 mM NaCl, 1 mM MgCl2, 5 mM KCl, and 2.5 mM CaCl2). Ten (10) microliters of FITC-conjugated AnnexinV was then added to give a final concentration of 0.5 μg/mL. The staining sample was incubated at room temperature for 20 minutes, without light. Subsequently, 5 μL of propidium iodine (PI) was added to the samples (final concentration of 1 μg/mL) and 10,000 cells were immediately analyzed. Cell cycle profiles were analyzed using MultiCycle software (PHENIX), and the sub-G1 peak was quantified to estimate apoptosis in the population. To test cell cycle, PI fluorescence of individual nuclei was measured by FCM. The cell cycle phase and the proportion of cells in G1 phase was determined using Multicycle software. To examine the production of ROS, cells were harvested in phosphate-buffered saline and incubated in the presence of 20 mM 2979-dichlorodihydrofluorescein (H2DCF) diacetate (Molecular Probes) in the dark at 37°C for 30 minutes. The shift in green fluorescence as measured in the FL-1 detector with a 530/30 band-pass filter was associated with ROS production and was determined from histogram data using CellQuest software (Becton Dickinson). A total of 20,000 events were collected for each histogram.
Immunocytochemistry for Prx I and caspase 3
A549 cells were planted into six-well plates and randomly divided into four groups: PBS group, scFv group, IR group, and scFv and IR combination group. After different treatments, cells were fixed with 4% paraformaldehyde for 10 minutes, washed three times with PBS buffer, and blocked by 5% BSA at room temperature for 1 hour, and then cells were incubated with Prx I scFv (50 μg/mL in 5% BSA/PBS) or caspase 3 antibody (Santa Cruz Biotechnology) at 4°C overnight. After washing with PBS, cells were incubated with the horseradish peroxidase (HRP)-conjugated anti-E tag monoclonal antibody (LifeSpan BioSciences) or HRP-conjugated anti-mouse monoclonal antibody (Santa Cruz Biotechnology) at room temperature for 1 hour.
Western blot for γ-H2AX and Rad51 protein level
Proteins were extracted from each group with precooled RIPA lysis buffer (50 mM Tris-HCl [pH 7.6], 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM PMSF, 21 μg/mL Aprotinin, and 0.5 μg/mL Leupeptin) in ice bath for 10 minutes after different treatments, followed by centrifugation at 12,000 rpm/min for 10 minutes. Supernatant was collected and protein concentration was measured using Lowry method. Then, protein was mixed with 2×SDS loading buffer (125 mM Tris-HCl [pH 6.8], 20% glycerol, 0.01% bromophenol blue, 4% SDS, and 200 mM DTT), boiled for 5 minutes, separated by 10% SDS polyacrylamide gel electrophoresis with 30 μg each lane. After electrophoresis, proteins were transferred to PVDF membrane and incubated with anti-β-actin antibody (Santa Cruz Biotechnology), anti-γ-H2AX (Upstate Biotechnology), or anti-Rad51 antibody (Santa Cruz Biotechnology), and then with HRE-conjugated secondary antibody. The protein band was detected by ECL assay (Santa Cruz Biotechnology) and the density was analyzed by Quantity One@ (Bio-Rad Laboratories). Target protein density was normalized with β-actin.
Colony-forming assay
We performed colony-forming assay to detect cell radiosensitivity. A549 cells were digested with 0.25% trypsin, pelleted, and resuspended in 1 mL fresh media. Then, single cell suspension was prepared to determine the cell viability by trypan blue dye exclusion method. Cells were planted at a density of 5×105/mL cells/well in six-well dishes in DMEM with 10% fetal bovine serum in a humidified atmosphere with 5% CO2 at 37°C and allowed to attach overnight. After that, cells were treated with PBS or Prx I scFv for 24 hours and then irradiated (0–10 Gy). Ten (10) days after irradiation, cultures were harvested, fixed by methyl alcohol for 15 minutes, washed, and stained with Giemsa staining buffer for 15 minutes, then washed with running water, and dried in air. A population of >50 cells was counted as one colony, and the number of colonies was expressed as a percentage of the number of mock-irradiated cells. The surviving curves were plotted by linear regression analyses. The number of clones was examined by macroscopic observation. Colony-forming efficiency (CFE) was calculated with the following formula: CFE=clone number/number of seeded cells.
Nude mouse xenograft model
The in vivo study was approved by the Animal Study Committee and was conducted in accordance with Chongqing Medical University guidelines and regulations on the use and care of laboratory animals. Female BALB/c nude mice with 4 weeks of age and 16±0.6 g of weight were purchased from Experimental Animal Center of Chongqing Medical University (Chongqing, P.R. China). Mice were housed in microisolator cages in a specific pathogen-free environment with 12-hour light–dark cycles. Mice were subcutaneously implanted with 4×106 A549 cells. Once tumors reached approximately 1×1×1 cm in volume, the mice were randomly assigned into four groups: PBS group, Prx I scFv group, PBS/IR group (PBS combined with IR), and Prx I scFv/IR group (Prx I scFv combined with IR). Mice were injected with PBS or Prx I scFv once every 3 days. Four (4) hours after drug administration, the mice received a dose of 6 Gy irridation to the tumor. Irradiation was performed using a Pantak irradiator with animals restrained in a custom jig. Tumor dimensions were measured once every other day and the tumor volumes calculated using the formula: ½×a×b 2 , where a and b, respectively, represented the largest and smallest tumor diameter. At the end of the treatment, mice were sacrificed by overdose of ketamine (400 mg/kg) and xylazine (50 mg/kg) and then necropsy was performed.
Statistical analysis
All data are presented as means±SEM. Data were statistically analyzed by Student's t-test with the use of Prism 4.0 (GraphPad Software Inc). *p<0.05 was considered to be statistically significant.
Results
Prx I protein expression was significantly decreased in A549 cells after Prx I scFv treatment
We have successfully produced scFv that highly specific against Prx I of lung adenocarcinoma and found that it could reduce Prx I protein level in A549 cells. To evaluate the effect of scFv, IR, or combination of scFv and IR on Prx I protein level, we performed immunocytochemistry in A549 cells of each group. For the Prx I scFv group, A549 cells were incubated with 100 nM Prx I scFv for 72 hours; for the scFv and IR group, A549 cells were incubated with 100 nM Prx I scFv for 72 hours, followed by 6 Gy radiation. The same dose of Prx I scFv and radiation were used in the following studies.
The positive staining for Prx I was in orange or brown, scattered or mottled within the cytoplasm of A549 cells. Compared with PBS group, we found that Prx I intensity was significantly decreased in Prx I scFv group, while increased in IR group. Combined treatment of scFv and IR abolished the increase of Prx I induced by IR (Fig. 1). Since decreased Prx I expression by siRNA treatment could enhance the radiosensitivity of cancer cells, we hypothesized that Prx I scFv could also enhance the inhibitory effect of IR on A549 cells growth.
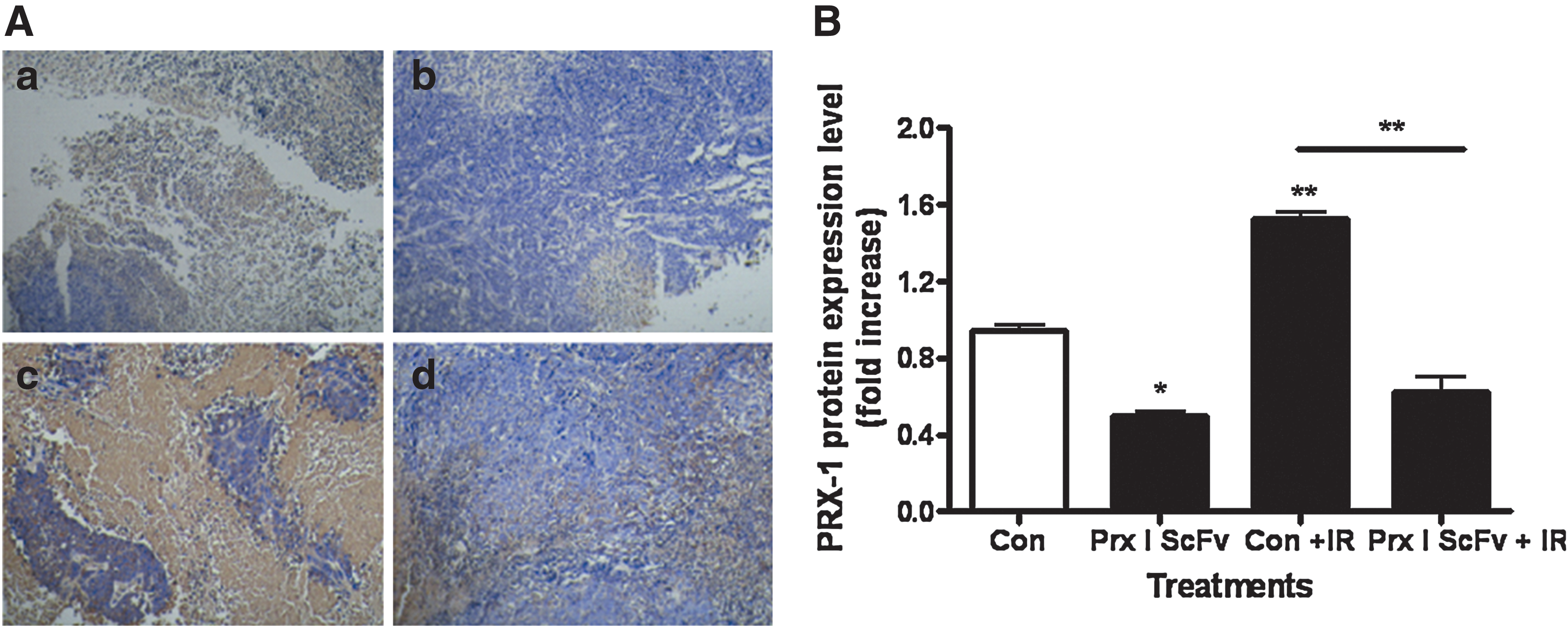
Peroxiredoxin I (Prx I) protein expression was decreased after treatment with Prx I single-chain variable fragment (scFv). Immunocytochemistry was performed to evaluate Prx I protein expression in A549 cells after different treatment
The inhibitory effect of IR on A549 cells growth was enhanced after pretreatment with Prx I scFv
Previous studies in our lab have demonstrated that Prx I scFv exhibited a dose-dependent growth inhibitory effect on A549 cells. We wondered whether preincubation of Prx I scFv could enhance the inhibitory effect of IR on A549 cells growth. A549 cells were treated with different doses of IR from 0 to 10 Gy after incubated with Prx I scFv or PBS. Then, cell proliferation was assessed by colony-forming assay. IR treatment exhibited a dose-dependent growth inhibition effect on A549 cells, and preincubation with Prx I scFv could enhance this effect, suggesting that Prx I scFv could increase A549 cell radiosensitivity (Fig. 2). Furthermore, we also carried out the same experiments on SPCA-1 (pancreatic cancer) and H446 (small cell lung cancer) cell lines and got similar results as A549 cancer cell line, indicating that the enhancement of Prx I scFv on IR-induced cell death is an universal phenomenon (Suppl. Fig. S1; Supplementary data is available online at
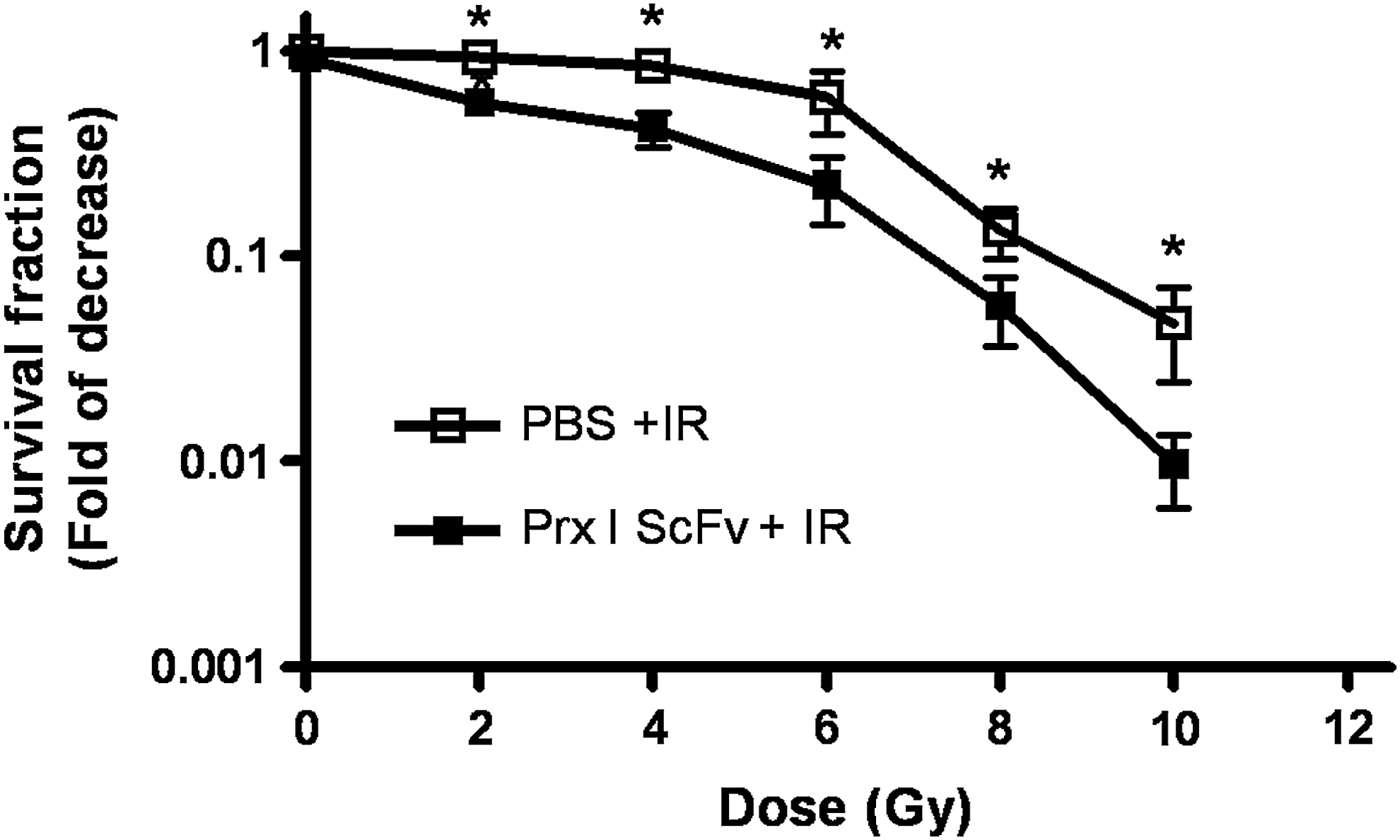
The inhibition effect of IR on A549 cells growth was enhanced after pretreatment with Prx I scFv. A549 cells were pretreated with 0, 2, 4, 6, 8, and 10 Gy irradiation alone or in combination with Prx I scFv and survival curves were determined by colony-forming assay. Student's t-test was used to compare the difference of cell survival fractions between the two groups. A549 preincubated with Prx I scFv were more radiosensitive than PBS-preincubated cells. *p<0.01 vs. control group, n=3.
The effect of IR on inducing cell apoptosis rate was enhanced after pretreatment with Prx I scFv
As Prx I scFv could increase A549 cell radiosensitivity, by using flow cytometry we further confirmed that A549 cell apoptosis rate was also increased. Results showed cell apoptosis rate of Prx I scFv with IR group was significantly higher than PBS with IR group, Prx I scFv group, and PBS group (Fig. 3), suggesting that the increased radiosensitivity may result from increased apoptosis rate.
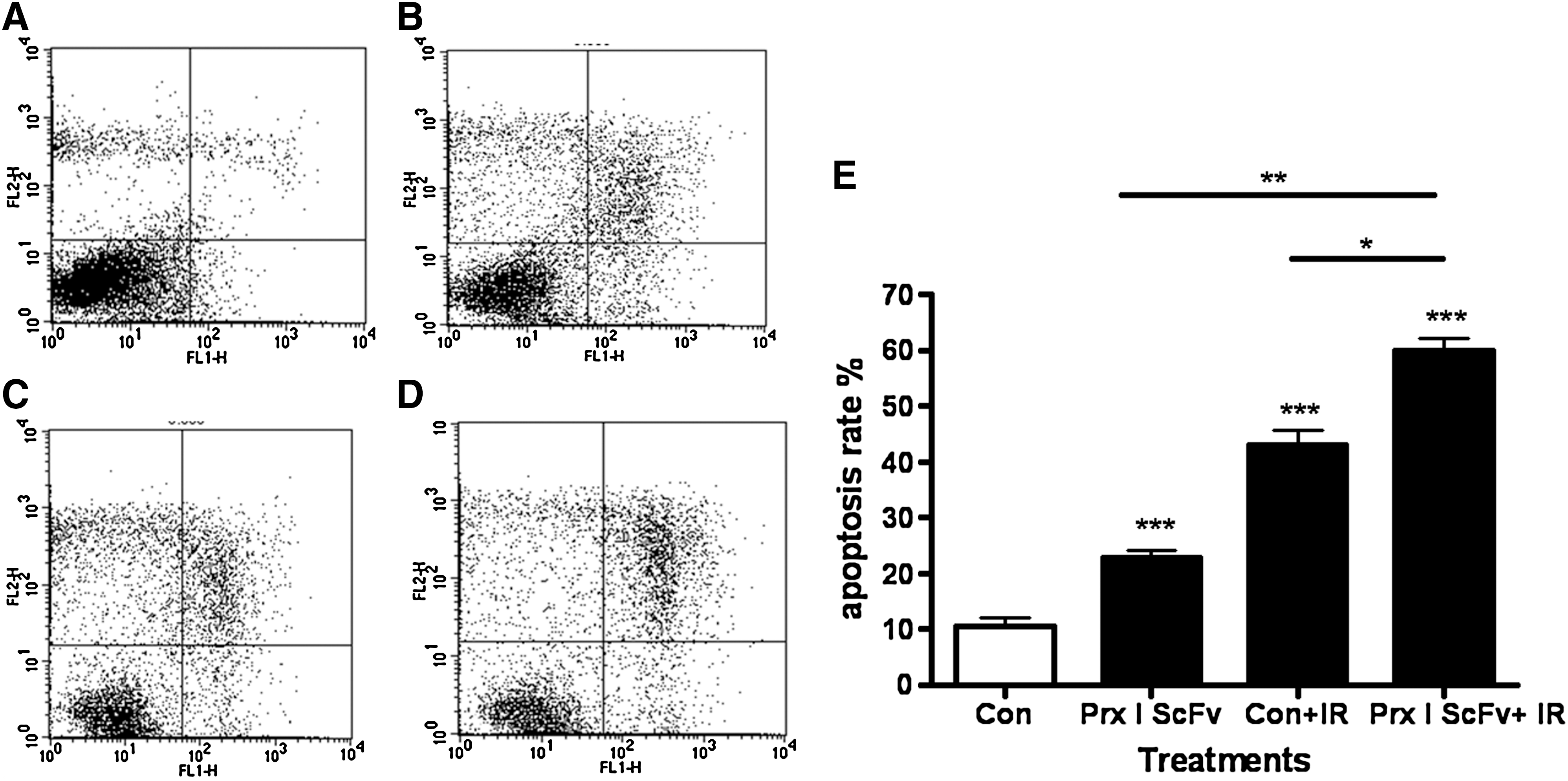
The effect of IR on increasing cell apoptosis rate was enhanced after combined treatment with Prx I scFv. Flow cytometry was used to measure cell apoptosis rate in A549 cells after different treatment. Representative FCM images for cell apoptosis rate in the A549 cells after PBS
The effect of IR on increasing cellular caspase 3 expressions was enhanced after pretreatment with Prx I scFv
As caspase 3 is an important apoptosis-inducing kinase, we wondered whether caspase 3 played a role in the enhanced cell apoptosis when cells were treated with IR and Prx I scFv. We performed immunocytochemistry to detect caspase 3 expression in A549 cells. The positive staining for caspase 3 was in orange or brown in the cytoplasm and nucleus of A549 cells (Fig. 4). Compared with PBS group, expression of caspase 3 was significantly increased in the Prx I scFv group, the IR group, and especially in the Prx I scFv with IR group (Fig. 4). Statistical results demonstrated that the effects of IR with Prx I scFv group differ significantly from IR or Prx I scFv group, indicating that the increased apoptosis effect of Prx I scFv with IR may through increased caspase 3 protein expression.

The effect of IR on increasing intracellular caspase 3 expression was enhanced after combined treatment with Prx I scFv. Immunocytochemistry was performed to evaluate caspase 3 expression in A549 cells after different treatment.
The effect of IR on increasing intracellular ROS was enhanced after pretreatment with Prx I scFv
Tumor cells with upregulated Prx I displayed stronger capability of antioxidation and lower intracelluar ROS level, which played a key role in antiapoptosis and drug resistance of tumor cell. As Prx I scFv could effectively reduce Prx I protein level and enhance cell apoptosis, we wondered the increased radiosensitivity may result from increased ROS level by Prx I, and measured intracellular ROS of A549 cell by FCM. Results showed that compared with PBS group, ROS level was significantly increased in the other three groups, especially in the combined treatment of scFv and IR group (Fig. 5).

The effect of IR on increasing intracellular reactive oxygen species (ROS) was enhanced after combined treatment with Prx I. FCM was performed to evaluate intracellular ROS level in A549 cells after different treatments.
The effect of IR on promoting cells entering G1 phase was enhanced after pretreatment with Prx I scFv
Since cells in the G1 and G2 stage are sensitive to IR, we performed cell cycle analysis to detect whether the cell cycle changed when cells were treated with IR and Prx I scFv. Cells received IR alone, Prx I scFv alone, or combination treatment with IR and Prx I scFv displayed G1 stage arrest, and the percentage of cells in the G1 stage was increased, whereas that of S stage was reduced. Among the four groups, the Prx I scFv/IR group could regulate cell cycle most significantly (Fig. 6 and Table 1). These results suggested that the enhanced radiosensitivity of A549 cells may through the changed cell cycle induced by Prx I scFv administration.
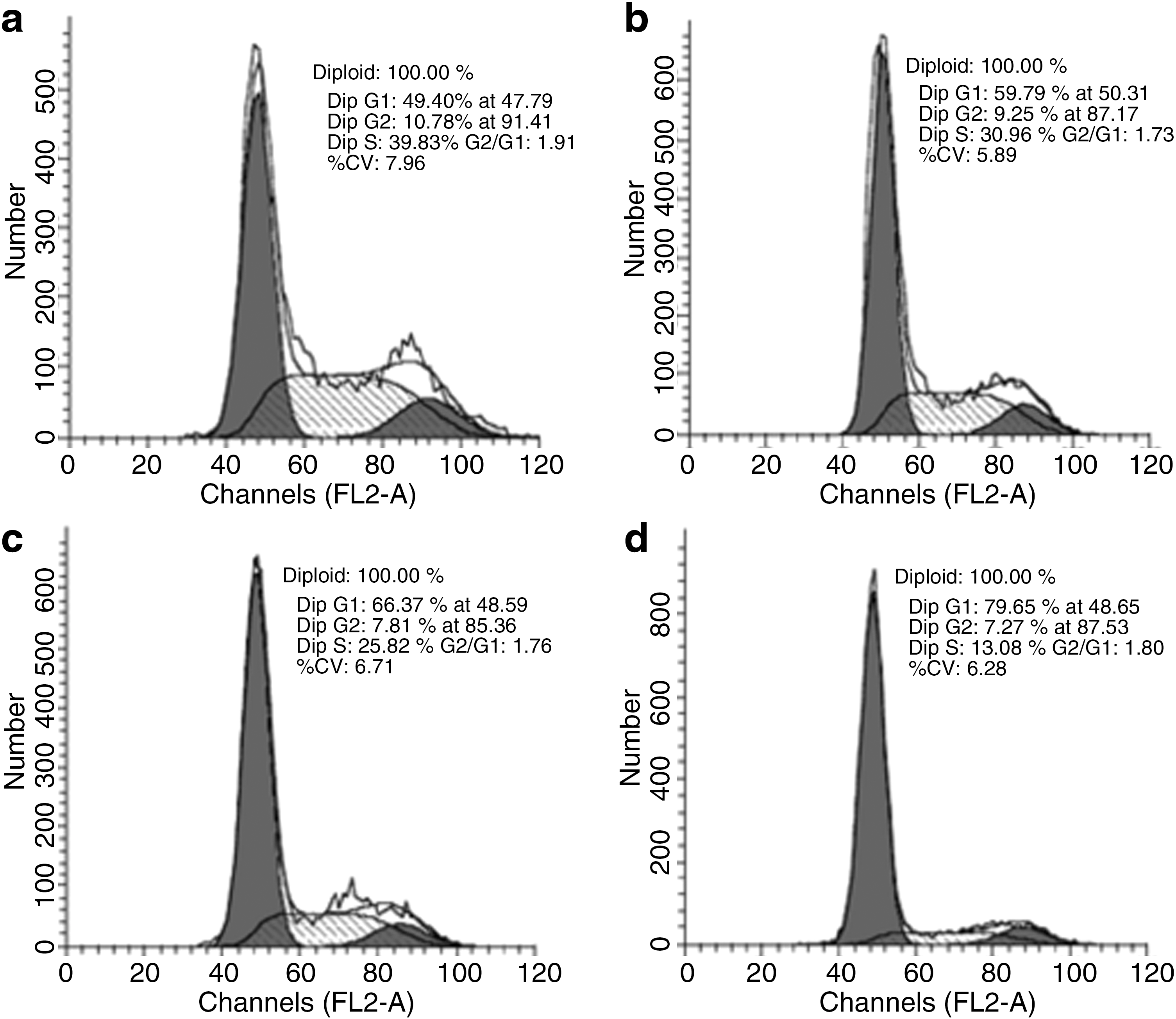
The effect of IR on increasing number of cells entering G1 phase was enhanced after combined treatment with Prx I scFv. Flow cytometry was performed to evaluate cell cycles in A549 cells after different treatments. Shown are representative FCM images for cell cycles in A549 cells after PBS
Percentage of cells in G1 stage was counted. Values represent means±SEM of three independent experiments. Percentage of cells that in G1 stage significantly increased in Prx I scFv, IR, Prx I scFv + IR groups, especially in Prx I scFv + IR group.
p<0.05 vs. control group, b p<0.05 vs. Prx I scFv + IR, c p<0.01 vs. Prx I scFv + IR, d p<0.01 vs. control group, n=3.
IR, ionizing radiation; Prx I, peroxiredoxin I; scFv, single-chain variable fragment.
The effect of IR on decreasing cellular RAD51 expression was enhanced after pretreatment with Prx I scFv
RAD51, which is involved in recombination and repair of damaged DNA, is found to be negatively correlated with tumor radiosensitivity. We performed Western blot to detect the RAD51 expression in A549 cells after different treatments. Compared with the PBS group, the RAD51 protein level was significantly decreased in the other three groups, especially in the IR/scFv (combined IR and scFv treatment) group (Fig. 7). Statistical results indicate that the RAD51 protein level of IR/Prx I scFv group differed significantly from IR or Prx I scFv alone group (Fig. 7), suggesting that decreased RAD51 expression might be one of the underlying mechanisms of the increased radiosensitivity induced by Prx I scFv administration.
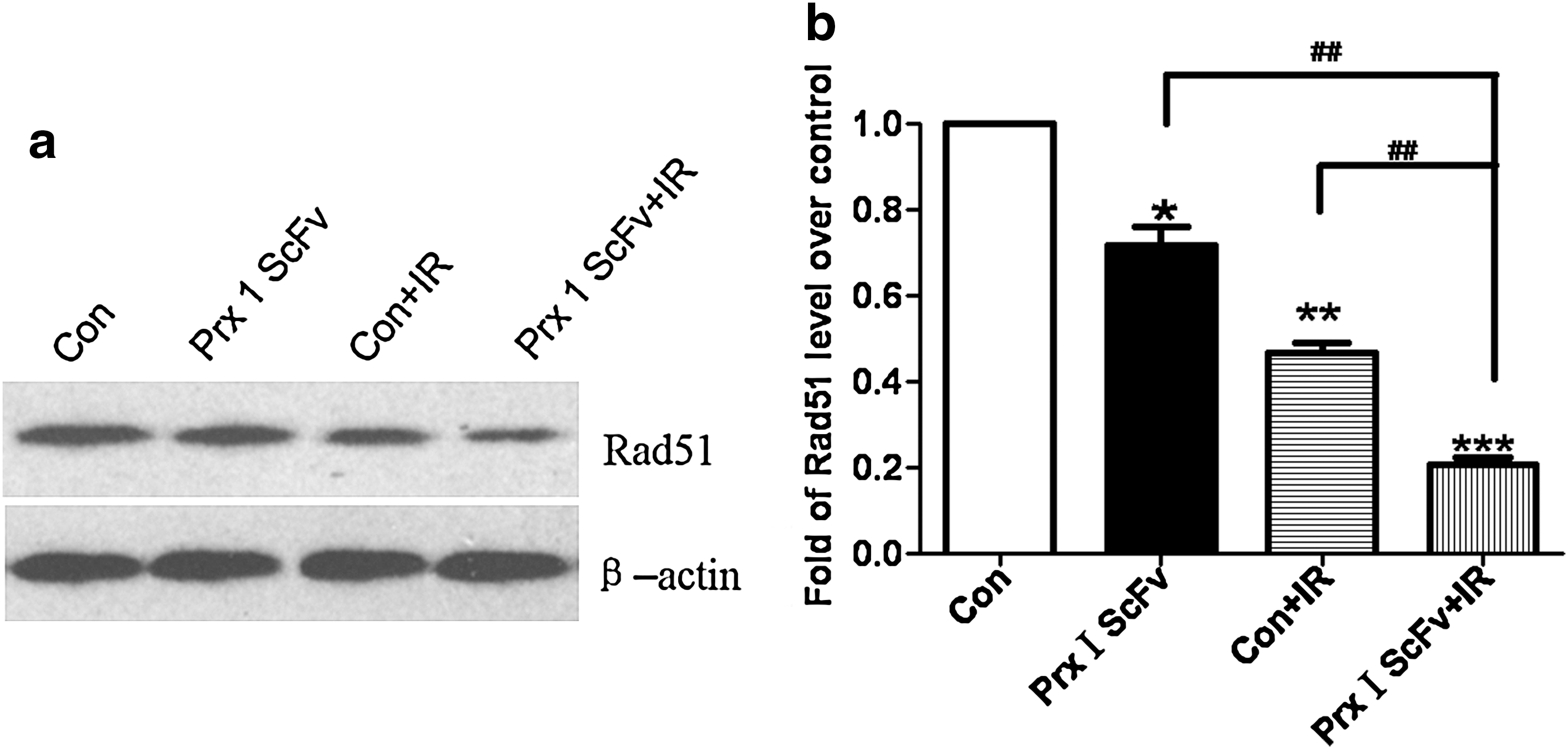
The effect of IR on decreasing cellular RAD51 expression was enhanced after combined treatment with Prx I scFv. Western blot was performed to evaluate RAD51 protein expression in A549 cells after different treatments.
The effect of IR on increasing cellular γ-H2AX expression was enhanced after pretreatment with Prx I scFv
γ-H2AX has been found positively related with tumor radiosensitivity. We performed Western blot to detect the γ-H2AX expression in A549 cells after different treatments. Compared with the PBS group, γ-H2AX protein level was significantly increased in the other 3 groups, especially in the combined treatment of scFv and IR group (Fig. 8). Statistical results indicated that γ-H2AX protein level of IR with Prx I scFv differ significantly from IR or Prx I scFv alone, suggesting that increased γ-H2AX expression might be one of the underlying mechanisms of the increased radiosensitivity induced by Prx I scFv administration.

The effect of IR on increasing cellular γ-H2AX expression was enhanced after combined treatment with Prx I scFv. Western blot was performed to evaluate γ-H2AX protein expression in A549 cells after different treatments.
Prx I scFv administration increased A549 radiosensitivity in vivo
As we had confirmed that Prx I scFv could effectively inhibit A549 radiosensitivity by increasing its apoptosis, next we want to know whether this treatment effect could appear in in vivo experiments. First we performed Western blot and confirmed Prx I scFv could effectively decrease Prx I protein level in vivo (data not shown). Then, we established nude mouse xenograft model and measured tumor growth continuously. Tumor growth curves were drawn to evaluate effects of Prx I scFv on tumor radiosensitivity. Results showed that tumor volume did not have significant difference in the four groups before treatments. Seven days after IR treatment, tumor volume of Prx I scFv/IR group reached 1.13±0.15 cm3, whereas that of PBS/IR group reached 1.39±0.17 cm3, and the difference reached statistical significance. Tumor volume of Prx I scFv/IR group was significantly smaller (p<0.05) (Fig. 9) from the 7th day after IR treatment until the 15th day, indicating that injection of Prx I scFv could enhance tumor radiosensitivity in vivo.
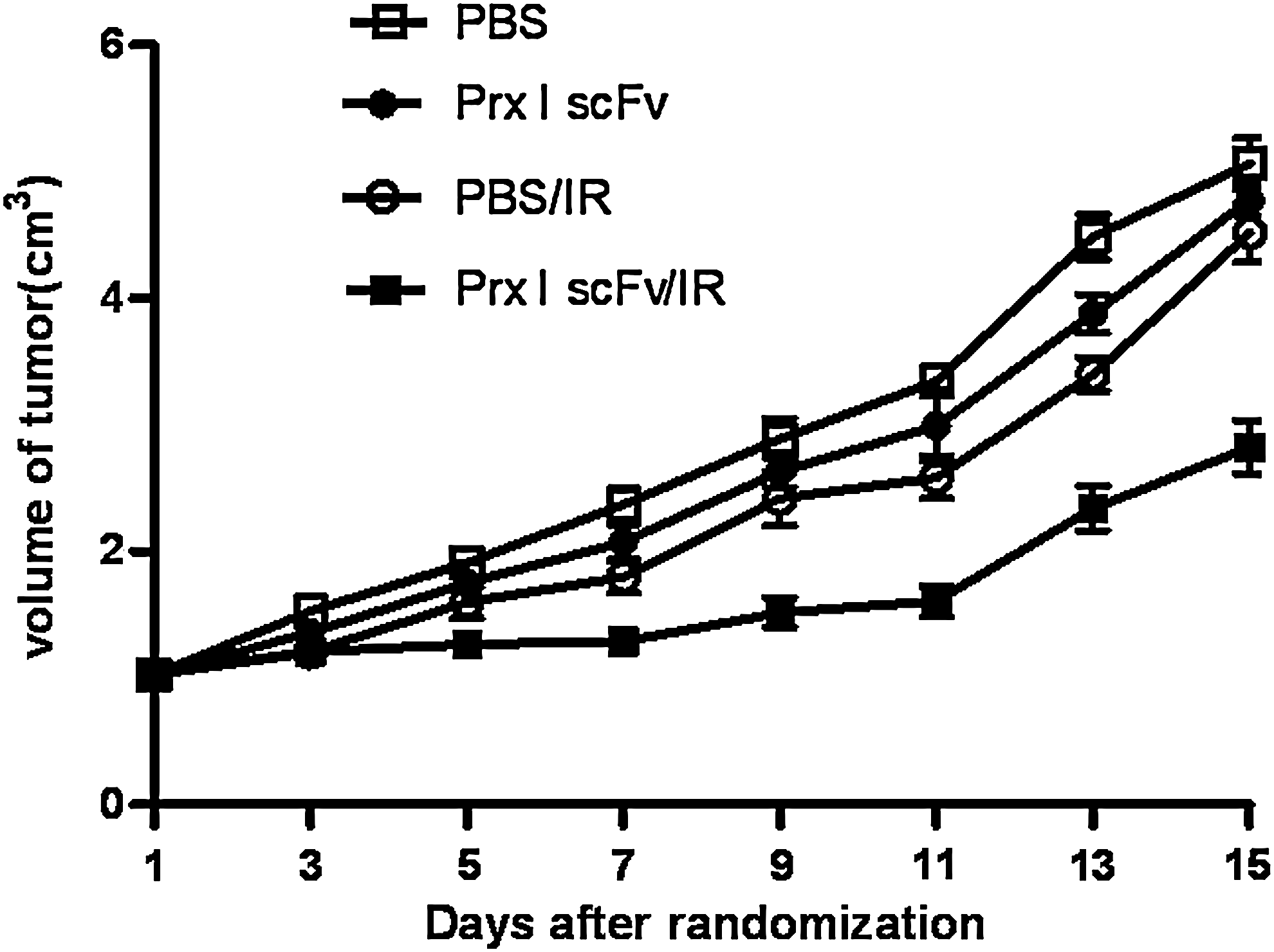
The effects of Prx I scFv on A549 xenograft tumors. When A549 tumors reached 1 cm3 in size, mice were randomized allocated into four groups: PBS, IR, PBS/IR, or Prx I scFv/IR. Mice were given PBS or Prx I scFv once every 3 days. Radiation (6 Gy) was delivered 4 hours after PBS or Prx I scFv treatment. n=3, Points, mean; bars, SE.
Discussion
Prx I, which belongs to a special antioxidant Prxs family, was expressed at a high level in carcinoma cells and was considered to be a potential target for cancer treatment. 4 The physiological role of Prx has been explored. Notably, Prx I has been shown to be a significant contributor to genome stability; its absence leads to a broad spectrum of spontaneous mutations and genome rearrangements that are thought to result from increased oxidative damage to the DNA. 11 Importantly, mutations affecting recombinational repair including Rad51 or Rad52, or the Rad 6-mediated postreplication repair pathway including Rad6, cause Prx I knockout cells to be very sick or to die rapidly under aerobic conditions but not anaerobic conditions, emphasizing the importance of Prx I for genome protection. 12 Furthermore, Prx I has a specific protective role toward peroxides produced during respiration, consistent with its mitochondrial localization. 13,14
We have developed Prx I scFv targeting Prx I and found that scFv incubation resulted in downregulation of Prx I expression. 10 In the present study, we investigated the effect of this novel Prx I scFv on human lung carcinoma cell line A549 radiosensitivity and the underlying mechanisms. After incubated with Prx I scFv, the Prx I protein level was decreased. Furthermore, consistent with previous studies, 15,16 we also found that 6 Gy IR could induce an increase of Prx I expression, whereas combination treatment with Prx I scFv successfully blocked this elevation.
Then, we found downregulation of Prx I expression by Prx I scFv resulted in significant growth inhibition when treated with IR, demonstrating that Prx I scFv could enhance A549 cell radiosensitivity, which was further confirmed by flow cytometric analysis of cell death.
Caspases play an important role in apoptosis, and most of the stimuli inducing apoptosis were through the caspase protein cascades. We observed that combination treatment with Prx I scFv could increase caspase 3 expression compared with IR alone, suggesting that caspase 3 activation might contribute to the increased cell death. By detecting intracellular ROS, we also found that Prx I scFv had not only greater intracellular ROS than wild-type cells but also a greater level of induced ROS after IR treatment. IR evokes cell oxidative stress response both directly and indirectly, and ROS induced by IR involved in cell death especially in apoptotic cell death, which were widely investigated in the field of cancer radiotherapy, and the deleterious effects by ROS include DNA damage and membrane oxidative damage. 17 Since numerous studies have found that Prx I could reduce ROS and protect DNA from ROS-induced damages (review in Refs. 18,19 ), it is meaningful that blockade of Prx I protein expression could elevate ROS level.
When analyzing cell cycle, we found that Prx I scFv could promote cells to enter G1 phase. Since cells in the G1 and G2 stage are more sensitive to IR, decreased Prx I expression might enhance cell radiosensitivity by regulating cell cycle. Although the detailed mechanisms was still poorly understood, it seemed that the cell cycle arrested effect of Prx I scFv probably conducted by deletion of Prx I cyc47, which was found to be the active site of Prx I and could protect cell go through the cell cycle check points in a recent study. In the same study, author also found deleted this active site could certainly arrested cell cycle at G1 phase. 20 As the cyc47 site was important for Prx I peroxidatic function, we speculated that the cell cycle arrested effect induced by Prx I scFv may conducted by losing protection to IR-induced ROS damages.
We performed Western blot to explore whether the expression of Rad51 and γ-H2AX that involved in DNA repair changed after IR and Prx I scFv treatment alone or in combination. Rad51, with structural and functional similarity to RecA protein, plays an important role in recombination repair, which is a key mechanism for repair of double-strand DNA breaks. 21 We observed that A549 cells not only exhibited decreased Rad51 protein expression after IR or Prx I scFv treatment alone but also displayed a decreased level after IR and Prx I scFv treatment in combination, which is consistent with previous studies, indicating that Rad51 negatively correlated with cancer radiosensitivity. 22,23 In contrast to the decreased Rad51 protein expression, γ-H2AX protein increased especially after IR and Prx I scFv treatment in combination. γ-H2AX is considerded as a biomarker for DNA double-strand breaks and has a positive association with radiosensitivity. 24 Our results demonstrated that Rad51 and γ-H2AX might play important roles in the enhanced radiosensitivity by Prx I scFv incubation.
In the present study, we found that downregulation of Prx I protein expression by Prx I scFv could increase A549 cell radiosensitivity, suggesting that inactivation of Prx I might be a promising approach to improve the treatment of lung cancer. Some literature has explored the possibility of anticancer therapy based on decreasing Prx I expression including RNA interference 8 and antisense strategies 25 and found that decreased Prx I expression could enhance the radiosensitivity of cancer cells.
In addition to RNA interference and antisense strategies, it has now been confirmed that combined therapy of monoclonal antibody and radiotherapy was effective for many types of cancer. 26,27 However, monoclonal antibodies generated from hybridoma technique are almost murine antibodies, which are not suitable for clinical application due to the generation of human anti-murine antibody (HAMA) and low blood serum clearance that raises potential therapy risk. 28,29 Besides, productions of monoclonal antibodies are laborious and expensive, 30,31 which prevented its clinical application.
The search for human scFv against cell molecules that may be used as targets for cancer therapies has recently led to the development of several novel technologies that efficiently generate such antibodies. Phage display is one such technology that, with the development of improved selection and screening methods, has proved to be very useful and is an attractive method that avoids the deficiencies of hybridoma technology, generating antibodies from both immune and nonimmune sources. 32,33 Fully human antibodies offer the greatest potential for success as human therapeutics because they would be less likely to elicit an immune response and should possess a longer half-life in vivo. 34
In conclusion, Prx I scFv could increase radiosensitivity of human being carcinoma A549 cell through specifically inhibition of Prx I and induce apoptosis of lung cancer cells. Combination therapy of Prx I scFv and radiotherapy may be a potential and promising strategy for curing lung cancer.
Footnotes
Acknowledgments
This work was supported by research grant from the National Natural Science Foundation of China (No. 30370422).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.