
Abstract
Inhibitor of growth 4 (ING4) is a novel member of ING tumor suppressor family and has apparent tumor-suppressive effect. Interleukin-24 (IL-24) as a unique cytokine-tumor suppressor displays ubiquitous antitumor property and tumor-specific killing activity. Multigene-based combination therapy may be an effective practice in cancer gene therapy. The therapeutic potential of a conjunction of ING4 and IL-24 for cancers is still elusive. This study evaluated the combined effect on SMMC-7721 and HepG2 human hepatocarcinoma cells by adenovirus-mediated ING4 and IL-24 coexpression (Ad-ING4-IL-24) and also elucidated its underlying molecular mechanism. It was demonstrated that Ad-ING4-IL-24 induced synergistic growth inhibition, apoptosis, invasion suppression, as well as an enhanced effect on upregulation of P21, P27, Fas, FasL, FADD, Bad, Bax, Bak, cleaved Bid, cleaved Caspase-8, -9, and -3, and cleaved PARP, downregulation of Bcl-2, Bcl-XL, matrix metalloproteinase (MMP)-2, 9, vascular endothelial growth factor (VEGF), IL-8, CD34, and microvessel density, and cytochrome c release from mitochondria into cytosol in in vitro SMMC-7721 and HepG2 hepatocarcinoma cells and/or in vivo SMMC-7721 hepatocarcinoma subcutaneous xenografted tumors in athymic nude mice. The in vitro and in vivo synergistic antitumor activity elicited by Ad-ING4-IL-24 was closely associated with the cooperative activation of extrinsic and intrinsic apoptotic pathways and reduced proangiogenic factors' production of VEGF and IL-8, leading to synergistic inhibition of tumor angiogenesis. Thus, results indicate that cancer gene therapy combining two or more tumor suppressors such as ING4 and IL-24 may constitute a novel and effective therapeutic strategy for hepatocarcinoma and other cancers.
Introduction
Inhibitor of growth (ING) is a type II tumor suppressor family implicated in carcinogenesis, apoptosis, cell cycle control, DNA repair, and senescence, including 5 known members (ING1–ING5). Each member of ING family contains a functionally conserved plant homeodomain-finger motif in the COOH-terminal region involved in chromatin remodeling-mediated transcriptional modulation via interaction with histone acetyltransferase and histone deacetylase complexes. 1 –3 ING4, a novel member of ING family, is frequently deleted or downregulated in breast carcinoma, 4 head and neck carcinoma, 5 glioblastoma, 6 hepatocellular carcinoma (HCC), 7 melanoma, 8 and gastric carcinoma, 9 which is closely correlated with tumor progression and prognosis. ING4 has recently attracted much attention as a strong candidate tumor suppressor. It has been reported that ING4 can significantly induce growth inhibition and apoptosis in a variety of tumor cells 6,10 –14 and enhance chemosensitivity to DNA-damage agents such as doxorubicin and etoposide in HepG2 hepatocarcinoma cells. 11 ING4 can repress the loss of contact inhibition elicited by MYCN or MYC family oncogenes. 4 ING4 can also suppress brain tumor angiogenesis via transcriptional repression of nuclear factor kappaB (NF-κB)-downstream genes including interleukin (IL)-6, IL-8, and prostaglandin-endoperoxide synthase 2 (Cox-2). 6 In addition, ING4 can suppress hypoxia inducible factor-1α (HIF-1a) activity and decrease expression of IL-8 and osteopontin in myeloma cells and consequently inhibit tumor angiogenesis. 15 Further, ING4 can inhibit tumor cell spreading, migration, and invasion by colocalizing and interacting with liprin α1 at lamellipodia 16 and downregulating expression and activity of matrix metalloproteinase-2 (MMP-2) and MMP-9. 8,12 These findings revealed that ING4 as a potent tumor suppressor negatively modulates tumor growth via multiple pathways.
Melanoma differentiation-associated gene-7/IL-24, originally identified in human melanoma cells treated with interferon-β (IFN-β) and mezerein by subtraction hybridization, 17 is a unique cytokine-tumor suppressor belonging to IL-10 family. 18 Extensive studies have shown that IL-24 displays ubiquitous antitumor property and tumor-specific killing activity. Ectopic expression of IL-24 can remarkably induce growth suppression and apoptosis in a broad spectrum of cancer cells but not in normal cells. 19 IL-24 can also inhibit tumor angiogenesis through directly suppressing vascular endothelial cell differentiation and migration 20,21 and indirectly downregulating production of proangiogenic factors such as vascular endothelial growth factor (VEGF), basic fibroblast growth factor, transforming growth factor-β, and IL-8. 20,22,23 Ramesh et al. further demonstrated that IL-24 can exert the antiangiogenic activity via interacting with IL-22R1/IL-20R2 heterodimeric receptor on vascular endothelial cells. 21 IL-24 as a pro-Th1 cytokine can also exhibit a potent immunostimulatory activity via inducing robust production of IL-6, tumor necrosis factor-α, and IFN-γ from peripheral blood mononuclear cells, leading to an enhanced antitumor immunity. 24 In addition, IL-24 can repress tumor cell invasion and migration by downregulation of phosphatidylinositol 3-kinase (PI3K), focal adhesion kinase, and MMP-2 and -9. 25 It has also been reported that IL-24 can sensitize tumor cells to radiation-, chemotherapy-, and monoclonal antibody-induced antitumor effect. 19 Further, IL-24 can exhibit a profound bystander antitumor activity and augment its antitumor therapeutic potential. 19 More recently, IL-24 can induce a toxic form of autophagy in tumor cells through protein kinase R-like endoplasmic reticulum kinase activation. 26,27 Thus, IL-24 is a promising tumor suppressor and now is being hailed as a “magic bullet” for cancer.
HCC is the fifth most common solid tumors worldwide, representing the third highest cause of cancer-related mortality. 28 Treatments for HCC depend on both the stage of tumor and liver function, including surgical resection, liver transplantation, locoregional therapy, and chemotherapy. 29 Unfortunately, the therapeutic potential and long-term prognosis of conventional treatments for HCC is still far from satisfactory. Hence, the search for novel therapeutic modalities for HCC is urgently needed. Gene therapy represents a promising therapeutic modality for cancers. Replacement and modification of tumor suppressor gene is one of the most gene therapy-based anticancer strategies. However, the carcinogenic process is multistep in terms of its etiology and multifactor contributing to its development, and tumor cells often undergo multiple genetic abnormalities, which limit efficacy of a single gene-mediated cancer therapy and becomes an obstacle of cancer gene therapy because of the difficulty to find a pivotal gene closely associated with tumor occurrence and progression. Therefore, multigene-based combination therapy may be an effective practice in cancer gene therapy, which can achieve greater therapeutic benefit by targeting multiple pathways. 30 Based on the antitumor features of ING4 and IL-24, the present study hypothesized that a combination of ING4 and IL-24 double tumor suppressors would elicit an enhanced antitumor efficacy in human cancers. In this report, recombinant adenoviruses coexpressing ING4 and IL-24 double tumor suppressor genes (Ad-ING4-IL-24) were constructed, its combined therapeutic effect on hepatocarcinoma cells in vitro and in vivo in an athymic nude mouse model was evaluated, and its underlying molecular mechanism was also elucidated.
Materials and Methods
Vectors, cell lines, reagents, and mice
The adenoviral transfer plasmid pAdTrack-CMV expressing green fluorescent protein (GFP) and Ad5E1- and E3-deleted backbone plasmid pAdEasy-1, 31 BJ5183 bacteria, and QBI-293A human embryonic kidney cell line were kindly provided by Dr. Jiang Zhong (Department of Microbiology, College of Life Science, Fudan University, Shanghai, China). The pGEZ-Term retroviral plasmid was kindly provided by Dr. Yongjing Chen (Biotechnology Institute, Soochow University, Suzhou, China). The pORF-mBcl-2a plasmid was purchased from InvivoGen (Shanghai, China). The SMMC-7721 (wild-type p53) and HepG2 (wild-type p53) human hepatocarcinoma cell lines and HL-7702 normal human liver cells were purchased from the American Type Culture Collection (Shanghai, China). The QBI-293A, SMMC-7721, HepG2, and HL-7702 cells were cultured in RPMI1640 (Gibco, Shanghai, China) supplemented with 10% fetal bovine serum (FBS) (Hyclone, Shanghai, China), respectively. The Lipofectamine2000, Trizol, and reverse transcriptase–polymerase MuMLV were purchased from Invitrogen (Shanghai, China). The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) kit, mammalian cell lysis kit, and mitochondria/cytosol isolation kit were purchased from Sigma (Shanghai, China). The Annexin V-PE/7-AAD apoptosis detection kit was purchased from BD Biosciences (Shanghai, China). A Transwell system was purchased from Corning Inc. (Shanghai, China). The monoclonal anti-IL-24 antibody and human IL-24, VEGF, and IL-8 enzyme-linked immunosorbent assay (ELISA) kits were purchased from R&D Systems (Shanghai, China). The polyclonal anti-ING4 antibody was purchased from Abcam (Shanghai, China). The antibodies specific for P53, P21, P27, Fas, FasL, FADD, Bid, Bad, Bcl-2, Bcl-XL, Bax, Bak, cytochrome c, Caspase-8, Caspase-9, Caspase-3, PARP, MMP-2, MMP-9, β-actin, CD34, VEGF, and IL-8 were purchased from Cell Signaling (Shanghai, China). The SuperEnhanced chemiluminescence detection kit was purchased from Applygen Technologies Inc. (Beijing, China). The UltraSensitiveTM SP kit was purchased from Maixin (Fuzhou, China). The TUNEL apoptosis detection kit was purchased from Beyotime Biotechnology (Beijing, China). The female athymic nude mice were purchased from Shanghai Experimental Animal Center (Shanghai, China) and maintained in the animal facility at Soochow University according to the animal research committee's guidelines of Soochow University.
Construction of recombinant adenoviruses coexpressing ING4 and IL-24 double tumor suppressor genes
Internal ribosome entry site (IRES) and multiple promoter expression cassettes are most popular strategies for multigene coexpression in a same vector. 32,33 Therefore, in the present study, recombinant adenoviral vectors coexpressing ING4 and IL-24 double tumor suppressor genes were constructed using the common modality diagrammed in Figure 1A. Briefly, the IRES fragments were amplified by polymerase chain reaction (PCR) using pGEZ-Term plasmids carrying IRES elements as templates and IRES-F (5′-acc gtc gac aat tcc gcc cct ctc cct ccc ccc ccc cta a-3′) and IRES-R (5′-gac gcg gcc gct tat cat cgt gtt ttt caa agg aaa acc ac-3′) as primers; the SV40 polyA + hEF1a-eIF4g promoter (termed polyA + prom) fusion fragments were amplified by splice overlapping extension (SOE)-PCR using pORF-mBcl-2a plasmids containing hEF1a-eIF4g promoter and SV40 polyA elements as templates and four primers as primers, namely, SV40 polyA-F (5′-ggg ttt ttt tat gga tct gcg atc gct ccg gt-3′), SV40 polyA-R (5′-gac gcg gcc gct ctc ctc tgt gat atc ctt tc-3′), hEF1a-eIF4g-F (5′-acc gtc gac aaa cct gcc cca aac aaa tatg-3′), and hEF1a-eIF4g-R (5′-cga tcg cag atc cat aaa aaa acc cgc cga ag-3′); and the ING4 and IL-24 cDNA fragments were amplified by PCR using pcDNA3-ING4 or pcDNA3-IL-24 plasmids as templates and primers specific for ING4 (ING4-F, 5′-tag aga tct acc atg gct gct ggg atg tat ttg g-3′; and ING4-R, 5′-acc gtc gac cct att tct tct tcc gtt ctt g-3′) or IL-24 (IL-24-F, 5′-gca ctc gag acc atg aat ttt caa cag agg ctg ca-3′; and IL-24-R, 5′-gct tct aga tca gag ctt gta gaa ttt ctg-3′) as primers. The ING4, IRES/polyA + prom, and IL-24 fragments were then subcloned into pAdTrack-CMV transfer plasmid expressing GFP at BglII, SalI; SalI, NotI; and XhoI, XbaI sites, respectively. After homologous recombination in BJ5183 bacteria, the recombinant adenoviruses Ad-ING4-IRES-IL-24 (Ad-ING4-IL-24I) and Ad-ING4-polyA + prom-IL-24 (Ad-ING4-IL-24P) were subsequently produced, abundantly amplified in QBI-293A cells, and purified by cesium chloride density-gradient ultracentrifugation as previously described. 31 The Ad-IRES (AdI), Ad-ING4-IRES (Ad-ING4I), and Ad-IRES-IL-24 (Ad-IL-24I); and Ad-polyA + prom (AdP), Ad-ING4-polyA + prom (Ad-ING4P), and Ad-polyA + prom-IL-24 (Ad-IL-24P) used as control adenoviruses were similarly prepared as described earlier. The titer of purified adenoviruses was determined using gene transfer unit (GTU) method by calculating the number of the reporter gene GFP-expressing QBI-293A cells within 18 hours after adenoviral infection under fluorescence microscopy. Accordingly, the ratio of infectious adenovirus (GTU) to target cells is called multiplicity of infection (MOI) in the present study.

Construction and identification of recombinant adenoviruses coexpressing inhibitor of growth 4 (ING4) and interleukin (IL)-24.
Analysis of adenovirus-mediated ING4 and IL-24 coexpression
To screen an optimal adenovirus coexpressing ING4 and IL-24 double tumor suppressor genes, the expression characterization of ING4 and IL-24 mediated by Ad-ING4-IL-24I and Ad-ING4-IL-24P in QBI-293A cells was compared. The QBI-293A cells were infected with Ad-ING4-IL-24I, Ad-ING4I, Ad-IL-24I, and AdI; and Ad-ING4-IL-24P, Ad-ING4P, Ad-IL-24P, and AdP at an MOI of 10 for 24 hours, respectively. The medium containing phosphate-buffered saline without adenovirus was used as a cell control (PBS control). The adenovirus-directed ING4 and IL-24 transgene expression in QBI-293A cells was analyzed by reverse transcription (RT)-PCR and western blot, respectively. To determine secreted expression of IL-24, the amount of IL-24 in the cellular supernatants generated from the infected and uninfected QBI-293A cells was further analyzed by ELISA using human IL-24 ELISA kit according to manufacturer's protocols. Based on the identification of adenovirus-mediated ING4 and IL-24 transgene expression in QBI-293A cells, the Ad-ING4-IL-24P (hereafter termed Ad-ING4-IL-24) adenovirus with two independent promoters driving ING4 (CMV) and IL-24 (hEF1a-eIF4g) expression and control adenoviruses Ad-ING4P, Ad-IL-24P, and AdP (hereafter termed Ad-ING4, Ad-IL-24, and Ad) were selected in the following functional assays. To assess the optimal MOI for a maximal infection and transgene expression in SMMC-7721, HepG2, and HL-7702 cells, the SMMC-7721 and HepG2 human hepatocarcinoma cells and HL-7702 normal human liver cells were infected with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, and Ad at various MOIs (0, 1, 10, 25, 50, 100, and 200) for 24 hours, respectively. The adenoviral infection efficiency was examined according to GFP expression by fluorescence microscopy. Further, the ING4 and IL-24 transgene expression mediated by adenoviral infection in SMMC-7721, HepG2, and HL-7702 cells was determined by RT-PCR, western blot, and ELISA analysis.
Cell viability assay
The in vitro suppressive effect of Ad-ING4-IL-24 on SMMC-7721 and HepG2 human hepatocarcinoma cells was assessed by MTT assay. Briefly, the SMMC-7721 and HepG2 human hepatocarcinoma cells and HL-7702 normal human liver cells (a normal liver cell control) were dispensed into 96-well culture plates at 1×104 cells per well, respectively. After 24 hours incubation at 37°C, the SMMC-7721, HepG2, and HL-7702 cells were infected with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad used as a blank adenovirus control at the respective optimal MOI of 50 (SMMC-7721), 25 (HepG2), and 25 (HL-7702) and cultured for the indicated time periods (0–4 days). The medium containing PBS without adenovirus was used as a cell control (PBS control). Before treatment and at different time points after treatment, the viability of SMMC-7721, HepG2, and HL-7702 cells was then analyzed using MTT kit according to manufacturer's protocols.
Flow cytometric analysis of apoptosis
The SMMC-7721 and HepG2 human hepatocarcinoma cells (1×106) were cultured with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad at MOIs of 50 (SMMC-7721) and 25 (HepG2) or without adenovirus for 48 hours, respectively. The treated and untreated SMMC-7721 and HepG2 tumor cells were harvested, washed with cold PBS, and then subjected to apoptosis analysis using Annexin V-PE (early apoptotic marker) and 7-AAD (late apoptotic marker) double staining following manufacturer's instructions by flow cytometry. Briefly, the tumor cells (1×105) were incubated in the presence of 5 μL Annexin V-PE and 5 μL 7-AAD in 100 μL of 1× Annexin V binding buffer at room temperature. After 15 minutes incubation, 400 μL of 1× binding buffer was added and the apoptotic cells were then analyzed by flow cytometry.
Tumor cell invasion assay
The effect of Ad-ING4-IL-24 on SMMC-7721 and HepG2 human hepatocarcinoma cell invasion in vitro was assessed using a Transwell system that incorporated a polycarbonate filter membrane with a diameter of 6.5 mm and pore size of 8 μm according to company's protocols. Briefly, the Matrigel (12.5 μg in 50 μL PBS) was added to the filter to form a thin gel layer, dried in a laminar hood overnight, and reconstituted in 100 μL PBS at 37°C for 2 hours. The Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721 and HepG2 tumor cells (1×105) were suspended in 100 μL of serum-free medium and added to the upper chamber of the Transwell insert. The lower chamber was filled with 500 μL of culture medium supplemented with 10% FBS. After 24 hours incubation at 37°C, the cells on the upper surface of the filter were removed using a cottons wab. The cells that penetrated to the lower surface of the filter were stained with hematoxylin and counted under an Olympus IX70 microscope in 10 randomized fields at 400× magnification. The assay was performed on three separate occasions.
Western blot analysis
The SMMC-7721 and HepG2 human hepatocarcinoma cells (1×107) were treated with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad at MOIs 50 (SMMC-7721) and 25 (HepG2) or without adenovirus, respectively. After 24 hours treatment, the Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721 and HepG2 tumor cells were collected, washed with cold PBS, and lysed in lysis buffer (1×107 cells/1 mL lysis buffer) for preparation of total cellular lysates using mammalian cell lysis kit. The mitochondrial and cytosolic protein fractions derived from treated and untreated SMMC-7721 and HepG2 tumor cells were also isolated using mitochondria/cytosol isolation kit according to manufacturer's protocols. The protein concentration was determined by BCA protein assay using spectrophotometer. The total cellular lysates (100 μg/lane) were then subjected to western blot analysis using a panel of antibodies specific for cell cycle- and apoptosis-related proteins such as P53, P21, P27, Fas, FasL, FADD, Caspase-8, Bid, Bad, Bcl-2, Bcl-XL, Bax, Bak, Caspase-9, Caspase-3, PARP, MMP-2, and MMP-9, respectively. Additionally, the mitochondrial and cytosolic proteins (100 μg/lane) were immunoblotted with cytochrome c antibody.
ELISA analysis
The SMMC-7721 and HepG2 human hepatocarcinoma cells were treated with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad at MOIs 50 (SMMC-7721) and 25 (HepG2) or without adenovirus, respectively. After 24 hours treatment, the numbers of infected and uninfected SMMC-7721 and HepG2 tumor cells were counted by trypan blue exclusion assay, and viable cells (5×105) separated with Ficoll separation medium (1.077) were further cultured for another 8 hours in 1 mL/well of fresh medium containing 10% FBS in a 24-well culture plate. The culture supernatants derived from Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721 and HepG2 tumor cells were then collected, and the amounts of proangiogenic factors such as VEGF and IL-8 in the supernatants were assessed by ELISA using human VEGF and IL-8 ELISA kits according to manufacturer's protocols.
Animal studies
The female athymic nude mice were subcutaneously (s.c.) inoculated on their armpits of right anterior limbs with 2×106 SMMC-7721 human hepatocarcinoma cells and then monitored daily for tumor growth. Tumor volume was measured with a caliper and calculated by the following formula: tumor size=ab 2 /2, where a is the larger and b is the smaller of the two dimensions. When the tumors grew up to a mean tumor volume of around 0.1–0.2 cm3, SMMC-7721 human hepatocarcinoma s.c. xenografted tumor-bearing mice were intratumorally (i.t.) injected with PBS (PBS control) or 1×108 GTU of Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, and Ad every other day for a total of six times, respectively. Tumor progression and regression were monitored and tumor volume was measured daily. In addition, the tumor-bearing mice (6 each group) were sacrificed at 2 weeks after treatment, and the xenografted tumors were removed, weighted, fixed by 10% neutral formalin, and embedded in paraffin for hematoxylin and eosin staining and immunohistochemistric analysis. In another set of experiment, animal tumor growth was monitored daily for up to 90 days after tumor cell inoculation, and survival period of the tumor-bearing mice (10 each group) was observed. For ethics reason, all mice with tumors that achieved a size of 1.5 cm in diameter were sacrificed.
Immunohistochemistric analysis
The expression of P53, P21, P27, Fas, FasL, Bcl-2, Bcl-XL, Bax, Bak, CD34, VEGF, and IL-8 in treated and untreated SMMC-7721 human hepatocarcinoma s.c. xenografted tumors was tested by immunohistochemistric analysis using UltraSensitive™ SP kit according to the company's protocols. The presence of buffy or brown diaminobenzidine precipitates is indicative of positive reactivity. The integral optical density (IOD) of immunohistochemical intensity was assessed by Image-Pro Plus 6.0 software. Microvessel density (MVD), detected by immunostaining for CD34, was determined as previously described by Weidner. 34 Any endothelial cell cluster immunoreactive for CD34 clearly separated from adjacent microvessels was considered as a single countable vessel. To detect the apoptotic cells in vivo in SMMC-7721 hepatocarcinoma xenografted tumors, tumor sections were further analyzed for apoptosis using TUNEL apoptosis detection kit according to manufacturer's instructions. Each value represents IOD, microvessels, or apoptotic cells counted at a high-power view (400×) by microscopy. The mean value represents for the average number derived from five high-power fields of each case.
Real-time RT-PCR analysis
To further confirm the effect of Ad-ING4-IL-24 on expression of VEGF and IL-8 in in vitro SMMC-7721 and HepG2 tumor cells and in vivo SMMC-7721 xenografted tumors, the total cellular RNA derived from Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721 and HepG2 human hepatocarcinoma cells and treated and untreated SMMC-7721 human hepatocarcinoma s.c. xenografted tumors were prepared for SYBR Green I-based real-time quantitative RT-PCR analysis using following primers: VEGF-F, 5′-tgg tag agt tca tgg atg tct atc a-3′ and VEGF-R, 5′-gca tgg tga tgt tgg act cct ca-3′; IL-8-F, 5′-atg act tcc aag ctg gcc gtg g-3′ and IL-8-R, 5′-tta tga att ctc agc cct ctt caa aa-3′; β-actin-F, 5′-tgc gtg aca tta agg aga ag-3′ and β-actin-R, 5′-ctg cat cct gtc ggc aat g-3′. The cDNA quantities were normalized to the internal control gene β-actin measured in the same samples. Relative gene expression was calculated using the pooled cDNA from all samples by the 2−ΔΔCT method as previously described. 35 The authenticity of the PCR products was verified by melting curve analysis and agarose gel electrophoresis. Each sample was analyzed in triplicate in independent reaction and the experiment was repeated three times.
Evaluation of combinatorial interaction
The interactive effects of ING4 and IL-24 by adenovirus-mediated ING4 and IL-24 coexpression were evaluated by Q-value calculated by the following formula 36 : Q=F(A + B)/FA +(1 − FA)FB, where F(A + B) represents the fraction affected by treatment with Ad-ING4-IL-24 compared with the untreated control group, FA represents the fraction affected by Ad-ING4 alone, and FB represents the fraction affected by Ad-IL-24 alone. A value of q>1.15 indicates a synergistic effect between ING4 and IL-24, q<0.85 indicates an antagonistic effect, and q between 0.85 and 1.15 indicates an additive effect.
Statistical analysis
All data are presented as the mean±standard deviation. The significance of the difference between groups was evaluated by one-way and two-way repeated measures analysis of variance and multiple comparisons with SPSS 10.0 software, and the animal survival in different groups was compared by log-rank test using Prism 3.0 software. A value of p<0.05 was considered statistically significant.
Results
Identification of recombinant adenoviruses coexpressing ING4 and IL-24
To compare coexpression characterization of ING4 and IL-24 double tumor suppressor genes induced by IRES element (CMV1-ING4-IRES-IL-24-polyA1) and independent promoter expression cassette (CMV1-ING4-polyA and hEF1a-eIF4g-IL-24-polyA1), the Ad-ING4-IL-24I- and Ad-ING4-IL-24P-mediated ING4 and IL-24 expression in QBI-293A cells was preliminarily side-by-side assessed by RT-PCR and western blot analysis. As shown in Figure 1B and C, significant amounts of ING4 and IL-24 expression at both transcriptional and translational levels were found in the Ad-ING4-IL-24I- and Ad-ING4-IL-24P-infected QBI-293A cells, whereas either ING4 or IL-24 expression was found in the Ad-ING4I or P- or Ad-IL-24I or P-infected QBI-293A cells but not in AdI or P-infected and uninfected QBI-293A control cells. Moreover, ING4-IRES-IL-24 transcriptional expression was found in Ad-ING4-IL-24I-infected QBI-293A cells, whereas there was no ING4-polyA-hEF1a-eIF4g-IL-24 RNA in Ad-ING4-IL-24P-infected QBI-293A cells (Fig. 1B), confirming that inserted hEF1a-eIF4g is a functional promoter with independently initiating transcriptional activity of downstream gene IL-24. ELISA analysis (Fig. 1D) further showed that secreted IL-24 was detected in the culture supernatants of QBI-293A cells infected with Ad-ING4-IL-24I or P or Ad-IL-24 I or P (p<0.05). Additionally, hEF1a-eIF4g promoter more efficiently induced downstream gene IL-24 translational expression than IRES element (Fig. 1C, D), although there was almost no difference in transcriptional activity between promoter hEF1a-eIF4g and CMV1 (Fig. 1B). These results indicated that two kinds of adenoviruses capable of coexpressing ING4 and IL-24 double tumor suppressor genes, Ad-ING4-IL-24I and Ad-ING4-IL-24P, were successfully constructed by genetically modification.
Adenovirus-mediated ING4 and IL-24 gene cotransfer
According to characterization of Ad-ING4-IL-24I- and Ad-ING4-IL-24P-mediated ING4 and IL-24 coexpression in QBI-293A cells, the Ad-ING4-IL-24P (hereafter termed Ad-ING4-IL-24) recombinant adenovirus coexpressing ING4 and IL-24 double tumor suppressor genes was preferably employed in gene therapy for hepatocarcinoma. The control adenoviruses Ad-ING4P, Ad-IL-24P, and AdP were correspondingly termed Ad-ING4, Ad-IL-24, and Ad. To determine the optimal MOI for a maximal transgene expression in hepatocarcinoma and normal liver cells, the SMMC-7721 and HepG2 human hepatocarcinoma cells and HL-7702 normal human liver cells were infected with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, and Ad at various MOIs for 24 hours and examined by fluorescence microscopy. More than 90% of GFP expression was found in the adenovirus-infected SMMC-7721, HepG2, and HL-7702 cells at MOIs of 50, 25, and 25 or above, whereas the GFP expression was not found in the uninfected SMMC-7721, HepG2, and HL-7702 control cells (data not shown). Therefore, 50, 25, and 25 MOIs were selected as an optimal dose for adenoviral infection and transgene expression of SMMC-7721, HepG2, and HL-7702 cells, respectively. To assess adenovirus-mediated ING4 and IL-24 transgene expression, the total cellular RNAs, lysates, and culture supernatants derived from Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721, HepG2, and HL-7702 cells were analyzed by RT-PCR (data not shown), western blot (Fig. 2A), and ELISA (Fig. 2B) (p<0.05), indicating that Ad-ING4-IL-24 harboring ING4 and IL-24 double tumor suppressor genes can also mediate ING4 and IL-24 coexpression in SMMC-7721 and HepG2 human hepatocarcinoma cells and HL-7702 normal human liver cells.

ING4 and IL-24 transgene expression in SMMC-7721, HepG2, and HL-7702 cells.
Synergistic tumor suppression by ING4 and IL-24 coexpression
To investigate whether combination treatment of ING4 and IL-24 tumor suppressors could exert enhanced antitumor activity, ING4 and IL-24 were coexpressed by adenovirus harboring ING4 and IL-24 double tumor suppressor genes (Ad-ING4-IL-24)-mediated gene cotransfer and its combined effect in SMMC-7721 and HepG2 human hepatocarcinoma cells and HL-7702 normal human liver cells was assessed. The SMMC-7721 and HepG2 hepatocarcinoma cells and HL-7702 normal liver cells were infected with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad at the respective optimal MOIs of 50, 25, and 25. The cell viability was examined daily for 4 days before and after treatment using MTT assay. As shown in Figure 3A, compared with the PBS and Ad control group, adenovirus-mediated ING4 and/or IL-24 expression significantly suppressed in vitro SMMC-7721 and HepG2 hepatocarcinoma cell growth in a time-dependant manner with peak inhibition at day 4 after infection (p<0.05), but exhibited minimal cytotoxicity on HL-7702 normal liver cells, indicating that ING4 and/or IL-24 transgene overexpression exerts selective tumor-killing activity in hepatocarcinoma cells. Further, combination treatment by ING4 and IL-24 coexpression resulted in a more significant and synergistic inhibition on the growth of SMMC-7721 and HepG2 tumor cells compared with the Ad-ING4- and Ad-IL-24-treated groups (p<0.05; SMMC-7721, Q=0.938, 1.157, 1.156, and 1.151, and HepG2, Q=1.116, 1.191, 1.172, and 1.179, at days 1, 2, 3, and 4 after treatment, respectively). To determine whether Ad-ING4-IL-24-mediated synergistic growth inhibition observed in vitro could be reproduced in vivo, combined effect of ING4 and IL-24 coexpression on hepatocarcinoma xenografted tumor growth was further evaluated by directly i.t. injecting Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad (1×108 GTU) into SMMC-7721 human hepatocarcinoma s.c. xenografted tumors in nude mice for a total of 6 times. The xenografted tumor growth in vivo was monitored daily, and tumor volume (Fig. 3B) and tumor weight (Fig. 3C) were measured. Compared with the Ad-ING4- and Ad-IL-24-treated groups, the SMMC-7721 hepatocarcinoma xenografted tumor growth in nude mice was also synergistically retarded in the Ad-ING4-IL-24-treated group (p<0.05; Q volume=1.106 and 1.183 at weeks 1 and 2 after treatment, and Q weight=1.169, respectively). In addition, all the mice injected with PBS or Ad died of SMMC-7721 hepatocarcinoma s.c. xenografted tumor within 36 days after tumor cell inoculation (Fig. 3D). Treatment of Ad-ING4 or Ad-IL-24 prolonged survival period of mice bearing SMMC-7721 hepatocarcinoma xenografted tumors, with 2–3/10 mice surviving within 90 days after tumor cell inoculation (Fig. 3D). Moreover, the tumor-bearing mice treated with Ad-ING4-IL-24 were able to survive much longer than those treated with Ad-ING4 or Ad-IL-24, with 6/10 mice surviving (Fig. 3D) (p<0.05; Q survival=1.364). These results indicated that Ad-ING4-IL-24 administration can remarkably suppress in vitro SMMC-7721 and HepG2 human hepatocarcinoma cell and in vivo in athymic nude mice SMMC-7721 human hepatocarcinoma xenografted tumor growth with synergistic effect, but no increasing toxicity in HL-7702 normal human liver cells.
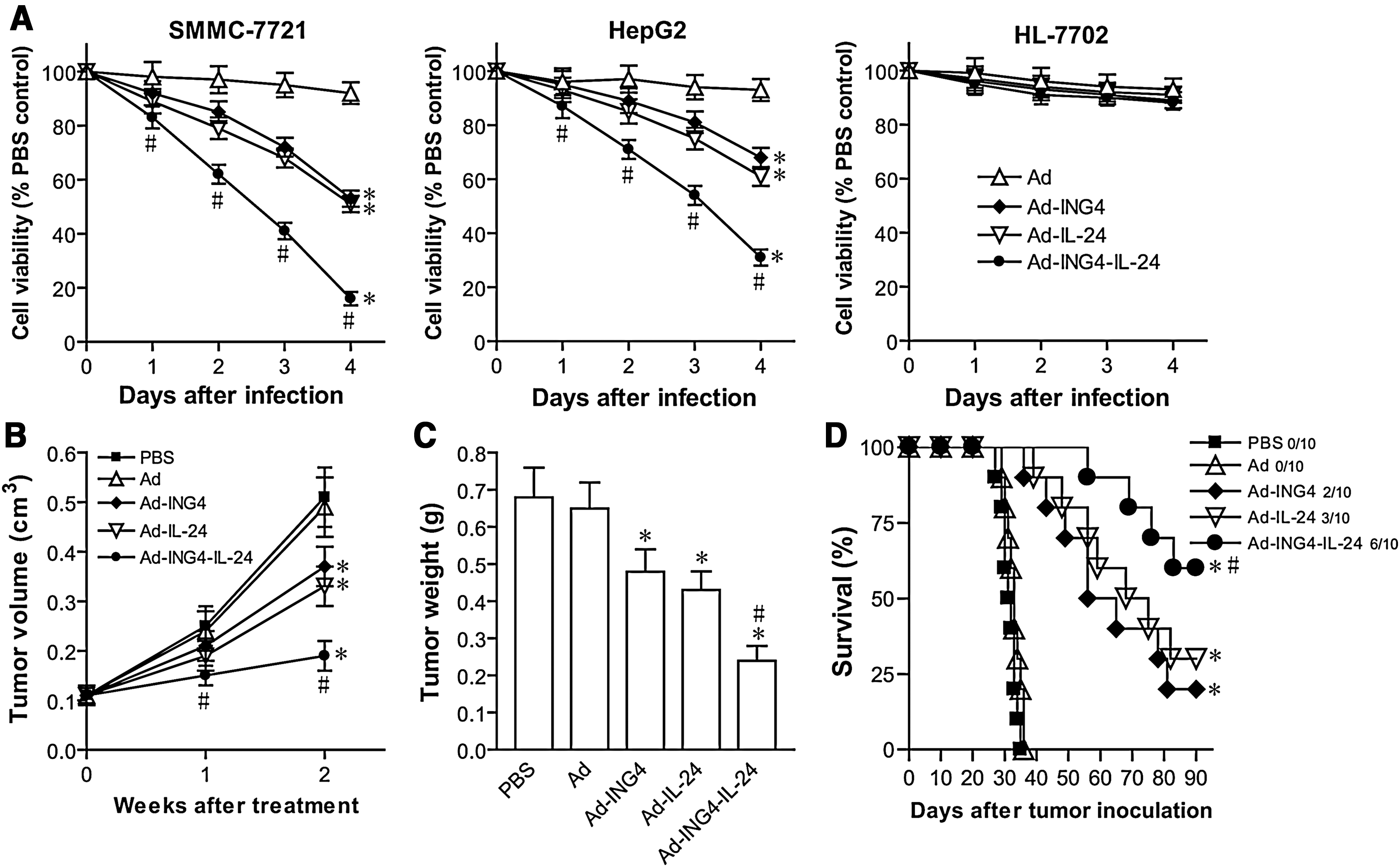
Ad-ING4-IL-24-induced synergistic tumor suppression in hepatocarcinoma cells.
Synergistic induction of apoptosis by ING4 and IL-24 coexpression
To explore the mechanism by which Ad-ING4-IL-24 synergistically inhibits tumor cell growth, the apoptosis of SMMC-7721 and HepG2 human hepatocarcinoma cells treated with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad for 48 hours were analyzed using Annexin V-PE and 7-AAD double staining by flow cytometry. As shown in Figure 4A, Ad-ING4-IL-24 treatment induced 62.8% and 48.3% apoptosis including early and late apoptosis in SMM-7721 and HepG2 hepatocarcinoma cells, whereas there was 2.8%, 3.5%, 29.2%, and 35.1% apoptotic SMMC-7721 tumor cells and 2.3%, 3.1%, 20.6%, and 25.9% apoptotic HepG2 tumor cells occurring in SMMC-7721 and HepG2 hepatocarcinoma cells grown in the medium containing PBS, Ad, Ad-ING4, and Ad-IL-24, respectively. Compared with the Ad-ING4- and Ad-IL-24-treated groups, adenovirus-mediated ING4 and IL-24 coexpression more efficiently induced SMMC-7721 and HepG2 hepatocarcinoma cell apoptosis with synergistic effect (p<0.05; SMMC-7721, Q=1.162, and HepG2, Q=1.173). To further confirm in vivo Ad-ING4-IL-24-induced apoptotic effect, the apoptosis in SMMC-7721 human hepatocarcinoma s.c. xenografted tumors was assessed by TUNEL assay (Fig. 4B, C). Consistent with the results in vitro by flow cytometric analysis, Ad-ING4-IL-24 also has a more effective and synergistic efficacy for in vivo inducing SMMC-7721 hepatocarcinoma cell apoptosis in nude mice with SMMC-7721 human hepatocarcinoma s.c. xenografted tumors (p<0.05; Q=1.253). These results showed that Ad-ING4-IL-24 significantly and synergistically induces SMMC-7721 and HepG2 human hepatocarcinoma cell apoptosis, leading to Ad-ING4-IL-24-mediated synergistic in vitro and in vivo growth inhibition of hepatocarcinoma cells.
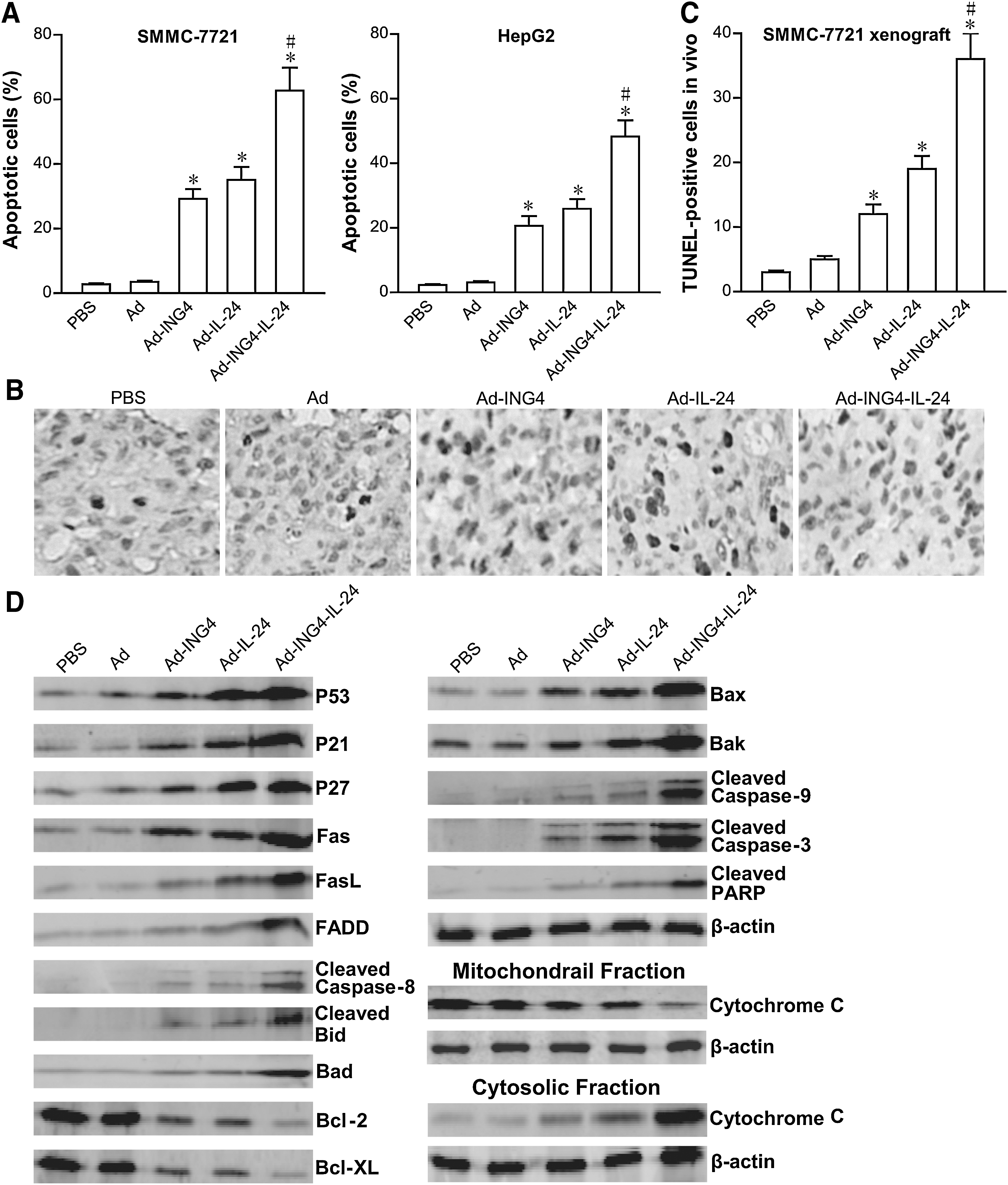
Ad-ING4-IL-24-elicted synergistic tumor apoptosis via cooperative activation of extrinsic and intrinsic apoptotic pathways.
Ad-ING4-IL-24 cooperatively regulates extrinsic and intrinsic apoptotic pathways
To further address the underlying molecular mechanism involved in Ad-ING4-IL-24-mediated synergistic antitumor effect, the expression of cell cycle- and apoptosis-related proteins such as P53, P21, P27, Fas, FasL, FADD, Caspase-8, Bid, Bad, Bcl-2, Bcl-XL, Bax, Bak, cytochrome c, Caspase-9, Caspase-3, and PARP in Ad-ING4-IL-24-, Ad-ING4-, Ad-IL-24-, or Ad-infected and uninfected SMMC-7721 and HepG2 human hepatocarcinoma cells was determined by western blot analysis. As shown in Figure 4D, the expression of P53 in the Ad-IL-24 and Ad-ING4-IL-24 groups and the expression of P21, P27, Fas, FasL, FADD, Bad, Bax, and Bak in the Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 groups was significantly increased, whereas the expression of Bcl-2 and Bcl-XL in the Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 groups was decreased. Additionally, cytochrome c significantly released from mitochondria and accumulated in the cytosol of Ad-ING4-, Ad-IL-24-, and Ad-ING4-IL-24-treated SMMC-7721 hepatocarcinoma cells. The cleavage of Caspase-8, Bid, Caspase-9, Caspase-3, and PARP was also found in the Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 groups but not in the PBS and Ad groups. Moreover, Ad-ING4-IL-24 treatment elicited an enhanced effect on the altered expression of P21, P27, Fas, FasL, FADD, Caspase-8, Bid, Bad, Bcl-2, Bcl-XL, Bax, Bak, cytochrome c, Caspase-9, Caspase-3, and PARP involved in the activation of extrinsic and intrinsic apoptotic pathways. A similar effect was also observed in HepG2 hepatocarcinoma cells (data not shown). The in vivo effect of Ad-ING4-IL-24 on expression of P53, P21, P27, Fas, FasL, Bcl-2, Bcl-XL, Bax, and Bak in SMMC-7721 human hepatocarcinoma xenografted tumors was further assessed and confirmed by immunohistochemical analysis (data not shown), which was consistent with above western blot analysis. These results indicated that Ad-ING4-IL-24 synergistically suppresses in vitro SMMC-7721 and HepG2 hepatocarcinoma cell and in vivo SMMC-7721 xenografted tumor growth and induces apoptosis closely associated with the cooperative regulation of extrinsic and intrinsic apoptotic pathways.
Ad-ING4-IL-24 downregulates VEGF and IL-8 expression involved in synergistic reduction of MVD
The positive expression of CD34 by immunohistochemical analysis was mainly presented as brownish yellow or brownish granules in vascular endothelial cells of SMMC-7721 human hepatocarcinoma s.c. xenografted tumors (Fig. 5A). Compared with the PBS and Ad groups, the CD34 expression of tumor vascular endothelial cells in the Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 groups was weaker or less (Fig. 5A, B), indicating that adenovirus-mediated ING4 and/or IL-24 expression can downregulate CD34 expression of SMMC-7721 human hepatocarcinoma xenografted tumor vessels. Additionally, the MVD (Fig. 5C) counted in the Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 groups was significantly less than that in the PBS and Ad control group (p<0.05). Further, Ad-ING4-IL-24 has a synergistic effect on the downregulation of CD34 and reduction of MVD in SMMC-7721 hepatocarcinoma xenografted tumors (p<0.05; Q CD34=1.194 and Q MVD=1.186), which may be involved in the Ad-ING4-IL-24-mediated in vivo synergistic growth inhibition of SMMC-7721 hepatocarcinoma xenografted tumor in nude mice. To elucidate the potential mechanism responsible for in vivo antiangiogenic effect of Ad-ING4-IL-24, first, the in vitro effect of Ad-ING4-IL-24 on the expression of proangiogenic factors VEGF and IL-8 in SMMC-7721 and HepG2 human hepatocarcinoma cells was examined by ELISA analysis. As shown in Figure 5D, the amounts of VEGF and IL-8 in Ad-IL-24- or Ad-ING4-IL-24-treated SMMC-7721 hepatocarcinoma cells were less than those in the cells treated with Ad or PBS (p<0.05), whereas Ad-ING4 only significantly downregulated IL-8 expression in SMMC-7721 hepatocarcinoma cells (p<0.05). Moreover, Ad-ING4-IL-24 had an overlapping effect on the downregulation of IL-8 but not VEGF in in vitro SMMC-7721 hepatocarcinoma cells (p<0.05; Q=1.050). A similar effect of Ad-ING4, Ad-IL-24, and Ad-ING4-IL-24 on in vitro VEGF and IL-8 expression was also observed in HepG2 hepatocarcinoma cells (data not shown). The in vivo expression of VEGF and IL-8 in SMMC-7721 hepatocarcinoma s.c. xenografted tumors with different treatments was further assessed by immunohistochemistric analysis. As shown in Figure 5E, the Ad-IL-24-induced in vivo downregulation of VEGF and IL-8 and the Ad-ING4-induced in vivo downregulation of IL-8 were consistent with in vitro ELISA analysis. Interestingly, Ad-ING4 was capable of downregulating VEGF expression in in vivo SMMC-7721 hepatocarcinoma xenografted tumors (Fig. 5E), suggesting that Ad-ING4 modulates VEGF expression dependent on the activity of VEGF upstream regulatory factors such as HIF-1a and NF-κB and VEGF basic level. Compared with the Ad-ING4- and Ad-IL-24-treated groups, Ad-ING4-IL-24 additively downregulated VEGF and IL-8 expression in in vivo SMMC-77721 hepatocarcinoma xenografted tumors (p<0.05; Q VEGF=1.039 and Q IL-8=1.110). The effect of Ad-ING4-IL-24 on VEGF and IL-8 expression in in vitro SMMC-7721 and HepG2 (data not shown) hepatocarcinoma cells and in vivo SMMC-7721 hepatocarcinoma xenografted tumors was further confirmed by real-time quantitative RT-PCR analysis (Fig. 5F). These results indicated that Ad-ING4-IL-24 synergistically inhibits tumor angiogenesis very possibly via indirectly and subtly downregulating the expression of VEGF and IL-8 proangiogenic factors.
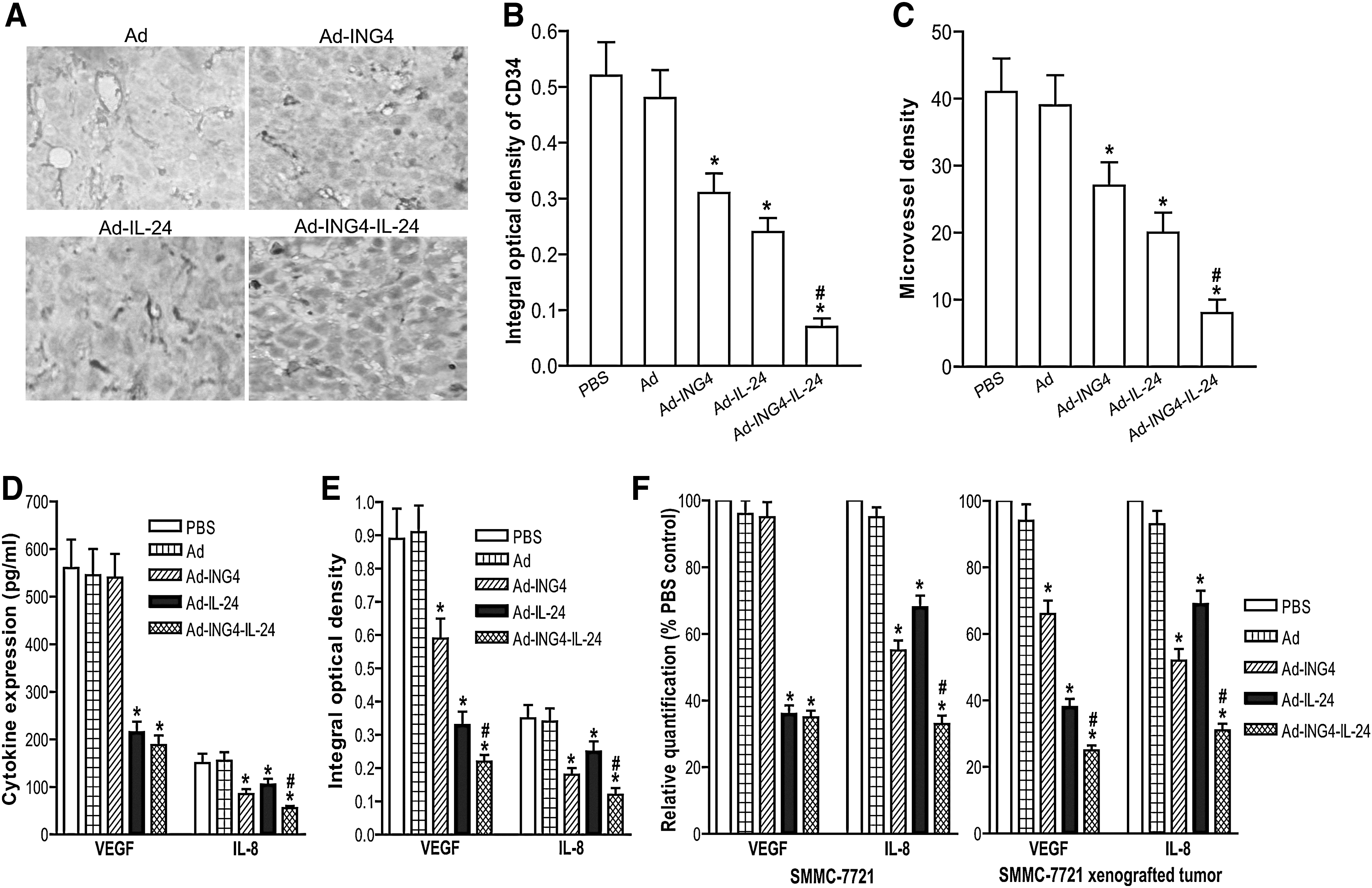
Ad-ING4-IL-24-mediated synergistic inhibition of tumor angiogenesis by downregulating vascular endothelial growth factor (VEGF) and IL-8 expression.
Ad-ING4-IL-24 synergistically inhibits tumor invasion associated with downregulation of MMP-2 and MMP-9
To examine whether adenovirus-mediated ING4 and IL-24 coexpression would elicit an enhanced inhibitory effect on hepatocarcinoma cell invasion, the in vitro invasive ability of SMMC-7721 and HepG2 human hepatocarcinoma cells infected with Ad-ING4-IL-24, Ad-ING4, Ad-IL-24, or Ad was assessed by Transwell invasion assay. As shown in Figure 6A, significant reductions in the invasive numbers of SMMC-7721 and HepG2 tumor cells were observed when those were infected with Ad-ING4-IL-24 (SMMC-7721, 85% reduction compared with invasive ability of PBS control cells; HepG2, 75% reduction), Ad-ING4 (SMMC-7721, 59% reduction; HepG2, 46% reduction), or Ad-IL-24 (SMMC-7721, 43% reduction; HepG2, 35% reduction) but not Ad (p<0.05). Moreover, Ad-ING4-IL-24 treatment more efficiently suppressed the invasiveness of SMMC-7721 and HepG2 tumor cells with a cooperative effect, compared with the Ad-ING4- and Ad-IL-24-treated groups (p<0.05; SMMC-7721, Q=1.161, and HepG2, Q=1.156). To address the molecular mechanism associated with the Ad-ING4-IL-24-induced anti-invasive effect, the effect of Ad-ING4-IL-24 on expression of invasion- and metastasis-related factors such as MMP-2 and MMP-9 in SMMC-7721 and HepG2 hepatocarcinoma cells was further investigated by western blot analysis. As shown in Figure 6B and C, treatment of SMMC-7721 tumor cells with Ad-ING4-IL-24 dramatically decreased the expression of MMP-2 and MMP-9 by 68% and 77%, respectively. There were also significant reductions of MMP-2 and MMP-9 expression in SMMC-7721 tumor cells infected with Ad-ING4 (MMP-2, 33% reduction; MMP-9, 62% reduction) or Ad-IL-24 (MMP-2, 58% reduction; MMP-9, 41% reduction) but not Ad (p<0.05). Significant downregulation of MMP-2 and MMP-9 was similarly observed in HepG2 tumor cells treated with Ad-ING4-IL-24 (MMP-2, 76% reduction; MMP-9, 61% reduction), Ad-ING4 (MMP-2, 42% reduction; MMP-9, 51% reduction), or Ad-IL-24 (MMP-2, 63% reduction; MMP-9, 29% reduction). Further, compared with the Ad-ING4- and Ad-IL-24-treated groups, Ad-ING4-IL-24 additively downregulated tumor metastasis-related factors MMP-2 and MMP-9 in SMMC-7721 (p<0.05; QMMP-2=0.946 and QMMP-9=0.993) and HepG2 (p<0.05; QMMP-2=0.968 and QMMP-9=0.935) tumor cells, which may be closely correlated with the Ad-ING4-IL-24-induced synergistic inhibition of invasion in hepatocarcinoma cells.
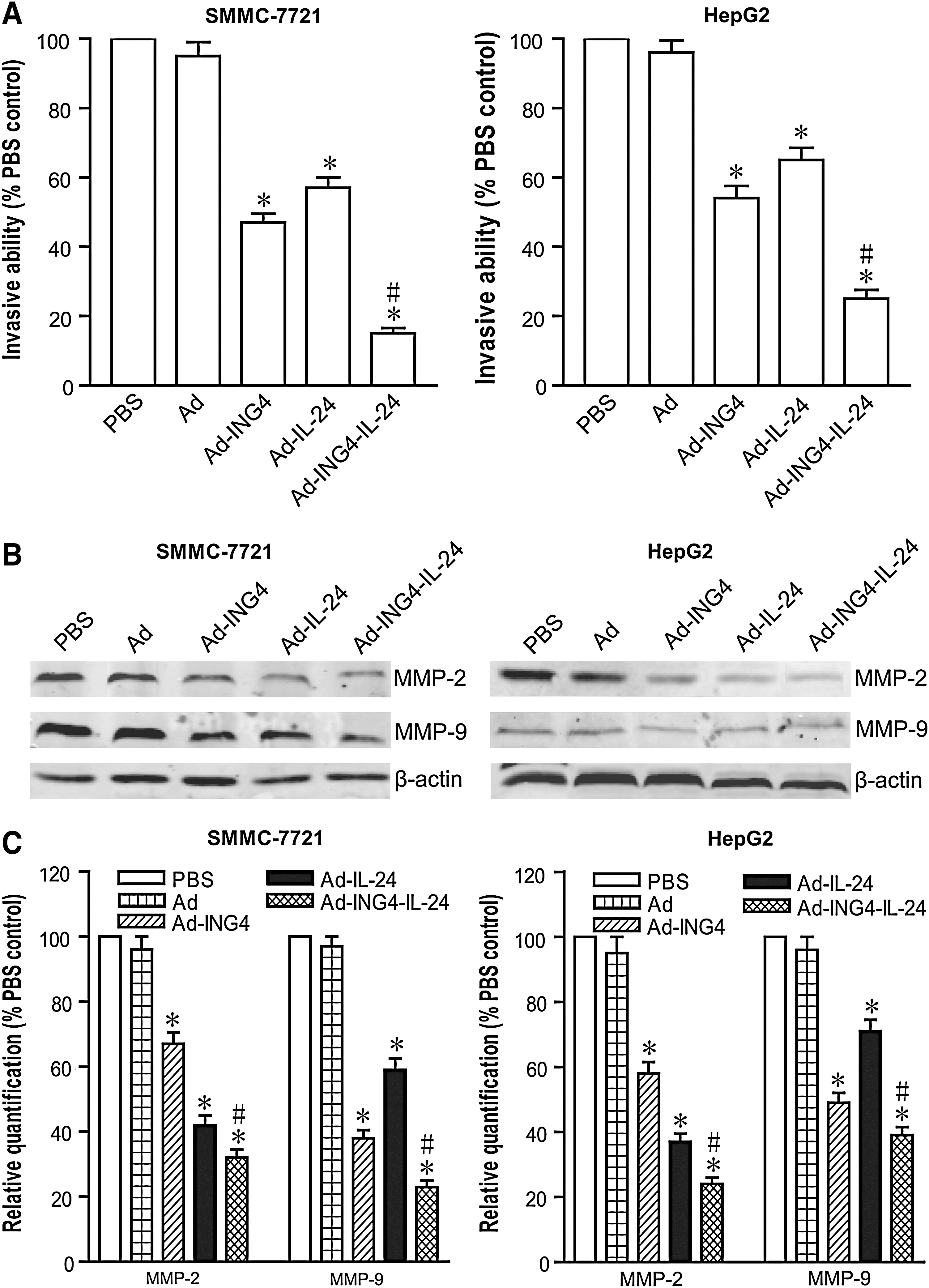
Ad-ING4-IL-24 synergistically suppresses tumor invasion associated with downregulation of matrix metalloproteinase (MMP)-2 and MMP-9.
Discussion
Recent studies have documented that ING4 as a novel tumor suppressor plays an important role in many cancer-related cellular processes including oncogenesis, cell cycle regulation, apoptosis, DNA damage response, invasion and migration, contact inhibition, and tumor angiogenesis, suggesting that it may serve as a potent tumor suppressor for cancer therapy. Expanding studies have demonstrated that the other tumor suppressor IL-24 can suppress tumor cell growth and induce apoptosis in a large spectrum of cancer cells via activation of double-stranded RNA-dependent protein kinase, p38 MAPK, c-Jun NH2-terminal kinase, and endoplasmic reticulum stress-mediated unfolded protein response signal pathways and inhibition of β-catenin and PI3K signal pathways. 37 In addition, IL-24 as a cytokine can be processed via class secretory pathways, bind specific cytokine receptor complexes (IL-20R1/IL-20R2 and IL-22R1/IL-20R2), and consequently activate Janus kinase/signal transducer and activator of transcription signal pathway. 37 The attributes of IL-24 to discriminate between normal and tumor cells, induce apoptosis, inhibit tumor angiogenesis, stimulate immune responses, promote bystander antitumor activity, and synergize with anticancer drugs and radiation warrant it as an effective agent for cancer treatment. Multigene-based combination therapy may be an effective practice in cancer gene therapy. Based upon the antitumor properties of ING4 and IL-24, it was speculated that the conjugation of ING4 and IL-24 double tumor suppressors would elicit enhanced tumor suppression. To develop better therapeutic strategies for HCC, in this study, recombinant adenoviruses coexpressing ING4 and IL-24 (Ad-ING4-IL-24) were constructed and its potential combined effect on SMMC-7721 and HepG2 human hepatocarcinoma cells in vitro and SMMC-7721 s.c. human hepatocarcinoma xenografted tumors in vivo in athymic nude mice was evaluated using adenovirus-mediated ING4 and IL-24 gene cotransfer.
The present study demonstrated that adenovirus-mediated ING4 and IL-24 coexpression induced in vitro synergistic growth inhibition and apoptosis in SMMC-7721 and HepG2 human hepatocarcinoma cells. Moreover, Ad-ING4-IL-24 treatment also synergistically suppressed in vivo hepatocarcinoma growth and induced apoptosis in SMMC-7721 human hepatocarcinoma xenografted tumors in nude mice. To elucidate the underlying mechanism involved in Ad-ING4-IL-24-mediated synergistic antitumor activity, the in vitro expression of cell cycle- and apoptosis-related proteins such as P53, P21, P27, Fas, FasL, FADD, Caspase-8, Bid, Bad, Bcl-2, Bcl-XL, Bax, Bak, cytochrome c, Caspase-9, Caspase-3, and PARP in SMMC-7721 and HepG2 hepatocarcinoma cells and the in vivo expression of P53, P21, P27, Fas, FasL, Bcl-2, Bcl-XL, Bax, and Bak in SMMC-7721 human hepatocarcinoma xenografted tumors were assessed by western blot and immunohistochemistric analysis, respectively. P53 as a transcription factor is an important tumor suppressor that can induce a permanent inhibition of cell growth through blocking cell cycle and activating apoptosis. 38 P21 and P27, important members of CDK inhibitors belonging to Cip/Kip family, can induce G1 and G2/M arrest. Fas and FasL as important apoptotic markers have been shown to regulate the cleavage of Caspase-8, leading to activation of extrinsic apoptotic pathway. 39 The ratio of anti- to proapoptotic Bcl-2 family proteins (Bcl-2, Bcl-XL; Bax, Bak) constitutes a rheostat that sets the threshold of susceptibility to apoptosis for the intrinsic pathway, which promotes pore formation in the mitochondrial outer membrane, leading to loss of mitochondrial integrity and the release of cytochrome c into the cytosol followed by the cleavage of Caspase-9 involved in the activation of intrinsic apoptotic pathway. 39 In addition, Bcl-2 family proapoptotic BH3-only protein Bad can activate multi-BH domain proapoptotic proteins Bax and/or Bak, which then allow for permeabilization of the mitochondrial membrane. 39 The present data showed that adenovirus-mediated ING4 and IL-24 coexpression elicited an enhanced effect on the altered expression of P21, P27, Fas, FasL, FADD, Bid, Bad, Bcl-2, Bcl-XL, Bax, and Bak, leading to the enhancement of cell cycle alteration and cooperative activation of extrinsic and intrinsic apoptotic pathways, which may be responsible for the Ad-ING4-IL-24-induced synergistic growth inhibition and apoptosis in SMMC-7721 and HepG2 tumor cells and SMMC-7721 xenografted tumors. Although Ad-ING4-IL-24 did not have an overlapping effect on increased P53 almost equal to that of Ad-IL-24, adenovirus-mediated ING4 and IL-24 coexpression dramatically upregulated the expression of P53-downstream genes P21 and Bax. The phenomenon may be explained by IL-24-induced P53 upregulation accompanied by ING4-mediated enhancement of acetylation and transcriptional activity of P53, 10 suggesting that adenovirus-mediated ING4 and IL-24 coexpression initiates synergistic tumor growth inhibition and apoptosis very possibly via their coordinate and functional links to P53.
The progressive growth and metastasis of solid tumors is dependent upon the process of angiogenesis. Tumor angiogenesis plays a critical role in the development and progression of HCC. It has been shown that ING4 can suppress tumor angiogenesis via downregulation of proangiogenic factors through inhibiting the activity of NF-κB and HIF-1a transcriptional factors implicated in cancers. 6,15 It has also been reported that IL-24 can inhibit tumor angiogenesis via directly interacting with IL-22R1/IL-20R2 on vascular endothelial cells 21 and indirectly reducing proangiogenic factors production. 20,22 To explore the combined effect of adenovirus-mediated ING4 and IL-24 coexpression on tumor angiogenesis in vivo, the MVD in SMMC-7721 human hepatocarcinoma s.c. xenografted tumor tissues was determined by CD34 immunohistochemical analysis. The present study found that Ad-ING4-IL-24 synergistically downregulated CD34 expression and decreased MVD in SMMC-7721 hepatocarcinoma xenografted tumors, which may be another important mechanism involved in Ad-ING4-IL-24-mediated in vivo synergistic growth inhibition of SMMC-7721 hepatocarcinoma xenografted tumors in athymic nude mice. Previous studies showed that VEGF and IL-8 are important proangiogenic factors involved in tumor angiogenesis. 40 –42 To address the potential mechanism of Ad-ING4-IL-24-mediated antiangiogenic regulation, the effect of Ad-ING4-IL-24 on the expression of VEGF and IL-8 in SMMC-7721 and HepG2 human hepatocarcinoma cells and SMMC-7721 hepatocarcinoma xenografted tumors was further examined by ELISA, immunohistochemical analysis, and real-time RT-PCR analysis. This study demonstrated that (1) Ad-ING4 downregulated IL-8 but not VEGF in in vitro SMMC-7721 and HepG2 tumor cells and VEGF and IL-8 in in vivo SMMC-7721 xenografted tumors; (2) Ad-IL-24 downregulated VEGF and IL-8 in in vitro SMMC-7721 and HepG2 tumor cells and in vivo SMMC-7721 xenografted tumors; and (3) Ad-ING4-IL-24 additively downregulated IL-8 in in vitro SMMC-7721 and HepG2 tumor cells and VEGF and IL-8 in in vivo SMMC-7721 xenografted tumors. The different effect of Ad-ING4 on VEGF expression in in vitro and in vivo indicated that Ad-ING4 downregulates VEGF expression dependent on the activity of VEGF upstream factors and VEGF basic level. Once VEGF upstream factors including HIF-1a and NF-κB were activated by cellular stress factors such as hypoxia, nutrient deprivation, and inducers of reactive oxygen species in tumor microenvironment, ING4 as a governor would exert reverse effect on VEGF production via a negative feedback mechanism. The difference between Ad-ING4- and Ad-IL-24-mediated effects on proangiogenic factors production also suggested that ING4 and IL-24 negatively regulate tumor angiogenesis via different pathways. Thus, Ad-ING4-IL-24-induced synergistic tumor angiogenesis suppression in SMMC-7721 hepatocarcinoma xenografted tumors may be closely associated with indirect and subtle downregulation of proangiogenic factors such as VEGF and IL-8.
Tumor metastasis is a fundamental and characteristic property of carcinogenesis, which involves adhesion, invasion, and migration. It has been demonstrated that both ING4 and IL-24 can significantly suppress tumor cell spreading, migration, and invasion. 8,12,25 The present study was also interested in assessing the effect of Ad-ING4-IL-24 on in vitro invasive ability of SMMC-7721 and HepG2 human hepatocarcinoma cells and its potential mechanism. The present study results showed that adenovirus-mediated ING4 and IL-24 coexpression synergistically suppressed invasion and additively downregualted MMP-2 and MMP-9 expression in SMMC-7721 and HepG2 tumor cells. The degradation of extracellular matrix and basement membrane is an initial and essential step in tumor cell invasion and metastasis and is mainly mediated by MMPs. 43 In particular, MMP-2 and MMP-9 are strongly implicated in invasion and metastasis of malignant tumors. 44,45 Therefore, additive downregulation of MMP-2 and MMP-9 elicited by Ad-ING4-IL-24 in SMMC-7721 and HepG2 hepatocarcinoma cells may contribute to Ad-ING4-IL-24-induced synergistic invasion inhibition of hepatocarcinoma cells.
Taken together, adenovirus-mediated ING4 and IL-24 coexpression induced synergistic growth inhibition, apoptosis, invasion suppression, and an enhanced effect on upregulation of P21, P27, Fas, FasL, FADD, Bad, Bax, Bak, cleaved Bid, cleaved Caspase-8, −9, and −3, and cleaved PARP, downregulation of Bcl-2, Bcl-XL, MMP-2, MMP-9, and IL-8, and cytochrome c release from mitochondria into cytosol in SMMC-7721 and HepG2 human hepatocarcinoma cells. Moreover, Ad-ING4-IL-24 treatment synergistically suppressed in vivo SMMC-7721 hepatocarcinoma xenografted tumor growth and induced apoptosis, synergistically or additively upregulated P21, P27, Fas, FasL, Bax, and Bak expression, downregulated Bcl-2, Bcl-XL, CD34, and MVD, and reduced expression of VEGF and IL-8 in SMMC-7721 xenografted tumors in athymic nude mice. The in vitro and in vivo synergistic antitumor activity elicited by Ad-ING4-IL-24 was closely associated with the cooperative activation of extrinsic and intrinsic apoptotic pathways and reduced proangiogenic factors production of VEGF and IL-8, leading to synergistic inhibition of tumor angiogenesis. Thus, the present study results indicate that cancer gene therapy combining two or more tumor suppressors such as ING4 and IL-24 may constitute a novel and effective therapeutic strategy for hepatocarcinoma and other cancers.
Footnotes
Acknowledgments
This research work was supported by grants from the National Natural Science Foundation of China (No. 81001016) and Medicine Research Foundation of Department of Public Health of Jiangsu Province (No. H200914).
Disclosure Statement
The authors have no financial conflicts of interest.