
Abstract
Introduction:
We aimed to investigate the effects of 12C6+ irradiation on the cell cycle and apoptosis as well as the associated mechanisms in the human lung cancer cell line H1299.
Methods:
After irradiation with different doses of 12C6+ for varying lengths of incubation, the changes in H1299 cells were assayed by flow cytometry and the microculture tetrazolium test. The expression of caspase-3 was detected by immunocytochemistry, western blot, and reverse transcription–polymerase chain reaction (RT-PCR).
Results:
The G2/M phase was blocked after treatment with 1 and 2 Gy at the 12-hour time point, and the most obvious block of G2/M occurred after treatment with 2 and 4 Gy at the 24-hour time point in a dose-dependent manner. The apoptosis rate increased with increasing radiation dose and reached a peak after the cells were irradiated with 2 Gy and incubated for 48 hours. In addition, the RT-PCR, western blot, and ICC results showed that irradiation with 12C6+ significantly increased the expression of caspase-3 compared with the control group (p<0.05).
Conclusions:
Irradiation of H1299 cells with 12C6+ induced apoptosis and significantly inhibited their growth through heavy ion irradiation-mediated activation of the caspase-3 pathway. Our results show that caspase-3 may play an important role in radiation-induced apoptosis through a p53-independent pathway.
Introduction
Intrinsic lung cancer has one of the highest mortality rates of malignant tumors, and the associated morbidity and mortality rates tend to increase each year. In addition, the five-year survival rate of patients with lung cancer is ∼20%. Radiotherapy has become the primary treatment option. Approximately 85% of lung cancer patients receive radiation therapy. Studies have shown that ionizing radiation can be used for therapeutic purposes by inducing apoptosis through a variety of signaling pathways. Caspase-3 is known to be the most important protease in the caspase family and is also a key effector enzyme in the multicomponent apoptosis pathway. In addition, X-rays can promote the apoptotic process in tumor cells. Our previous data showed that heavy ion irradiation can induce apoptosis in the SMMC7721 (p53 wt) cell line through a p53-dependent pathway. 1 However, the ability of heavy ion irradiation to induce apoptosis in p53-null cell lines through p53-independent pathways has not been reported to date. In this study, we chose the human lung cancer cell line H1299 (p53 null) to investigate the mechanisms of apoptosis induced by 12C 6 +-ray irradiation and to provide some basic data for heavy ion radiation therapy of lung cancer.
Materials and Methods
Cell culture
The H1299 cell line (p53-null human non-small cell lung cancer cell line) was obtained from the Shanghai Second Military Medical University (Shanghai, P.R. China) and used in this study. The cells were cultured in RPMI 1640 medium (Gibco, Co.) supplemented with 10% heat-inactivated fetal bovine serum (Minhai), 1% sodium pyruvate, and 2 mM glutamine (Sigma) at 37°C with 5% CO2 and 95% humidity. The cells were seeded at a density of 5×106 cells/mL in 35-mm glass Petri dishes (Nunc) 24 hours prior to irradiation with 12C6+.
Cell irradiation
Exponentially growing H1299 cells in Petri dishes were irradiated with a 50 Me V/u 12C6+ heavy ion beam supplied by the Heavy Ion Research Facility in Lanzhou. The linear energy transfer (LET) of the beam was 44.56 keV/μm. The intensity of the carbon ion beam was 2 Gy/min for cell exposure. After irradiation, the cells were incubated in normal culture medium for up to 72 hours. The dose applied to the cells ranged from 0 to 6 Gy, and nonirradiated H1299 cells were used as a control.
Hoechst 33258 staining
After irradiation with different doses of heavy ions, the cells were digested with trypsinase. The cells were washed with phosphate-buffered saline (PBS; pH 7.4), fixed with 4% paraformaldehyde for 10 minutes, washed again with PBS, and then stained with 1 mM Hoechst 33258 (Sigma) for 30 minutes. Morphological changes in the apoptotic cells were observed under a fluorescence microscope (Olympus IX50) with an excitation wavelength of 330–380 nm.
Cell cycle analysis
H1299 cells in the logarithmic phase were irradiated randomly with 12C6+ at doses of 0, 1, 2, 4, and 6 Gy. The cell suspensions were then digested with 0.25% trypsinase and collected after culturing for 12, 24, 48, and 72 hours. The cells were washed two times with prechilled (4°C) PBS and then fixed with 70% ethanol overnight at 4°C. The next day, the cells were washed twice with PBS and then stained with propidium iodide for 20 minutes. Finally, flow cytometry (FACScan; Becton Dickinson) was performed to determine the cell cycle phase and apoptosis rate using Cell Quest ModFit software (Verity Software House) for analysis.
Microculture tetrazolium test
After irradiation with different doses of heavy ions, logarithmically growing cells were digested with trypsinase. The cells were counted and the concentration was adjusted to 5×104 cells/mL. The cells were seeded in a 96-well plate at 200 μL/well (∼1×104 cells/well) in triplicate for each dose level. The cells were cultured at 37°C in a 5% CO2 incubator for 12, 24, 48, and 72 hours, respectively, and then 20 μL of freshly prepared microculture tetrazolium test (MTT) (5 mg/mL) was added to each well. The cells were then incubated at 37°C for an additional 4 hours. At the end of the incubation, the supernatant was removed and 150 μL dimethyl sulfoxide was added to each well. The optical density (OD) value of each well was measured with an enzyme-linked immunosorbent assay (ELISA; Bio-Tek Powerwave X) instrument at a wavelength of 492 nm. Each measurement was repeated at least 3 times. Cell viability (%) was calculated as the average OD value of each experimental group divided by the average OD value of the control group ×100%.
Enzyme activity of caspase-3
To determine the activity of caspase-3, a caspase-3 fluorescence assay kit (Beyotime Biochem) was used according to the manufacturer's instructions. The cells were collected and lysed on ice for 30 minutes and the lysate was then centrifuged at 10,000 rpm for 10 minutes. The supernatant was then collected, placed on ice, and 5 mL of caspase-3 substrate (AC-DEVD-pNA) was added to a final concentration of 50 μL/L. The samples were incubated at 37°C for 2 hours and the absorbance was subsequently measured at 405 nm with an ELISA instrument.
Reverse transcription–polymerase chain reaction of caspase-3 RNA
To determine caspase-3 RNA expression levels, the cells were divided into the following groups: 0, 2, and 4 Gy. After the cells were cultured for 24 hours, RNA was extracted from each group using Trizol reagent (Invitrogen), and then cDNA was synthesized by reverse transcription–polymerase chain reaction (RT-PCR). The RT reaction contained the following components: 1 μg RNA, 0.5 μL random primer, 1 μL deoxyribonucleoside triphosphate (dNTP), 0.5 μL reverse transcriptase, 0.25 μL RNase, 1 μL 10× reverse transcriptase buffer, and the final volume of the reaction was increased to 10 μL with RNase-inactivated H2O. The PCR contained the following components: 10 μL 5× PCR buffer, 0.25 μL Taq, 0.5 μL each of upstream- and downstream-specific primers for caspase-3 as well as β-actin internal control, respectively, and sterile H2O to a final volume of 50 μL. The primers were synthesized at the Shanghai Shenggong Company. The caspase-3 primers were as follows: sense, 5′-CGT GTA TTG TGT CCA TGC TCA C-3′ and antisense, 5′-CCA TCA TTG ACA GTT ACT TGC TCC-3′. The β-actin internal control primer sequences were as follows: sense, 5′-ACA TCG TGC GTG ACT TCA-3′ and antisense, 5′-CTC GTC ATA CTC CTG CTG-3′. The amplification conditions were as follows: 94°C for 5 minutes, followed by 30 cycles of 94°C for 30 seconds, 50°C for 30 seconds, and 72°C for 45 seconds. This was followed by a final cycle of 72°C for 10 minutes. A total of 10 μL of the PCR products was resolved by agarose gel electrophoresis and the UVP EC3 Imaging System (UVP; LLC) was used to take and scan photographs.
Western blot analysis of caspase-3 protein expression
H1299 cells were irradiated at different doses and then cultured for 24 or 48 hours. Total protein was extracted using a routine method. The Lowry protein assay was used to calculate the protein concentration in the samples. Cell lysate was normalized and then mixed with 5× sodium dodecyl sulfate (SDS) loading buffer. The samples were then incubated at 10°C for 5 minutes and allowed to cool to room temperature naturally. The samples (50 μg) and protein marker were then resolved by SDS-polyacrylamide gel electrophoresis. After electrophoresis, the gel was rinsed with deionized water and then transferred to a polyvinylidene difluoride membrane (Millipore) using the semidry electric transfer method. After the transfer was complete, the membrane was blocked in 5% skim milk powder in TBS-T solution (TBS/0.05% Tween-20) at room temperature for 1 hour. To visualize the protein on the membrane, the blots were probed with the rabbit anti-human caspase-3 antibody (dilution 1:500; BioLegend) and incubated at 4°C overnight. The next day, the membrane was washed twice with TBS-T solution and then incubated with a secondary antibody (dilution 1:4000; Southern Biotech) at room temperature for 1 hour. Immunoblots were developed using an enhance chemiluminescene western blotting detection kit (ZhongShan, Co.). The immunoblots were subsequently reprobed with a 1:5000 dilution of an anti-β-actin antibody (Lab Vision) and developed as previously described. For densitometric analysis, scanned photographs were quantified using the AlphaEasy FC software package (Alpha Innotech).
Immunocytochemistry
H1299 cells were grown on glass coverslips; irradiated with 0, 2, or 4 Gy; and then cultured for 24 hours. The cells were fixed with 4% paraformaldehyde for 30 minutes at room temperature, and then treated with cold acetone (−20°C) for 5 minutes. The coverslips were washed with PBS and then incubated with anti-caspase-3 antibody (Beyotime Biochem) for 2 hours at room temperature. They were then washed again with PBS and incubated with an anti-mouse secondary antibody (dilution 1:500; Southern Biotech) for 50 minutes at room temperature. The coverslips were mounted on slides and the images were captured with a digital camera (Nikon-DXM1200).
Statistical analyses
Data were calculated as mean±standard error of the means. Potential differences among the experimental groups (irradiated vs. nonirradiated rats) were assessed by using the Student's unpaired t-test for 2 groups. Values were considered significantly different when p≤0.05.
Results
12C6+-induced apoptosis of H1299 cells
12C6+ irradiation of H1299 cells induced apoptosis, as shown by staining with Hoechst 33258. Control cells showed normal nuclear morphology. After irradiation with 4 Gy and subsequent incubation of the cells for 24 hours, membrane bubbling and apoptotic body formation were observed, which are characteristics of apoptosis (Fig. 1).
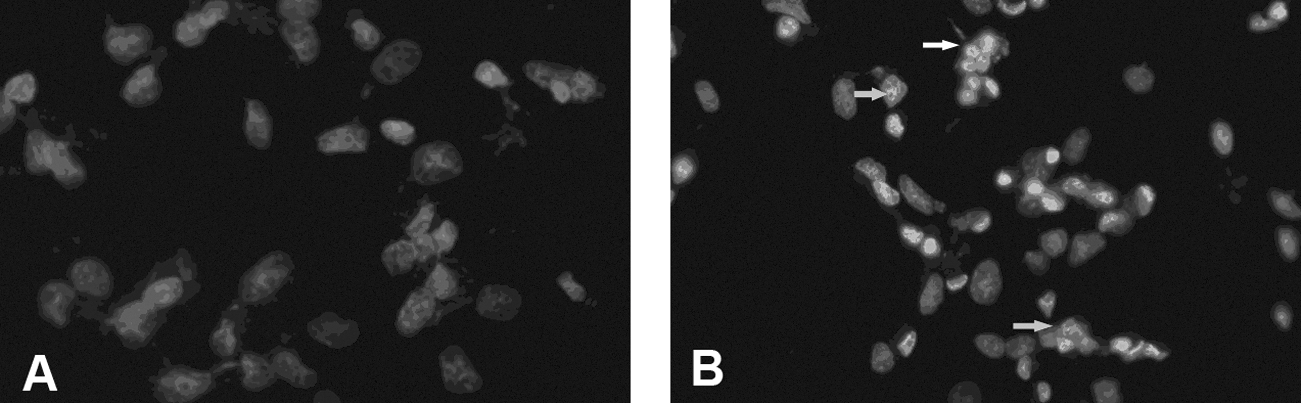
A photograph of H1299 cells after irradiation with 4 Gy of 12C6+ and subsequent incubation for 24 hours.
The effect of 12C6+ on proliferation of H1299 cell in a time- and dose-dependent manner
The proliferation of H1299 cells was assayed by MTT. When cells were treated with 2 Gy of 12C6+ and then cultured for 24 hours, there was a statistically significant reduction in proliferation compared with control cells (OD 0.820±0.023 vs. 0.993±0.002, respectively, p<0.05). The survival rates of cells cultured for 24, 48, and 72 hours were 82.58%, 71.25%, and 71.80%, respectively, compared with the control group (p<0.05). These results showed that treatment of H1299 cells with 2 Gy of 12C6+ irradiation had a time- and dose-dependent effect on proliferation. Similarly, as the dose of 12C6+ was increased, the survival rate of the cells decreased (Fig. 2).

Survival rate of the H1299 cells after irradiation with 12C6+ using different doses and for varying lengths of subsequent incubation time.
The effect of 12C6+ on cell cycle in H1299 cells
The number of H1299 cells in G2/M phase significantly increased and the number of cells in G1 phase decreased after irradiation with 12C6+ at different doses followed by a 24-hour culture period (Fig. 3) (p<0.01). When the radiation dose of 12C6+ was 6 Gy, the number of cells in G1 phase significantly increased, which indicated that the cells were dead. The cell death maybe attributed to cytotoxicity from irradiation at a high dose of 12C6+. The retardation in the G2/M phase induced by 12C6+ also occurred in a time-dependent manner. Irradiation of the cells with 4 Gy caused an increase in cell retardation in the G2/M phase (p<0.05), and the number of cells in S phase concomitantly decreased (p<0.05). These results indicated that irradiation inhibits DNA replication, which thereby reduced the cell proliferation.
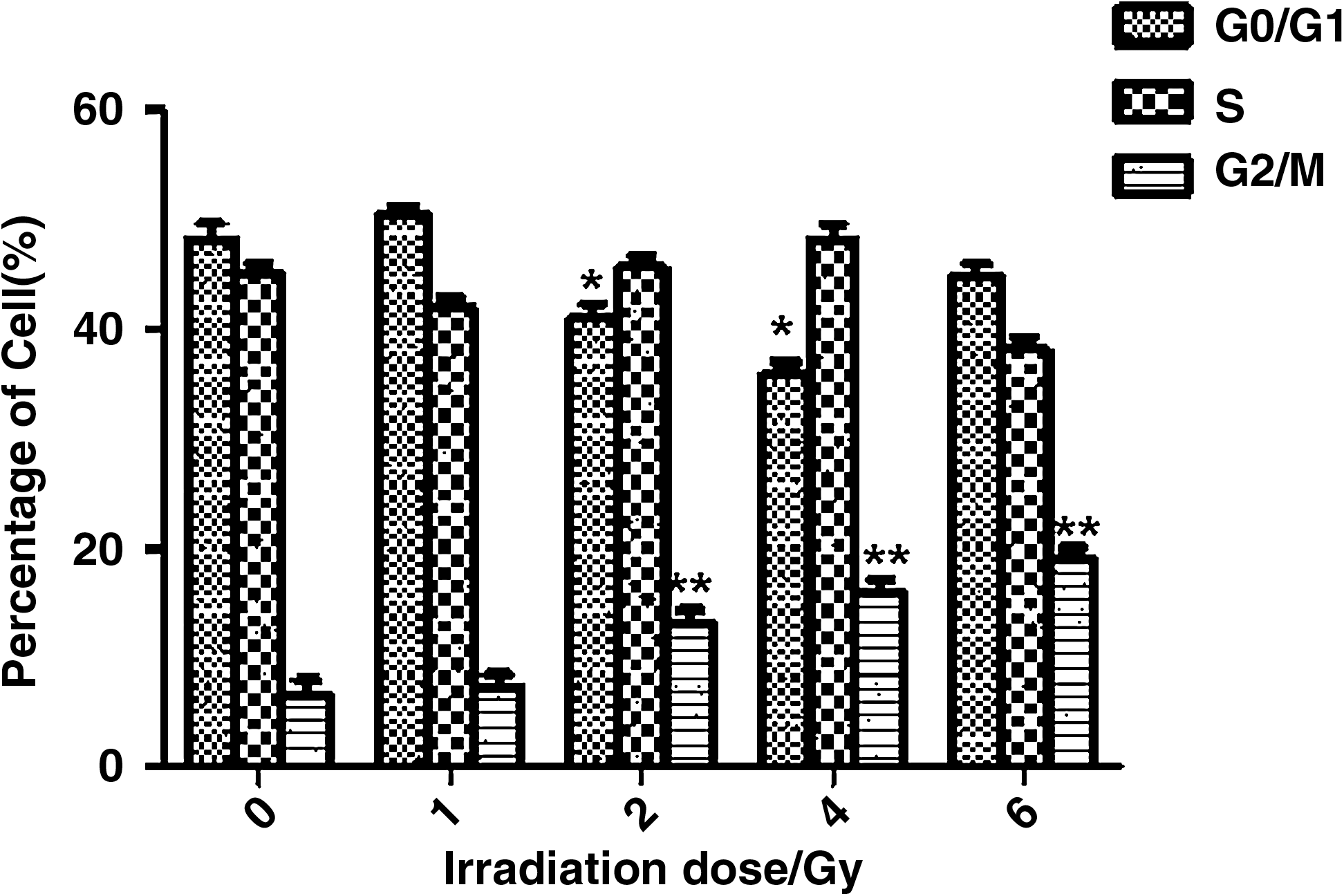
Cell cycle analysis of H1299 cells after irradiation with different doses of 12C6+ and subsequent incubation for 24 hours. Statistically significant differences compared with control groups are indicated by asterisks: *p<0.05 and **p<0.01.
The effect of 12C6+ on caspase-3 activity in H1299 cells
Caspase-3 activity significantly increased (p<0.05) after cells were treated with 12C6+ and cultured for 12, 24, and 48 hours in a dose-dependent manner (Fig. 4).
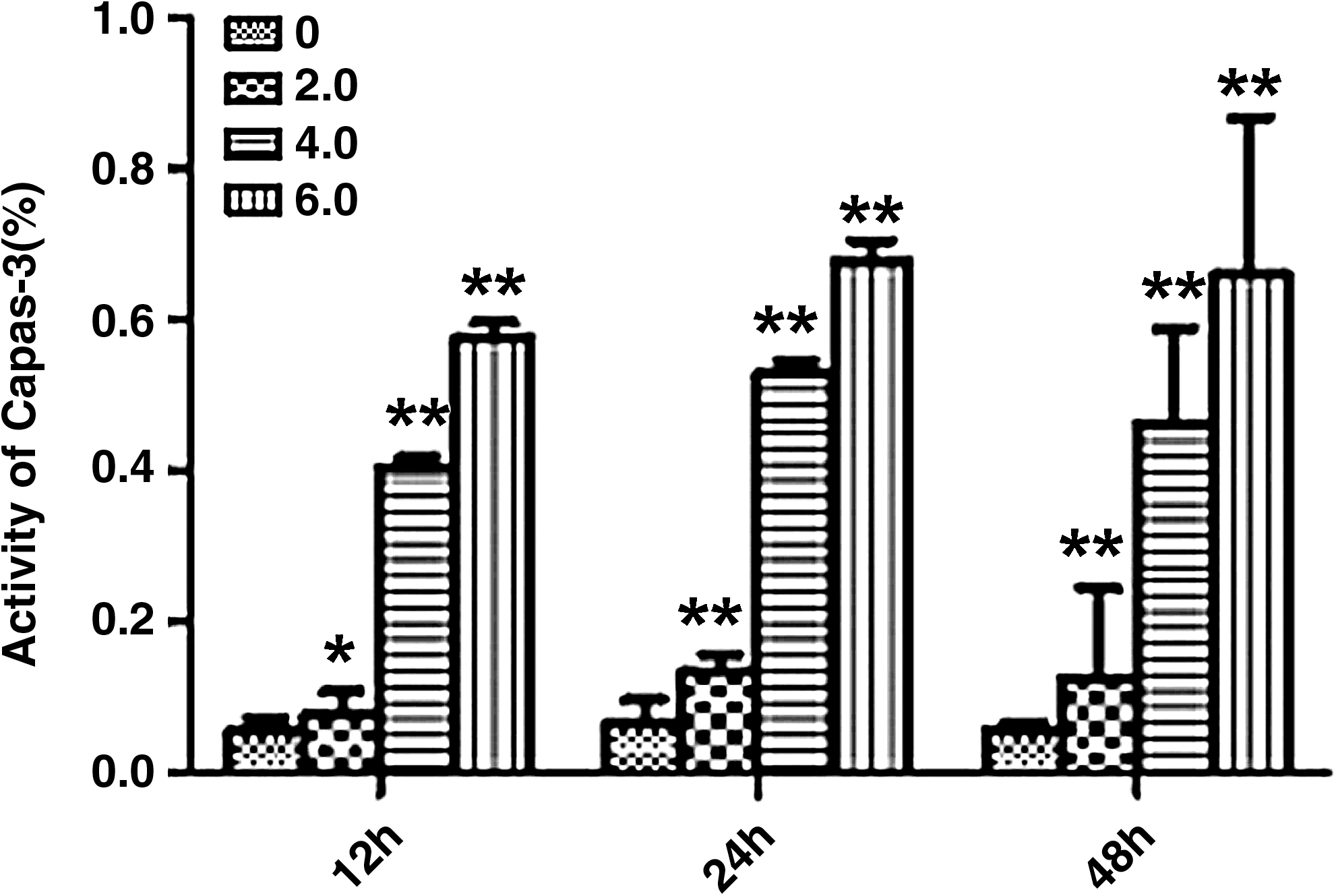
Caspase-3 activity at different irradiation doses and time points. Statistically significant differences compared with control groups are indicated by asterisks: *p<0.05 and **p<0.01.
Caspase-3 mRNA expression in H1299 cells by RT-PCR
After H1299 cells were irradiated with 12C6+ heavy ions, the expression of caspase-3 mRNA increased in a dose-dependent manner, which was shown by the intensity of the mRNA band that corresponded to caspase-3 (Fig. 5). The control H1299 cells only exhibited a single band, which corresponded to the β-actin internal control. These results showed that caspase-3 mRNA was not expressed or detected in the H1299 control group, whereas it was induced and expressed in a dose-dependent manner by 12C6+ in the test groups (Fig. 5).
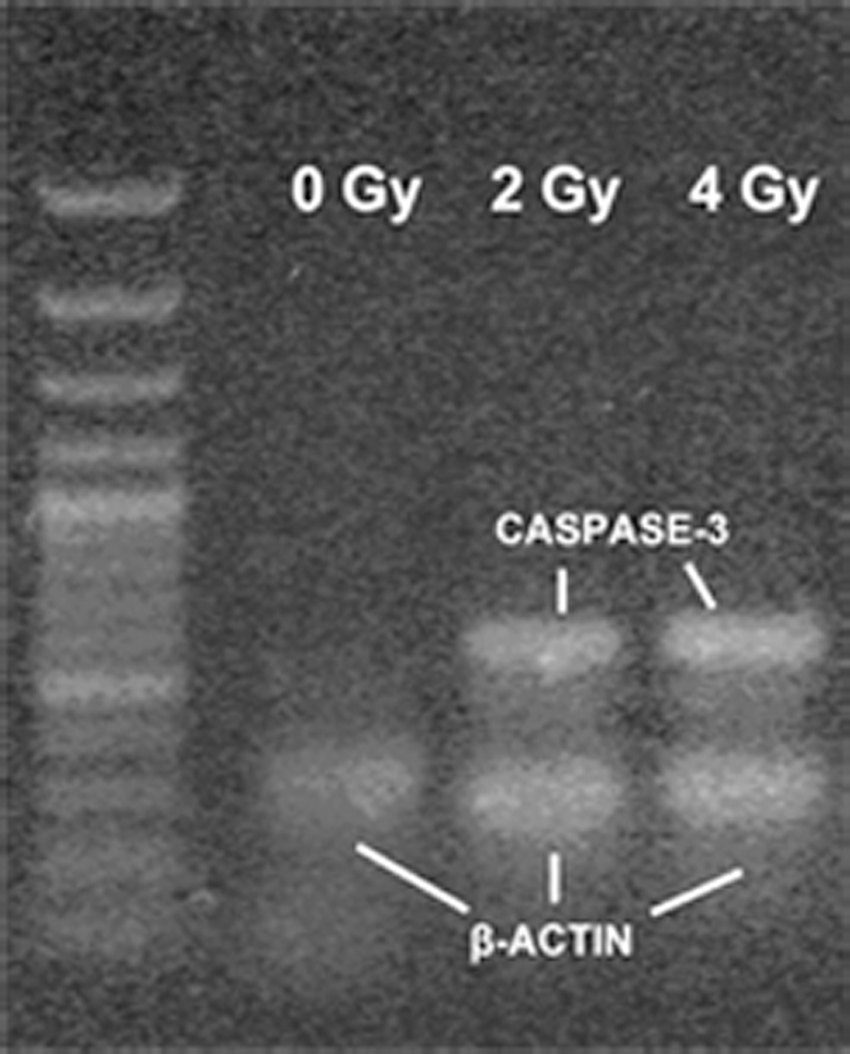
Caspase-3 mRNA expression in H1299 cells after exposure to 12C6+ and subsequent incubation for 24 hours. A caspase-3 mRNA band was not detected in the H1299 control group, while the test groups irradiated with heavy ions had 2 bands. The band corresponding to caspase-3 escalated as the radiation dose increased.
Detection of caspase-3 protein by western blot
After H1299 cells were irradiated with 12C6+ heavy ions, the expression of caspase-3 protein increased in a dose-dependent manner, which was shown on western blot by the intensity of the protein band that corresponded to caspase-3 protein (Fig. 6).
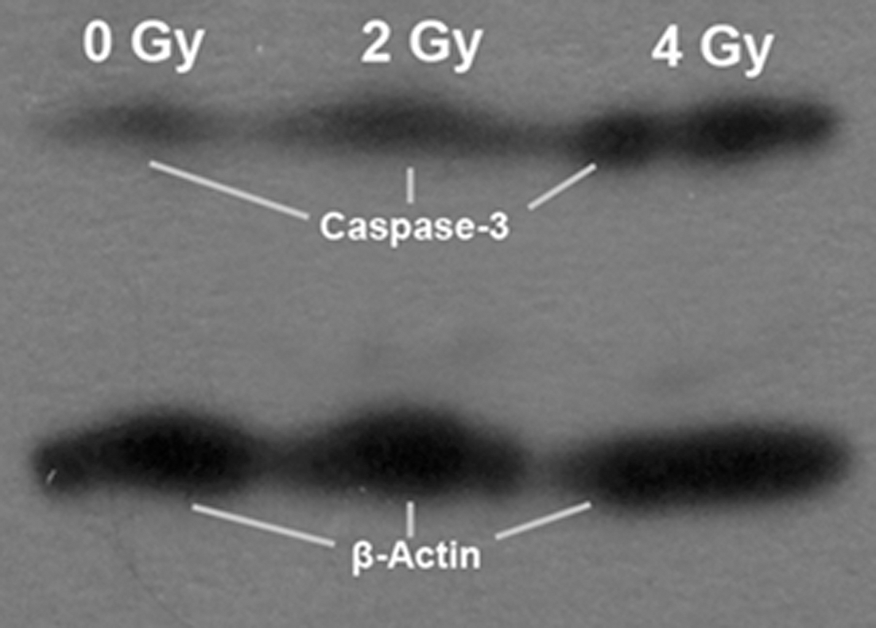
Western blot analysis of caspase-3 protein in H1299 cells after irradiation and subsequent culture.
Detection of caspase-3 protein in H1299 cells by ICC
Immunocytochemistry (ICC) experiments showed that caspase-3 was located in the cell nucleus and cytoplasm, as shown by brown granules, and accompanied by abnormal cell morphology (Fig. 7). The number of caspase-3–positive cells significantly increased when the dose of 12C6+ radiation was increased compared with the control group (p<0.01).

Immunocytochemistry analysis of caspase-3 protein in H1299 cells.
Discussion
Radiation therapy is an important tool for the clinical treatment of cancer. Radiation biology studies have shown that ionizing radiation can cause serious DNA damage in cells. Cells use a variety of mechanisms to maintain genome integrity. In general, a cell uses complex regulatory mechanisms to respond to DNA damage, including cellular repair, cell cycle retardation, apoptosis, and necrosis. Apoptosis is induced by ionizing radiation and represents the predominant mechanism for cell death. 2 Apoptosis is an important mechanism that organisms used to maintain stability and balance. Apoptosis is an initiative suicide process of the cell encoded by gene regulation and provides an ideal approach for preventing the abnormal proliferation of cells. 3 DNA damage or mutations can block the damaged cell at the corresponding checkpoint. 4 The ultimate goal of these safeguards is to ensure the genetic stability of the genome. It is generally believed that high doses of ionizing radiation cause cell death, while low doses induce apoptosis.
It is well-known that the p53 gene is one of the most important genes involved in cell cycle regulation and plays an important role in apoptosis. The treatment of cancer with X-ray radiation can trigger p53 that can quickly stimulate and induce the process of apoptosis. The delay of cell cycle progression caused by the activation of radiation-induced p53 mainly occurs in the G1-S phase, which provides the cells with time for damaged DNA to be repaired before replication and division. However, once the repair fails, p53 induces the apoptosis pathway. In addition, it is well-known that caspase-3 is a key molecule in the apoptotic signaling pathway. Activation of caspase-3 plays a central role in apoptosis signal transduction. Moreover, caspase-3 is the key protein kinase that is activated during the process of apoptosis.
Caspase-3 is a signaling factor in the caspase-dependent pathway. Gashegu et al. 5 reported that caspase-3 is a key protease in ionizing radiation-induced apoptosis, and in radiation-induced cell death, apoptosis, and necrosis. Michelin et al. 6 reported that primary cerebral cortex nerve cells of mice irradiated with 0–4 Gy of X-rays exhibited a dose-dependent increase in caspase-3 expression. Various factors, such as ionizing radiation, can activate caspase-3. Caspase-8 and caspase-9 the initiators of the caspase pathway can lead to the activation of caspase-3, which is the key enzyme that executes the signal of the pathway. In addition, caspase-3 is necessary not only in terms of the mitochondrial pathway, but also in death receptor pathway–induced apoptosis.
In this study, our results showed that the proliferation of H1299 (p53 null) cells was clearly inhibited after heavy ion irradiation in a dose-dependent manner. As the dose of 12C6+ radiation increased, the cells arrested in the G2/M phase in a dose-dependent manner (p<0.01). Ning and Knox 7 also reported that the effect of radiation-induced apoptosis can be promoted by extending the G2/M block. When the dose of 12C6+ was 6 Gy, the number of cells arrested in G1 significantly increased, indicating that the cells were dead. The observed cell death maybe attributed to cytotoxicity from the high dose of 12C6+ irradiation. When H1299 cells were irradiated with 12C6+ and then cultured for 24 hours, caspase-3 was clearly detected by western blot and ICC. Moreover, the protein band that corresponded to caspase-3 became gradually stronger in a dose-dependent manner, which showed that irradiation with 12C6+ can activate caspase-3 and causes an upregulation of expression in a dose-dependent manner. Our assessment also showed an increase in caspase-3 enzymatic activity, which was in agreement with the increase in caspase-3 mRNA expression. The differences between the mRNA expression, protein level, and caspase-3 activity detected by RT-PCR, western blot, and ICC, respectively, were due to the occurrence of apoptosis. Therefore, caspase-3 activation may be one important mechanism of 12C6+-induced apoptosis in H1299 cells. Irradiation induces a reduction in the membrane potential of mitochondria, the release of cytochrome C, and activation of caspase-9. Activated caspase-9 then activates the downstream effector protease, caspase-3. Activated caspase-3 hydrolyzes proteins and leads to cell disintegration, apoptotic body formation, and apoptosis promotion. 8 In addition, it has been shown that apoptotic pathways triggered by high LET radiation in H1299 do not require p53. 9
In summary, heavy ion irradiation may activate the expression of caspase-3 in H1299 cells as a mechanism for inducing apoptosis, and the expression level of caspase-3 is proportional to the radiosensitivity of the cells. Caspase-3 is also involved in the regulation of the cell cycle as well as the induction of apoptosis as a mechanism of the antitumor effects. Irradiation with 12C6+ can induce apoptosis in H1299 cells through a p53-independent pathway. By regulating the expression of the caspase-3 gene, the radiosensitivity of tumor cells may change, which would be expected to increase the effectiveness of tumor radiation therapy and provide new ideas and methods for therapeutic intervention.
Conclusions
Irradiation with 12C6+ can induce apoptosis in the H1299 human lung cancer cell line. The mechanisms were shown to be associated with the activation of caspase-3. Caspase-3 may play an important role in a p53-independent pathway in these cells. In addition, the results in this study provide basic radiobiological data for human lung cancer therapy using heavy ions.
Footnotes
Disclosure Statement
No completing financial interests exist.