
Abstract
The 5-year survival rate of nasopharyngeal carcinoma (NPC) is disappointing despite the much improved technologies in its treatment. Thus, finding more effective treatment for NPC has become an urgent priority. The tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) can selectively induce apoptosis in most tumor cells while sparing normal cells. However, its potential in the treatment of NPC has been limited by the eventual emergence of drug resistance. Bcl-2 and Akt contribute to TRAIL resistance in some cancer cells. In this study, CNE-2 was found to be the most resistant NPC cell line to TRAIL, and whether Bcl-2 small-interfering RNA (siRNA) and phosphatidylinositol 3-kinase (PI3-K) inhibitors (LY294002 and Wortmannin) could prevent TRAIL resistance in CNE-2 was also investigated. Results showed that both Bcl-2 siRNA and PI3-K inhibitors could prevent TRAIL resistance in CNE-2. Bcl-2 siRNA sensitized CNE-2 by activating the intrinsic apoptotic pathway and PI3-K inhibitors sensitized CNE-2 by activating both intrinsic and extrinsic pathways. Further, simultaneously targeting Bcl-2 and Akt was found to be a more efficient approach to prevent TRAIL resistance in CNE-2 and this synergistic effect happened at the level of Bid downstream. At last, the combinative treatments did not enhance toxicity of TRAIL in MRC5, a human benign fibroblast cell line. This study suggests that simultaneously targeting Bcl-2 and Akt pathway might be effective in preventing TRAIL resistance of NPC cells.
Introduction
Tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) is a cytokine of the TNF family and capable of inducing apoptosis in a variety of cancer cells without having significant effects on normal cells. 1 TRAIL induces apoptosis by binding to their cognate receptors, TRAIL-R1 (death receptor 4, DR4) and TRAIL-R2 (DR5). 2 The Fas-associated death domain and caspase-8 are recruited to the DR-TRAIL complex to form a death-inducing signaling complex (DISC). 2 The apoptotic signal is then transmitted through the extrinsic and intrinsic pathways. The extrinsic pathway is mitochondrion independent but the intrinsic pathway is mitochondrion dependent. 3 Signals from DISC are linked to mitochondria by Bid, which contains a homologous domain of the Bcl-2 protein family. Bid is cleaved by caspase-8 and then translocates to mitochondria to activate Bcl-2 family members Bax and Bak, leading to mitochondrial depolarization. 4 Apoptotic factors are consequently released from mitochondria into cytosol to activate caspase-9 and -3 through formation of apoptosome complex. 4 Studies have shown that some cancer cells are resistant to the apoptotic effect of TRAIL, the so-called TRAIL resistance. 5 However, TRAIL resistance can be overcome by combining TRAIL with different chemotherapeutic drugs, 6 indicating that an approach by combining multiple targets can be utilized to prevent or reverse TRAIL resistance in cancer cells. Thus, studying the factors that cause TRAIL resistance may be an important way to find more effective therapies for cancer.
Nasopharyngeal carcinoma (NPC) has a distinct racial and geographical distribution. It is common in Southeast Asia and southern area of People's Republic of China but is rare in most other parts of the world. 7 Most patients with NPC are diagnosed at an advanced stage. Radiotherapy alone was the standard treatment for NPC at all stages until 1990. 8 Despite significant progress in methods of radiotherapy, the outcome with radiotherapy alone has been disappointing. The 5-year survival rate is between 34% and 52%. 9 Thus, finding more effective treatments for NPC has become an urgent priority. Death receptors have been detected in specimens of head and neck tumors but not in surrounding normal tissues. 10 These findings support the notion that the presence of death receptors on some head and neck cancers may make them more susceptible to the TRAIL-mediated apoptosis. Some have shown robust induction of apoptosis in NPC cell line TW02 by TRAIL, 11 whereas others had difficulty in inducing apoptosis in some other NPC cell lines by TRAIL. 12 Activation of Bcl-2 and Akt pathway is known to reduce chemosensitivity of NPC cells. Yin et al. 13 showed that suppression of Bcl-2 gene by RNA interference increased chemosensitivity to cisplatin in NPC cell line CNE-1. Zhang et al. 14 found that the resistance to taxol in NPC cell line HNE1 was associated with activity of Akt pathway. Bcl-2 and Akt also play important roles on TRAIL resistance in some cancer cells. It has been shown that Bcl-2 overexpression inhibits TRAIL-induced apoptosis in many kinds of cancer cells. 15,16 Activation of phosphatidylinositol 3-kinase (PI3-K)/Akt pathway can protect cells from the TRAIL-induced apoptosis, and blockade of the PI3-K/Akt pathway sensitizes some cancer cells to the TRAIL-induced apoptosis. 17
In this study, the combinational approaches were tested to reactivate TRAIL apoptosis in resistant NPC cells by inhibition of Bcl-2 expression and Akt activity, and whether simultaneously targeting Bcl-2 and Akt was a more efficient approach to prevent TRAIL resistance was also explored. The molecular mechanisms that mediated activation of extrinsic and intrinsic apoptotic pathways were also examined.
Materials and Methods
Cell lines and reagents
Four human NPC cell lines (CNE-1, CNE-2, 5-8F, 6-10B) were used in this study. CNE-1 is a well-differentiated squamous cell carcinoma NPC cell line, and CNE-2 is a poorly differentiated cell line. 5-8F is a highly metastasized cell line, and 6-10B is a poorly metastasized cell line. All NPC cells were kindly provided by the Department of Dtolaryngology and Head Neck Surgery Xiangya Second Hospital in Central South University. MRC5, a benign human fibroblast cell line, was kindly provided by the Cell Research Center of Central South University. They were grown in RPMI-1640 media with supplemented 10% fetal bovine serum and penicillin/streptomycin and maintained in a humidified atmosphere of 5% CO2 at 37°C. TRAIL and PI3-K inhibitor (LY294002 and Wortmannin) were purchased from Pharmacia Corporation/Pfizer, Inc., and G. D. Searle & Co., respectively. The drugs were dissolved in dimethylsulfoxide (DMSO) at an appropriate concentration and stored at −20°C until use.
Cell viability assay
NPC cells were plated into 96-well plates (3×103 cells/well) in 100 μL of medium. Twenty-four (24) hours later, cells were treated with drugs or not and incubated for 24 hours. Cell viability was assessed by MTT assay. Ten microliters of 5 mg/mL MTT solution (Sigma) was added to each well. After 3 hours of incubation, 200 μL of DMSO (Fisher Biotech) was added to each well. The absorbance of each well was measured at 570 nm with a microplate reader (Viability=ODtreat/ODcontrol).
Flow cytometric analysis
Cells were incubated with TRAIL at a concentration of 100 ng/mL. After 24 hours, cells were harvested. The cell pellets were suspended in 500 μL binding buffer (10 mM HEPES [pH 7.4], 140 mM NaCl, 1 mM MgCl2, 5 mM KCl, and 2.5 mM CaCl2) at a density of ∼1×106 cells/mL. Samples were incubated with 1 μL annexin V-FITC and 5 μL PI for exactly 5 minutes at room temperature in the dark and then measured on a FACSCalibur cytometer (Becton Dickinson). Annexin V-FITC and PI fluorescence were detected in the FL-1 (green) and FL-2 (red) channels, respectively, after correction to the spectral overlap between the two channels. Data were analyzed using CellQuest software (Becton Dickinson).
Western blotting
Cells were washed with phosphate-buffered saline (PBS) and dissolved in lysis buffer (20 mM Na2PO4, 150 mM NaCl, 1% Triton X-100, 1% aprotinin, 1 mM phenylmethylsulfonyl fluoride, 100 mM NaF, and 2 mM Na3VO4). Proteins were separated by polyacrylamide gel electrophoresis on a sodium dodecyl sulfate–polyacrylamide gel and transferred to a polyvinylidene difluoride membrane. Membranes were incubated with the first antibody overnight at 4°C. On the next day, the membranes were washed with PBS three times and then membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit secondary antibodies (all the antibodies were obtained from Santa Cruz Biotechnology). Finally, proteins were detected by the enhanced chemiluminescence procedure.
Bcl-2 small-interfering RNA transfection
pSilencer 3.1-H1 linear vector was purchased from Invitrogen Corporation. The Bcl-2 small-interfering RNA (siRNA) insert sequence with the following sense and antisense sequences were for Bcl-2 sense 5′-AGT ACA TCC ATT ATA AGCT-3′ and antisense 5′-AGCTTATAATGGATGTACT-3′. Negative control vector that expresses a hairpin siRNA with limited homology to any known sequences in the human was provided by vector kit. Plasmid DNA was purified by CsCl–ethidium bromide gradient centrifugation. The purified DNA was diluted to 1 g/L and frozen at −20°C. The transfection of siRNA was performed using Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. In brief, the cells were seeded into wells of a six-well plate at 105 cells/cm2 and cultured for 24 hours until 60%–70% confluence. Then Bcl-2 siRNA or negative siRNA plasmid (4 μL) was diluted with 100 μL OPTI-MEM (Invitrogen) for 5 minutes at room temperature. During this incubation period, 5 μL Lipofectamine 2000 (Invitrogen) was diluted in 100 μL OPTI-MEM. These two mixtures were combined, mixed gently, and incubated for 20 minutes at room temperature for complex formation. This 200 μL siRNA–Lipofectamine 2000 mixture was then added into the cells. The transfected cells were cultured for 48 hours and then they were assayed.
Assessment of mitochondrial transmembrane potential
Assessment of mitochondrial transmembrane potential (Δψm) by the cationic lipophilic fluorochrome 3,3′-dihexyloxacarbo-cyanide iodide (DiOC6(3)) (460 ng/mL; Molecular Probes) was used to measure the mitochondrial transmembrane potential. Cells were treated for 24 hours with TRAIL (100 ng/mL), then incubated for 30 minutes at 37.8°C in the media with DiOC6(3), washed twice with PBS, and analyzed in a FACSort flow cytometer.
Statistical analysis
All tests were done three times and statistical analysis was performed on SPSS 15.0 statistical software. Descriptive statistics used means±SD and one-way analysis of variances was used to analyze significant differences. A p-value of <0.05 was considered to be statistically significant.
Results
The sensitivity to TRAIL varied among NPC cells
Effect of TRAIL on cell survival in NPC cells was evaluated using MTT cell viability assays. As shown in Figure 1A, treatment with TRAIL caused cell death in CNE-1, 5-8F, and 6-10B cells in a dose-dependent manner but had little effect on of CNE-2 cells. Cells were treated with 100 ng/mL TRAIL and examined by flow cytometry analysis for apoptotic rate. TRAIL induced most significant apoptosis in CNE-1 cells but caused a very minimal level of apoptosis in CNE-2 cells (Fig. 1B).
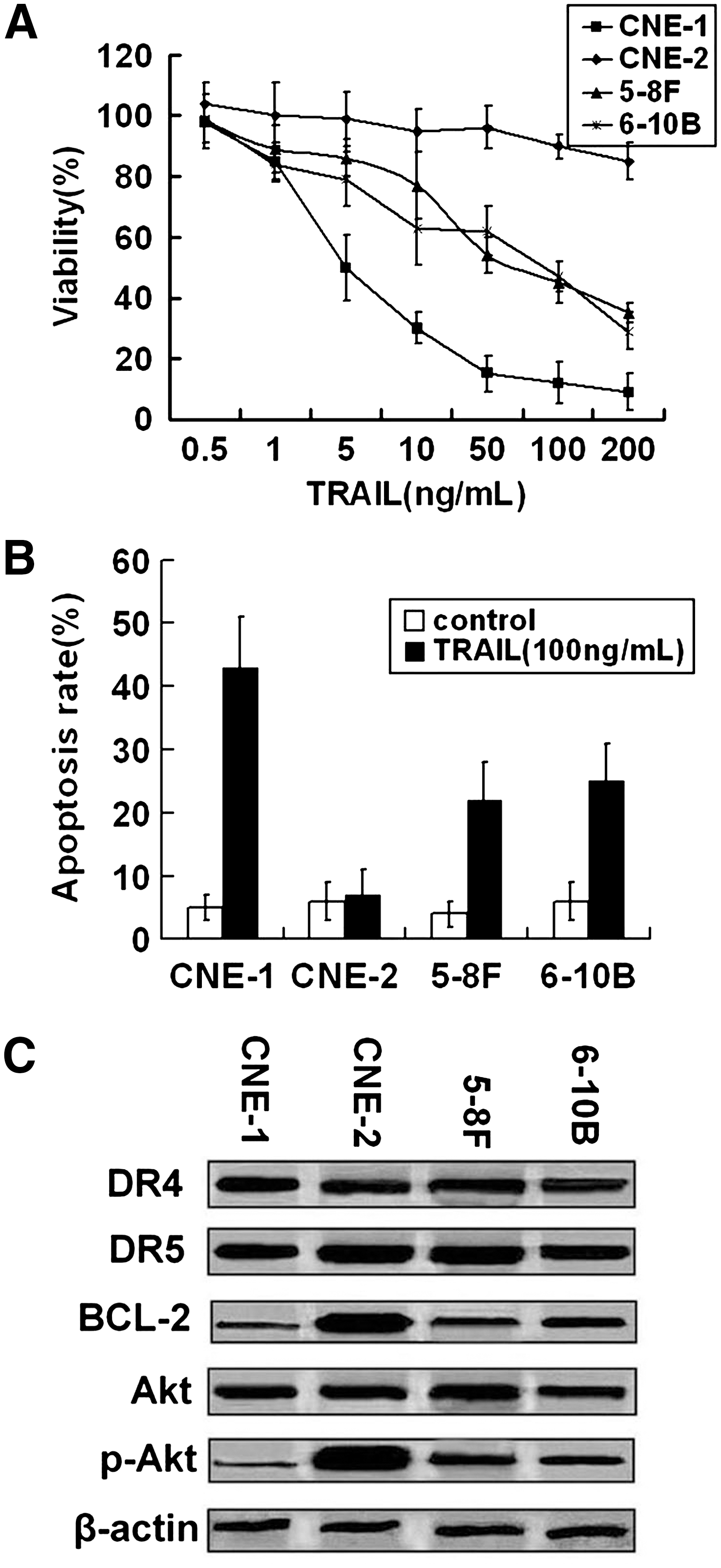
The sensitivity to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) varied among nasopharyngeal carcinoma (NPC) cells.
To examine the effect of protein expression levels of death receptors, Bcl-2, and active Akt on TRAIL resistance, cell extracts from untreated cell lines were examined by western blot analysis. Total Akt, DR4, and DR5 expression levels in these cells were quite similar. The expressions of Bcl-2 and p-Akt were highest in CNE-2, whereas they were lowest in CNE-1, which is most sensitive to TRAIL (Fig. 1C). These results suggested a possible association between baseline levels of Bcl-2, p-Akt, and TRAIL sensitivity.
Effect of Bcl-2 siRNA on TRAIL-induced apoptosis in NPC cells
To study the mechanism of TRAIL resistance in NPC cells, the most resistant cell line, CNE-2, was selected to perform the next experiments. As shown in Figure 2A, Bcl-2 was efficiently knocked down by specific siRNA without effects on expression of DR4, DR5, Akt, and p-Akt. The results of flow cytometry analysis showed that knockdown of Bcl-2 enhanced TRAIL-induced apoptosis (Fig. 2B, C).

Effects of Bcl-2 knockdown on sensitivity to TRAIL in CNE-2.
The cleavage of caspase-8 is the most upstream event in TNF-α family-induced extrinsic apoptotic pathway. Responding to TRAIL treatment, caspase-8 is cleaved to two subunits: p43/p41 and p18. 18 FLIP is thought to have homology to caspase-8 and to be able to inhibit the cleavage of it. 19 As shown in Figure 2C and D, cleavage of caspase-8 and expression of FLIP could not be affected by Bcl-2 siRNA. The intrinsic apoptotic pathway is another mechanism of TRAIL-induced apoptosis. Caspase-9 and -3 are the primary death effecters of the mitochondrial pathway. 20 The present study found that the cleavage of caspase-9 and -3 was enhanced by Bcl-2 siRNA (Fig. 3A). To determine the caspase-9 and -3 upstream molecular mechanism by which Bcl-2 siRNA prevented TRAIL resistance in NPC cells, the effect of Bcl-2 siRNA on cleavage of Bid and Δψm was examined. The Bid cleavage was assessed as a reduction in whole Bid protein, because the antibody recognized only whole Bid molecule, but not the cleavage product. As shown in Figure 3A and B, knockdown of Bcl-2 increased loss of Δψm without effect on cleavage of Bid. These results suggest that Bcl-2 siRNA activates TRAIL-induced apoptosis on the level of Bid downstream.
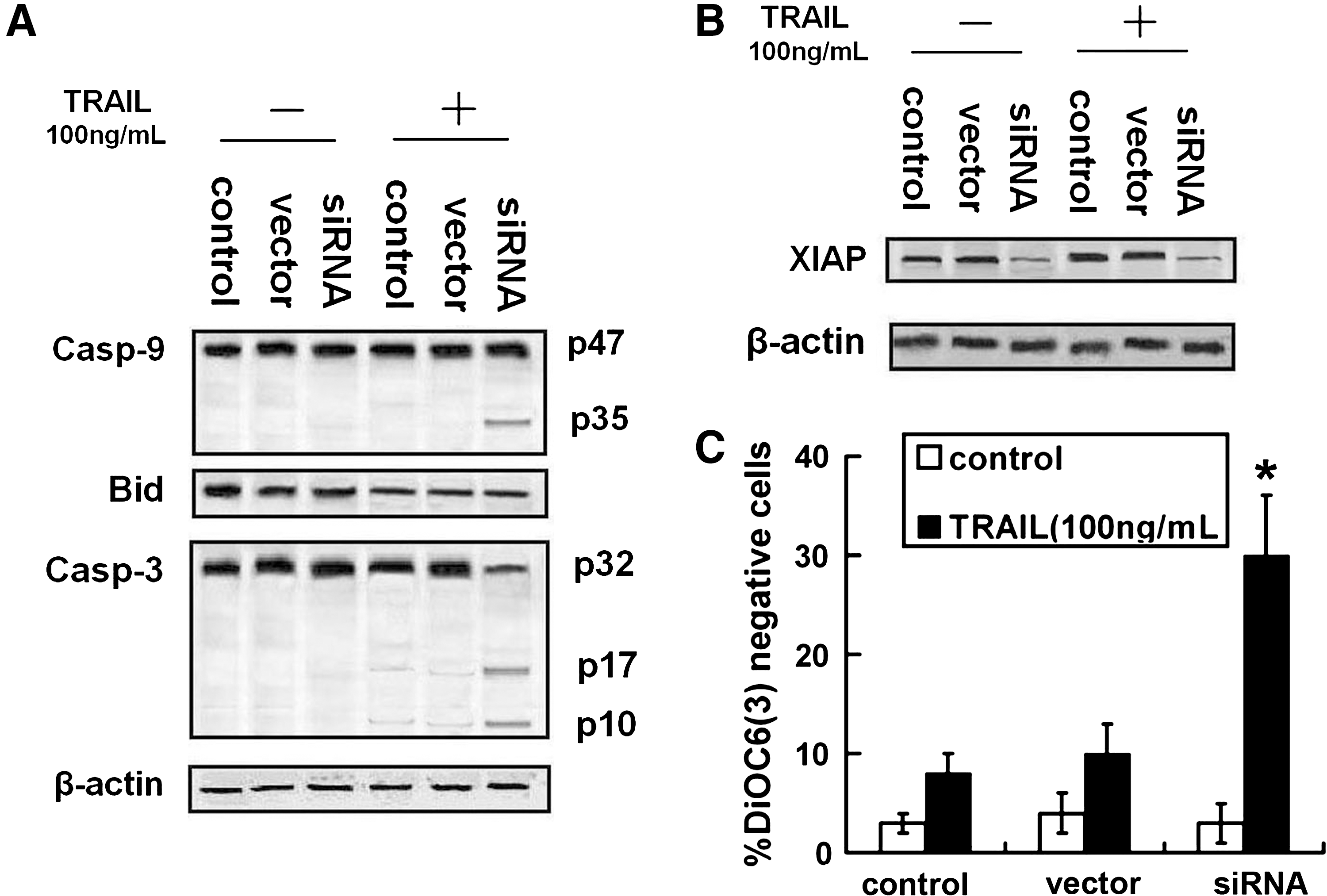
Effects of Bcl-2 downregulation on TRAIL-induced intrinsic pathway activation in CNE-2. After being treated with TRAIL (100 ng/mL) for 24 hours, CNE-2 transfected with Bcl-2-siRNA or not was detected.
XIAP exerts their antiapoptotic function by inhibiting proteolytically processed caspases including caspase-3 and caspase-9. 21 Therefore, XIAP was considered to be a regulator of intrinsic pathway. In this study, protein expression of XIAP was decreased by Bcl-2 siRNA (Fig. 3C).
Effects of PI3-K inhibitors on TRAIL-induced apoptosis in NPC cells
As shown in Figure 4A, treatment with LY-294002 (1 μM) or Wortmannin (1 μM) for 8 hours reversed the high constitutive activity of Akt obviously without effects on protein expressions of death receptors and Bcl-2. CNE-2 cells were pretreated with LY-294002 or Wortmannin for 8 hours, followed by treatment with 100 ng/mL TRAIL for 24 hours, and then apoptotic rate was detected by flow cytometry analysis. As expected, pretreatment with LY-294002 or Wortmannin further enhanced TRAIL-induced apoptosis (Fig. 4B).
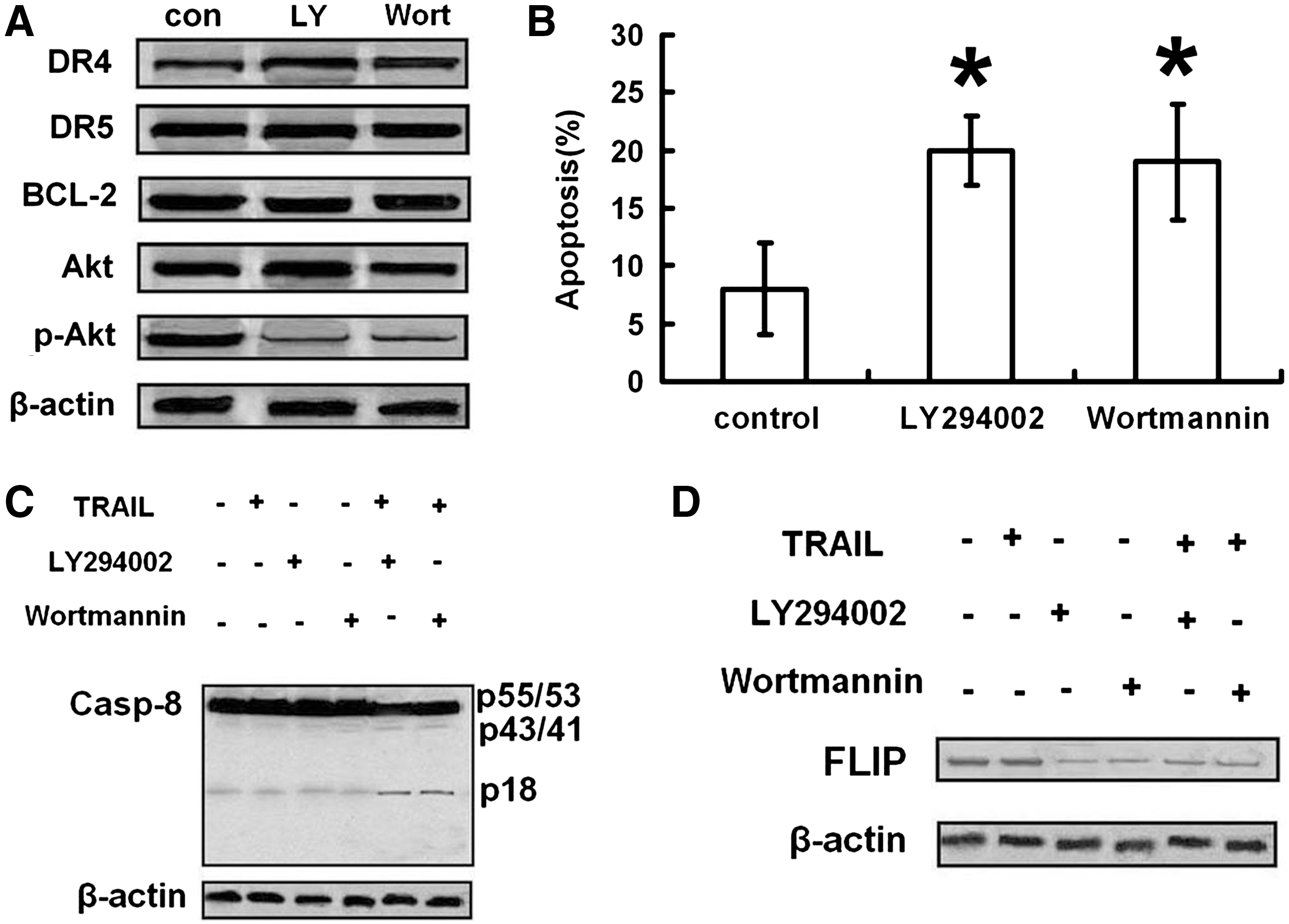
Phosphatidylinositol 3-kinase (PI3-K) inhibitors sensitized CNE-2 to TRAIL through activating the extrinsic pathway.
As shown in Figures 4C and D and 5A–C, PI3-K inhibitors increased cleavage of caspase-8, -9, and -3 and Bid, loss of Δψm, and decreased expressions of FLIP and XIAP.
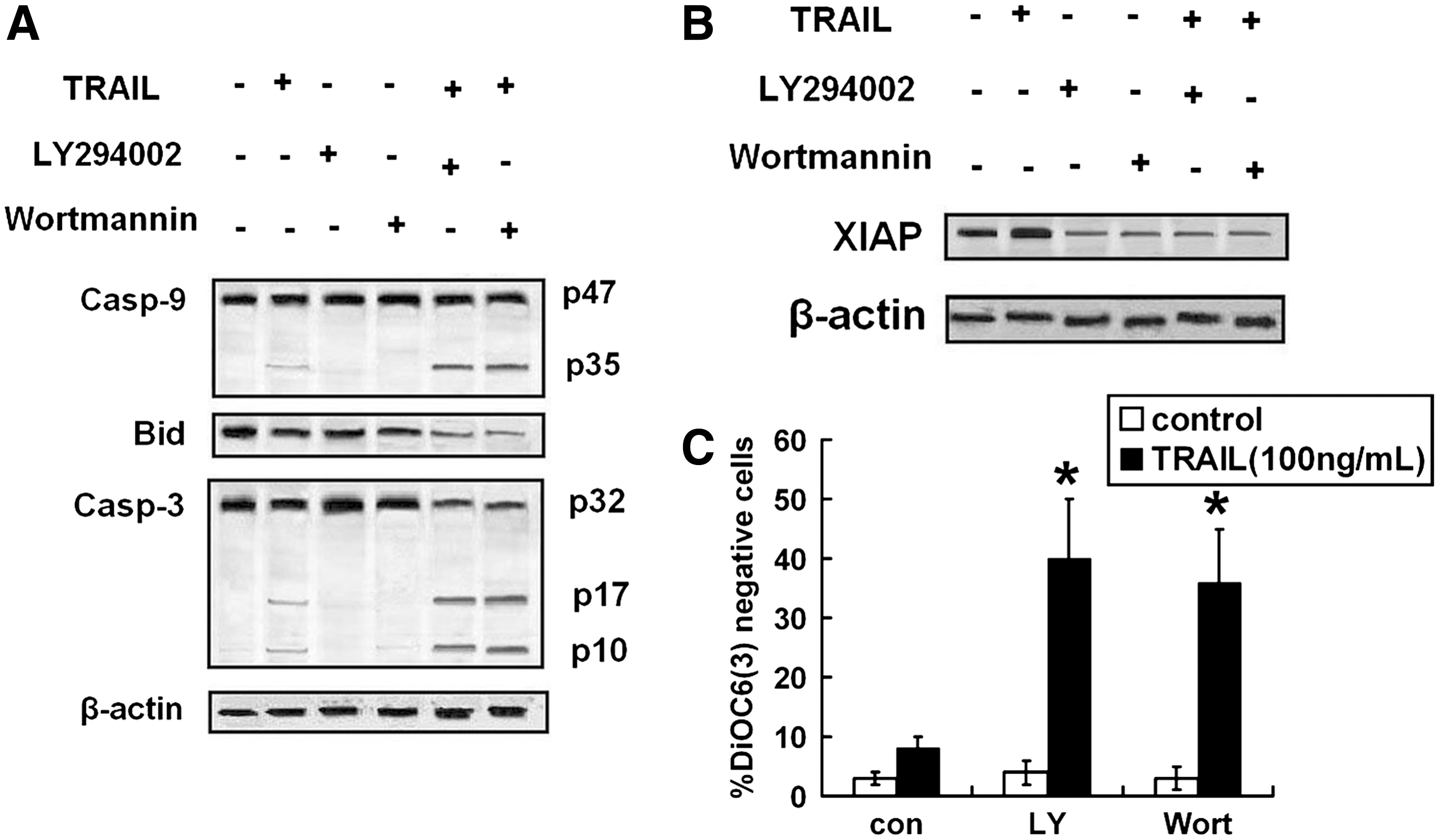
PI3-K inhibitors activated TRAIL-induced intrinsic pathway. CNE-2 cells were pretreated with LY-294002 (1 μM) or Wortmannin (1 μM) for 8 hours, followed by treatment with TRAIL (100 ng/mL) for 24 hours, and then
Effects of simultaneously targeting Bcl-2 and Akt on TRAIL-induced apoptosis in NPC cells
In this study, both Bcl-2 siRNA and PI3-K inhibitors sensitized CNE-2 to TRAIL by different mechanisms. These results lead to a speculation that simultaneously targeting Bcl-2 and Akt may enhance TRAIL-induced apoptosis synergistically. To test this hypothesis, the effects of combinative therapy on TRAIL-induced apoptosis were compared with single therapies. The results of flow cytometry analysis confirmed our hypothesis. Combinative therapy of Bcl-2 siRNA and LY294002 prevented TRAIL resistance more efficiently than single therapies (Fig. 6A). As shown in Figure 6B and C, combinative treatment enhanced cleavage of caspase-8 and Bid and decreased expression of FLIP to the same level of single treatment of LY294002. Further, combinative treatment enhanced cleavage of caspase-9 and -3 and loss of Δψm and decreased expression of XIAP more than single treatments (Fig. 6C, D).
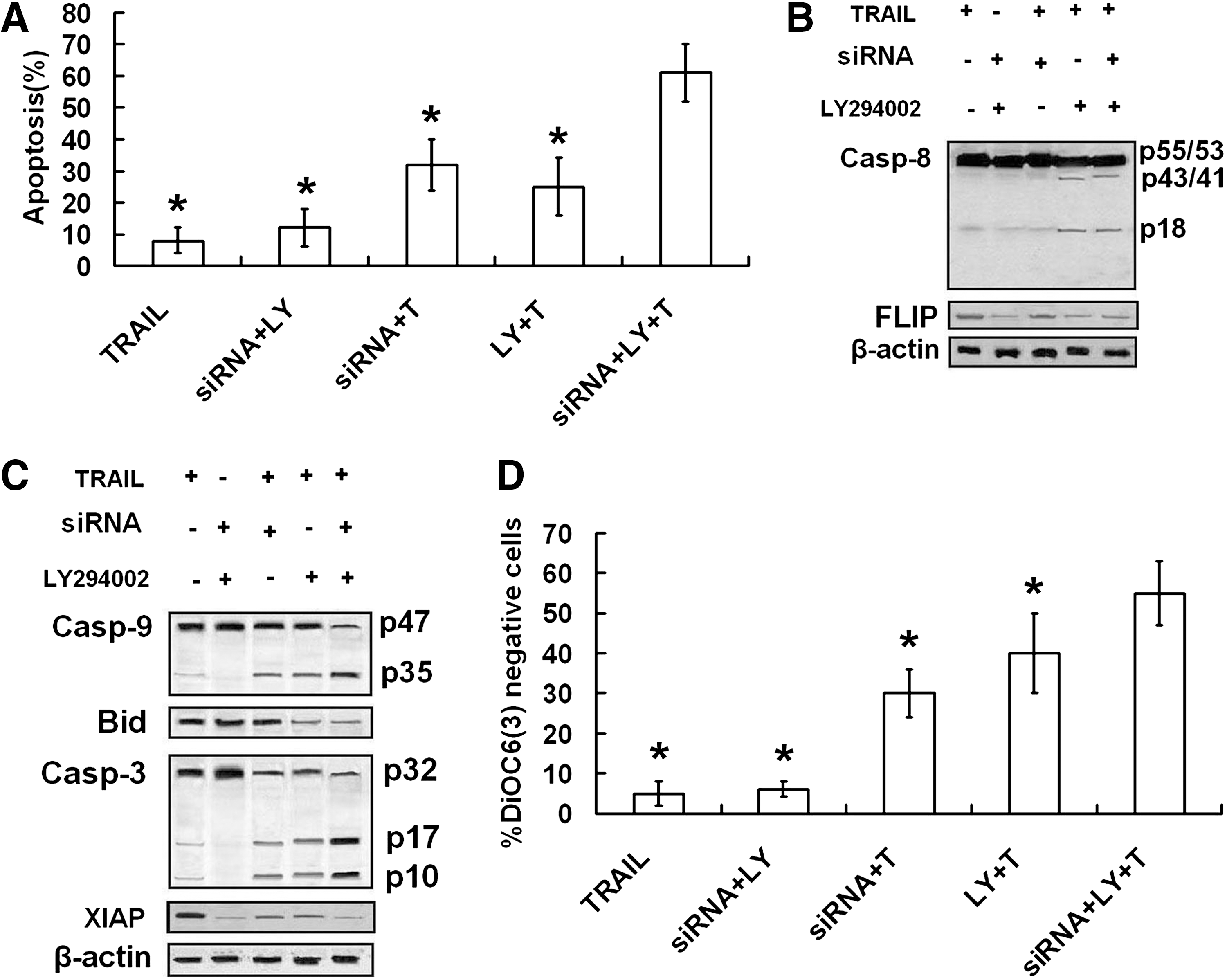
Effect of combinative treatment of Bcl-2-siRNA and LY294002 on TRAIL-induced apoptosis. CNE-2 transfected with Bcl-2-siRNA or not were pretreated with LY-294002 (1 μM) or Wortmannin (1 μM) for 8 hours, followed by treatment with TRAIL (100 ng/mL) for 24 hours, and then
Effects of combinative treatment of Bcl-2-siRNA and PI3-K inhibitors on TRAIL-induced apoptosis in normal cells
The extremely high interest in TRAIL is due to its selective antitumor and low toxicity toward normal cells. To understand the effects of combinative treatment on TRAIL-induced apoptosis in normal cells, the human benign fibroblast cell line MRC5 was selected to test the toxicity of the new therapy. As shown in Figure 7, combinative treatment of Bcl-2-siRNA and LY-294002 did not alter TRAIL-induced apoptosis in benign fibroblast cell.

MRC5, the human benign fibroblast cell line, transfected with Bcl-2-siRNA was pretreated with LY-294002 (1 μM) for 8 hours, followed by treatment with TRAIL (100 ng/mL) for 24 hours. The viabilities were detected by MTT assay.
Discussion
Despite aggressive therapies, resistance to current treatment protocols has been a major obstacle in clinical oncology. Most anticancer agents kill cancer cells by inducing apoptosis in cancer cells. 22 TRAIL is a promising candidate of cancer therapy via induction of apoptosis but its potential application has been limited by the development of drug resistance. Some studies have shown that TRAIL cannot induce apoptosis effectively in some NPC cell lines. 12 Therefore, defining the approaches of TRAIL-resistance prevention has become an important issue. In this study, it was observed for the first time that simultaneously targeting Bcl-2 and Akt pathway prevents resistance to the TRAIL-mediated apoptosis and cell death in NPC cells.
Bcl-2 overexpression in NPC correlates with progression and a poor prognosis, 23 and it is a factor in patient responses to chemotherapy or radiotherapy. 13,24 Prior studies have demonstrated a role of Bcl-2 on protecting cells from TRAIL-mediated apoptosis. 15 Inhibition of Bcl-2 has been recently tested as reactivators of the mitochondrial apoptotic pathway in many cancer cells. 25 In this study, Bcl-2 expression was decreased and TRAIL resistance was prevented. This augment of the TRAIL effect was associated with increases in activated caspase-9 and -3 consistent with apoptotic signaling. Bid, a subset of the Bcl-2 family, contains only the BH3 domain. BH3-only proteins interact with proapoptotic Bcl-2 family members to augment their activity. Once cleaved by caspase-8, Bid translocates to the mitochondria and activates Bax and Bak, leading to dissipation of the Δψm and cleavage of caspase-9 and -3. 4 In this study, knockdown of Bcl-2 increased loss of Δψm without effect on cleavage of Bid, suggesting that knockdown of Bcl-2 activated TRAIL-induced mitochondrion-dependent manner at the level of Bid downstream. Further, expression of XIAP was suppressed by Bcl-2 inhibition in the present study. In TRAIL-induced apoptosis, caspase activity is regulated by the inhibitor of apoptosis proteins (IAP). XIAP is the best characterized, which inhibits caspase-9 and -3 through binding to their intermediate and fully cleaved forms. 21 So it can be considered that inhibition of XIAP plays an important role on TRAIL-resistance prevention caused by knockdown of Bcl-2. In the present study, results show that knockdown of Bcl-2 reactivates TRAIL-mediated apoptosis through activation of intrinsic pathway.
A variety of reports have suggested the role of Akt on chemotherapeutic and radiotherapeutic resistance. 26,27 It has been demonstrated that Akt might inhibit a variety of apoptotic stimuli in multiple ways. 28 However, once expressed, active Akt is under tight regulation by PI3-K and other kinases of the signaling pathway that promote cell survival. As PI3-K targets Akt for survival, the activation of Akt was modulated by PI3-K inhibitors LY294002 and Wortmannin. In this study, increased cell apoptosis of NPC cells was observed when TRAIL combined with LY-294002 or Wortmannin. This suggests that activation of Akt in NPC cells is responsible for TRAIL resistance. Although ineffective alone, TRAIL in combination with LY-294002 enhanced cleavage of Bid and loss of Δψm and subsequently activated caspase-9 and -3 in CNE-2 cells. Prior studies showed that inhibition of Bid expression by Akt leads to resistance to TRAIL-induced apoptosis and LY294002 prevented TRAIL resistance through increasing of Bid cleavage. 29 FLIP has homology to caspase-8 and -10, but lacks protease activity. 19 It is, therefore, thought that FLIP recruitment to the DISC in the place of caspases blocks their activation to inhibit the extrinsic apoptotic pathway. Some overexpression studies have suggested that FLIP is an inhibitor of caspase-8 activation and TRAIL-induced apoptosis. 30 In this study, FLIP expression was inhibited by PI3-K inhibitors and resulted in cleavage of cleavage-8. These results suggest that targeting Akt pathway prevents TRAIL resistance through activation of both extrinsic and intrinsic pathways.
It was observed that Bcl-2-siRNA and PI3-K inhibitors sensitized NPC cells to TRAIL through affecting different modulators, respectively, providing a further rationale for the combinative therapy of them. When Bcl-2-siRNA and LY294002 were combined, the expected synergy in sensitizing NPC cells to TRAIL was observed. The synergistic effects were found in loss of Δψm and cleavage of caspase-9 and -3 but not in Bid cleavage and FLIP inhibition. In other words, the synergistic effects happened at the level of Bid downstream. Further, XIAP expression was decreased by combinative treatment more than single treatments. Altogether, the main functions of simultaneously targeting Bcl-2 and Akt were increasing of TRAIL-induced mitochondrial depolarization and suppression of XIAP synergistically.
Low toxicity toward normal cells leads to an extremely high interest in TRAIL. In the present experiment, neither single treatment nor combinative treatment (simultaneously targeting Bcl-2 and Akt) enhanced the toxicity of TRAIL in the benign fibroblast cells.
In summary, results from this study show that targeting Bcl-2 or Akt prevents TRAIL resistance of NPC cells through different mechanisms. Simultaneously targeting Bcl-2 and Akt prevents TRAIL resistance more efficiently through increasing of TRAIL-induced mitochondrial depolarization and suppression of XIAP synergistically. Thus, results from this study provide a new approach to treat NPC.
Footnotes
Disclosure Statement
There are no financial and personal relationships with other people or organizations that could inappropriately influence our work.