
Abstract
Using a uterine cervical cancer cell line expressing human papillomavirus (HPV) 16 E7 antigen and bioluminescent imaging (BLI), we evaluated the therapeutic potential of combined immunotherapy using transfected dendritic cells (DC-E7) and human sodium/iodide symporter (hNIS) radioiodine gene therapy in a xenograft animal cancer model. Dendritic cells expressing either E7 antigen (DC-E7) or no-insert (DC-no insert) were made for immunization materials, and murine uterine cervical cancer cell line coexpressing E7, firefly luciferase, hNIS, and EGFP genes (TC-1/FNG) were prepared for the animal tumor model. C57BL/6 mice were divided into five therapy groups (phosphate-buffered saline [PBS], DC-no insert, DC-E7, I-131, and DC-E7+I-131 groups). Single therapy with either DC-E7 or I-131 induced greater retardation in tumor growth compared with PBS or DC-no insert groups, and it resulted in some tumor-free mice (DC-E7 and I-131 groups, 40% and 20%, respectively). Combination therapy with DC-E7 and I-131 dramatically inhibited tumor growth, thus causing complete disappearance of tumors in all mice, and these effects were further confirmed by BLI in vivo. In conclusion, complete disappearance of the tumor was achieved with combined DC-E7 vaccination and hNIS radioiodine gene therapy in a mouse model with E7-expressing uterine cervical cancer, and serial BLIs successfully demonstrated antitumor effects in vivo.
Introduction
Dendritic cells (DCs) are potent antigen-presenting cells (APCs) that prime helper and killer T cells in vivo. 1 –3 Since DCs highly express MHC class I and II molecules, costimulatory molecules (B7), and adhesion molecules (ICAM-1, ICAM-3, and LFA-3), they can stimulate T cells (CD4 and CD8 T cells), which have an important role in cancer immunotherapy. Due to their unique characteristics, dendritic cells (DCs) have been used to identify tumor-specific antigens, such as human papillomavirus (HPV)-16 E6/E7, or with undefined tumor antigens for cancer immunotherapy. 4 Although DC vaccines have been applied to various tumors, they have only been partially effective. To strengthen the effects of the DC vaccine, several strategies have been used: (1) DCs were pulsed with peptides or proteins, 2,5 –7 (2) DCs were transfected with nonviral or viral DNA encoding a tumor antigen, 8 –11 and (3) they were armed with irradiated tumor cells. 12,13 Although these strategies have achieved their preclinical or clinical aims by the generation of tumor-specific (or associated) immune responses and antitumor effects, they are not always effective due to their rather weak potency and various hindrances, such as regulatory T-cell hindrance or cytokine imbalance in the host. 14
Sodium iodide symporter (NIS) is a specialized active iodide transporter that cotransports sodium and iodide ions. 15,16 Many investigators have attempted to apply this promising system to cancer gene therapy by directly transfecting the NIS gene using a viral or non-viral vector system into the tumor bearing animal model. NIS gene expression in cancer cell allows the accumulation of therapeutic radioisotope (Re-188 and I-131), which enables the successful production of therapeutic effects in various cancer models. 17 –19 NIS gene therapy with tissue-specific promoter has allowed the accumulation of therapeutic radioisotope within transfected cells and, consequently, generates a striking reduction of tumor volume in athymic nude mice. 20,21
We previously demonstrated several possible therapeutic applications of human NIS (hNIS) radioiodine gene therapy with alternative strategies, such as synergistic tumoricidal effects induced by combined hNIS radioiodine gene therapy and DNA vaccination, 22 and with improved killing effects of cytotooxic T cells by hNIS radioiodine gene therapy. 23 These findings suggest that hNIS radioiodine gene therapy could be combined with other immunological therapeutic strategies including cancer DC vaccine, cytokine administration, tumor-derived peptide vaccine, and so on.
Since hNIS radioiodine gene therapy is known to be a powerful anticancer therapeutic tool with the ability to increase cytotoxic T-cell-mediated immunity against cancer, we postulated that combined DC-based immunotherapy and hNIS radioiodine gene therapy could be an effective therapeutic strategy for aggressive and low immunogenic cancers that do not respond to single therapy. Since two HPV oncogenic proteins, E6 and E7, are low immunogenic tumor-specific antigens, the generation of a cellular immune response using a DC vaccine is very difficult. Thus, we applied our combination therapy to uterine cervical cancer models, which express HPV E7 protein (referred to as the TC-1 cancer cell line). The TC-1 cell line has proved to be an appropriate cancer model for investigating the efficacy of immunotherapeutic reagents, such as DNA vaccine, viral vaccine, and the DC vaccine expressing HPV oncogenic protein E7. 24 –26
In this study, we explored the therapeutic potential of combined DC (transfected with E7) vaccination and hNIS radioiodine gene therapy in an animal model transplanted with a low immunogenic cancer, and its therapeutic response was monitored using bioluminescent imaging (BLI) in vivo.
Materials and Methods
Animals
Specific pathogen-free, 6-week-old female C57BL/6 mice were obtained from SLC, Inc. All animal experiment protocols were approved by the Committee for the Handling and Use of Animals of the University.
DC vaccines
DC vaccine with or without E7 (DC-E7 and DC-no insert) was kindly provided by Dr. T. W. Kim at Korea University. Briefly, the immortalized DC2.4 cell line 27 was genetically manipulated using a retroviral system. This DC2.4 cell line was used to construct the DC vaccines using a retroviral vector plasmid. Phoenix packaging cells were transfected with retroviral vector plasmids, pMSCV and pMSCV-E7, using a Lipofectamine 2000 (Invitrogen). Retroviral supernatant from the transfected phoenix cells was incubated with 50% confluent DC in the presence of 8 μg/mL polybrene (Sigma). Transduced cells were grown in a culture medium containing 7.5 μg/mL puromycin for selection. The expression of the E7 antigen was previously confirmed by Western blot analysis. 28
Establishment of a stable cancer cell line coexpressing E7, Fluc, and hNIS gene
Murine cervical cancer cells expressing the Fluc gene for BLI (TC-1/Fluc cells) were kindly provided by Dr. T.C. Wu. 29 TC-1/Fluc cells were transduced with a retrovirus coexpressing hNIS and GFP genes. Two (2) days later, GFP positive cells were enriched using FACSaria (BD Bioscience). The selection procedure was performed twice over 4 weeks. After selection of stable cells, flow cytometry analysis and a luciferase assay were conducted to determine the gene expression of GFP and Fluc using FACSaria and a microplate luminometer (Molecular Devices). The expression of E7 mRNA and protein in TC-1 cells also was previously confirmed by RT-PCR and immunofluorescent staining. 30
Immunofluorescent staining
TC-1 and TC-1/FNG cells were seeded at a density of 4×104 cells per well on collagen-precoated chamber slides (Nunc) and grown for 24 hours. The hNIS-specific immunostaining was performed with a Cytofix/cytoperm kit (BD Pharmingen). The cells were washed twice with phosphate-buffered saline (PBS). The washed cells were fixed and permeabilized with fixation/permeabilization buffer (eBioscience) for 20 minutes at 4°C. Afterwards, the cells were washed with 1xBD Perm/Wash™ buffer (BD Bioscience) and stained with anti-hNIS antibody (Millipore) diluted with 1xBD Perm/Wash buffer for 1 hour at room temperature. After the cells had been washed thrice with 1xBD Perm/Wash buffer, Alexa fluor 568-conjugated secondary antibody (Molecular Probes, Inc.) was applied for 40 minutes at room temperature. The cells were washed thrice with 1xBD Perm/Wash buffer. The slides were mounted with Vectashield Mounting Medium (Vector Laboratories), covered with glass cover slips, and examined via a laser confocal scanning system with Leica TCS SP2 (Leica).
Radioiodide uptake assays
TC-1 and TC-1/FNG cells (5×104, 1×105, 3×105, and 5×105) were plated in 24-well plates and cultured with DMEM containing 10% FBS for 24 hours. I-125 uptake was determined by incubating the cells with 500 μL of Hanks balanced salt solution (bHBSS; GibcoBRL Co.) containing 3.7 kBq (0.1 μCi) of carrier-free I-125 NaI and 10 μM NaI at 37°C for 30 minutes to yield a specific activity of 740 MBq/mmol (20 mCi/mmol). After incubation, the cells were quickly washed twice with bHBSS and detached using 500 μL of trypsin. Radioactivity was measured using a gamma counter (CobraII, Packard; Perkin Elmer). For the inhibition study, radioiodine uptake was measured after pretreatment with 50 μM potassium perchlorate for 1 hour, and cellular radioactivity was measured.
Iodine uptake was measured at various time points, as just described. Cells (200,000) were plated and incubated with 500 μL of bHBSS containing 3.7 kBq (0.1 μCi) carrier-free I-125 NaI and 10 μM NaI at different time points, to yield a specific activity of 740 MBq/mmol (20 mCi/mmol).
For the iodine efflux study, cells were incubated with 500 μL of bHBSS containing 3.7 kBq (0.1 μCi) of carrier-free I-125 NaI and 10 μM NaI at different time points, to yield a specific activity of 740 MBq/mmol (20 mCi/mmol). Cells were washed twice and incubated with bHBSS at 37°C. bHBSS was removed at different time points, and its radioactivity was measured. Cells were collected after the last time point, and residual radioactivity was measured in counts per minute. All data were expressed as the percentage remaining for the total activity.
In vitro clonogenic assay
For I-131 treatment in vitro, 5×106 TC-1 and TC-1/FNG cells were seeded in T75-tissue culture flasks. The cells were washed once with 5 mL of HBSS and incubated with 5 mL of HBSS (control) or 5 mL of HBSS containing 18.5 MBq of I-131. After incubating with I-131 for 7 hours, cells were washed twice with cold HBSS, trypsinized, and counted. One thousand cells were plated in triplicate in six-well plates and incubated for 1 week at 37°C. Cells were washed once with PBS and stained with a crystal violet solution (per 250 mL, 0.5 g of crystal violet, 25 mL of 40% formaldehyde, 50 mL of ethanol, and 175 mL of water). Colonies of more than 30 cells were counted, and all values were adjusted for plating efficiency. Cell survival was expressed as the percentage of colonies relative to that in the untreated control.
Nuclear imaging with Tc-99m pertechnetate
For the in vivo imaging study, 1×105 TC-1 and TC-1/FNG cells in a volume of 0.1 mL were subcutaneously implanted into the left and right thighs of C57BL/6 mice (Group 1, n=6). Fourteen days after inoculation, 18.7 MBq (500 μCi) of Tc-99m pertechnetate in 100 μL of PBS was intravenously injected, and a whole-body scan was performed 30 minutes post injection using a gamma camera equipped with a pinhole collimator (Infinia II; GE). After data acquisition, tumoral uptake was quantified from regions of interest (ROIs) over the tumor region. Mice were sacrificed after imaging, and tumor weight was measured.
In vivo combination therapy and BLI
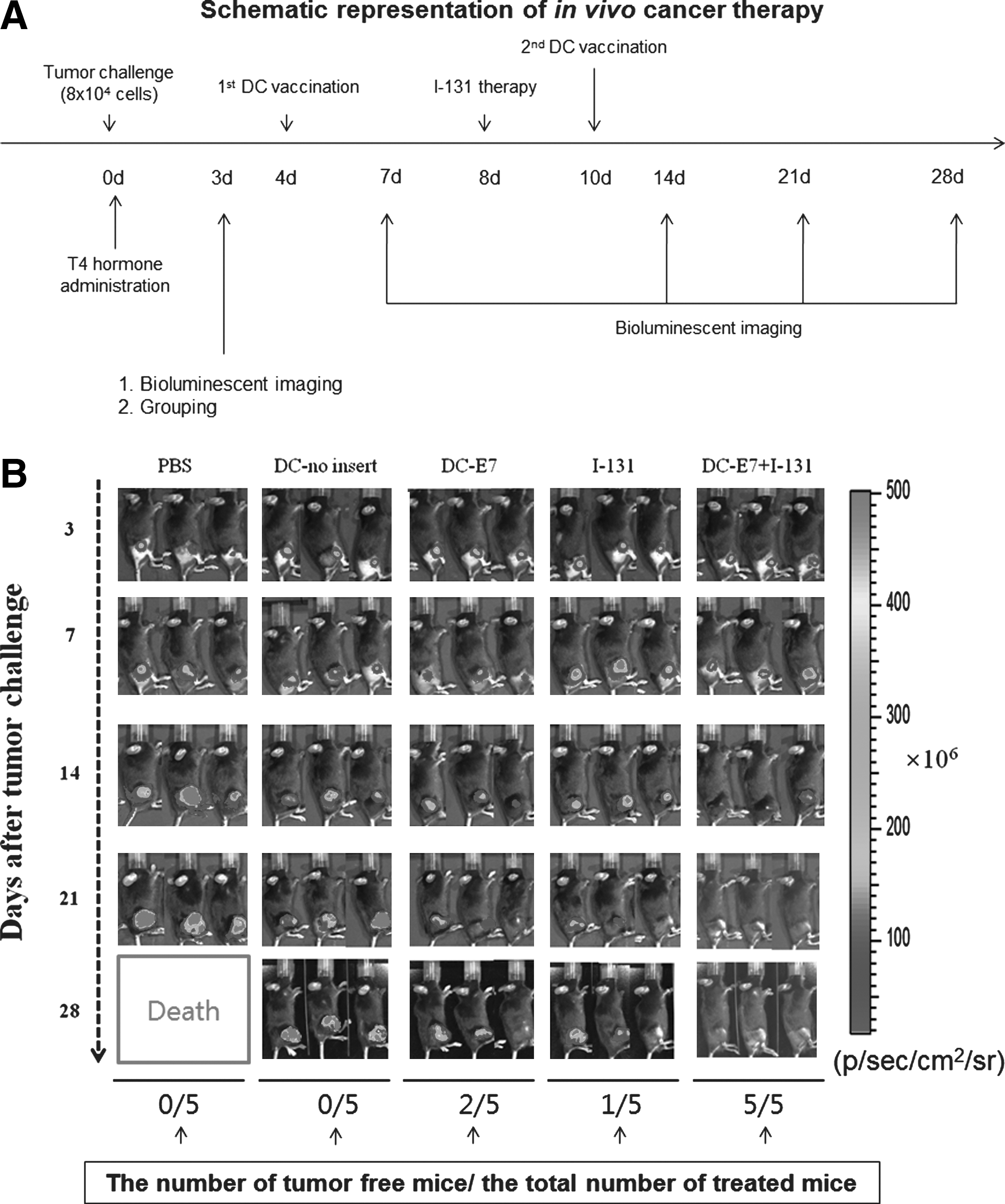
The scheme of the in vivo cancer therapy is described in Figure 5A. Briefly, C57BL/6 mice were divided into five groups according to various therapy protocols (PBS as a negative group, DC-no insert as a mock group, DC-E7 as a immunotherapy group, I-131 as a radiation therapy group, DC-E7+I-131 as a combination therapy group; five mice per group), and 8×104 TC-1/FNG cells were subcutaneously challenged into the right hind limb of each mouse. On the fourth and tenth days, mice were intramuscularly injected with 1×106 DC-no insert cells or DC-E7 cells. A single injection of I-131 (37 MBq) was intravenously administered into mice of I-131 and DC-E7+I-131 group at seven days afterwards. Tumor growth was monitored with palpation and BLI at 3, 7, 14, 21, 24, and 28 days. An IVIS Lumina II (Caliper Life Sciences) was used for BLI acquisition and analysis.
Statistical analysis
Data are presented as the mean±standard deviation. Intergroup differences were assayed using the two-tailed Student's t-test. Tumor-free mice were determined using the Kaplan–Meier curve and log-rank test. p-Values<0.05 were considered statistically significant.
Results
Assessment of hNIS and GFP gene expression in TC-1 and TC-1/FNG cells
Protein expression for the hNIS and EGFP genes in a single retroviral construct was analyzed using flow cytometry and immunofluorescent staining. Flow cytometric studies revealed an enriched GFP-positive cell fraction in TC-1/FNG cells but not in TC-1 cells (Fig. 1A). hNIS-specific immunoreactivity was detected in TC-1/FNG cells by immunofluorescent staining but not in parental TC-1 cells (Fig. 1B). Isotype control did not show any hNIS-specific immunoreactivity in TC-1/FNG cells.
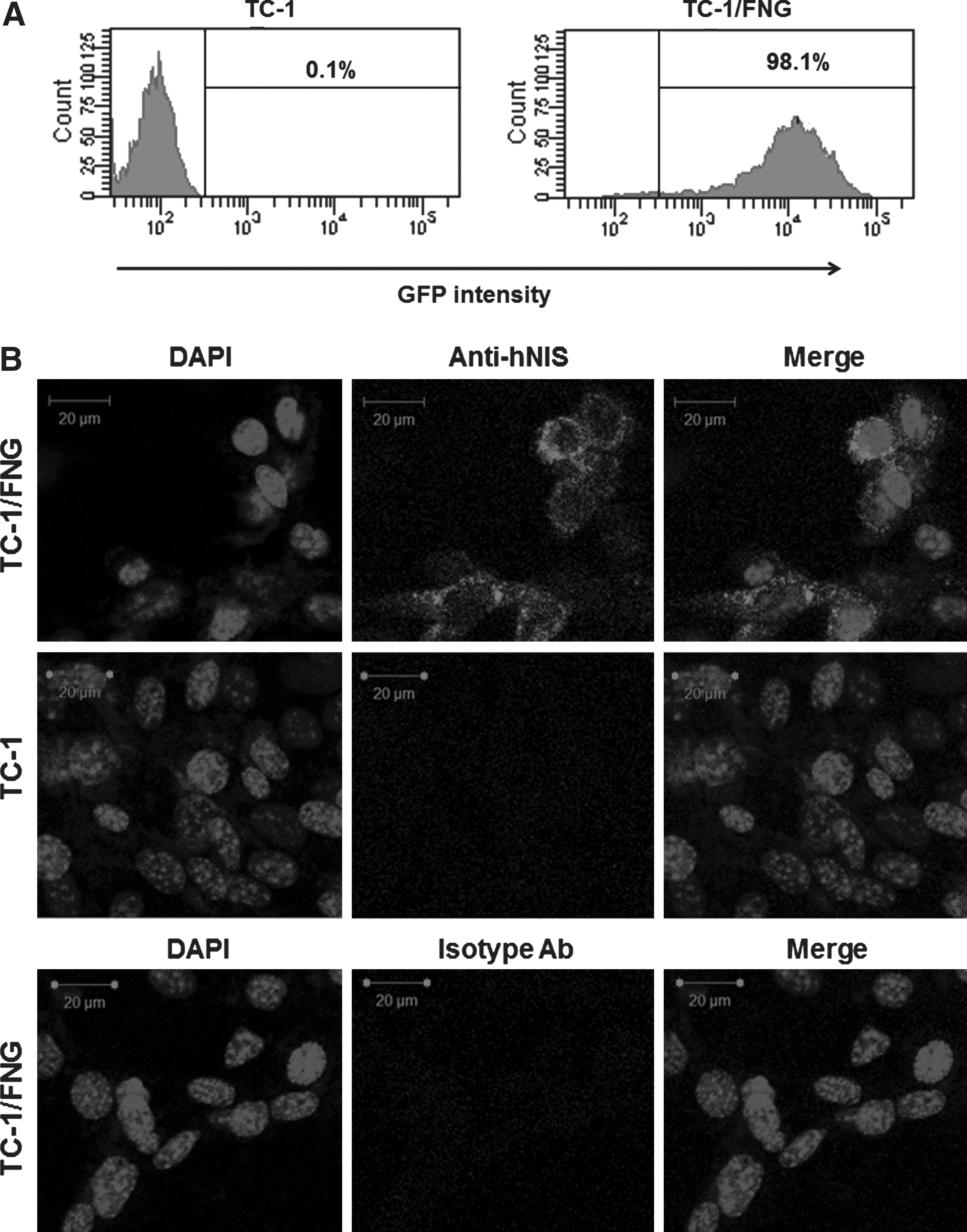
Establishment of a stable cell line coexpressing Fluc, hNIS, and GFP genes.
In vitro I-125 kinetics in TC-1/FNG cells
I-125 uptake increased proportionally with increases in TC-1/FNG cell number but not for parental TC-1 cells (Fig. 2A, upper panel). I-125 uptake in TC-1/FNG cells was ∼90-fold higher than in TC-1 cells, and it was almost completely inhibited by perchlorate, a competitive NIS inhibitor. The accumulation of I-125 in TC-1/FNG cells reached a plateau at 15 minutes and remained high for the 60 minutes measured (Fig. 2A, middle panel). The efflux of I-125 was relatively rapid, and the half time was ∼4 minutes (Fig. 2A, bottom panel). Bioluminescent signals correlated with cell number in TC-1/FNG cells but not in TC-1 cells (Fig. 2B, R 2 =0.99, p<0.001).

Validation of functional activity of hNIS and Fluc protein in vitro. In vitro kinetics of I-125 in TC-1/FNG cells.
In vitro cytotoxic effects of I-131 in TC-1/FNG cells
As shown in Figure 3, clonogenic assay of TC-1/FNG cells demonstrated that 18.5 MBq (500 μCi) of I-131 treatment resulted in fewer colony formations. Percentage survival of TC-1/FNG cells and parental TC-1 cells was 38.0%±4.0% and 92.0%±5.0%, respectively, with I-131 treatment (p<0.05).

In vitro cytotoxicty of I-131. Detailed procedure for clonogenic assay is described in Materials and Methods section. Results were expressed as mean±SD of three experiments. *p<0.05 compared with parental TC-1 cell treated I-131.
Effects of viral infection on tumor growth and nuclear imaging for NIS gene expression with Tc-99m pertechnetate
As shown in Figure 4A, there was no significant difference in tumor weight between parental TC-1 cells and TC-1/FNG cells at 14 days post-tumor challenge. Scintigraphic imaging demonstrated a higher uptake of Tc-99m pertechnetate in TC-1/FNG tumors than in the parental TC-1 tumor, which was shown to be 4.3-fold higher by quantitative analysis (Fig. 4B, p<0.05).
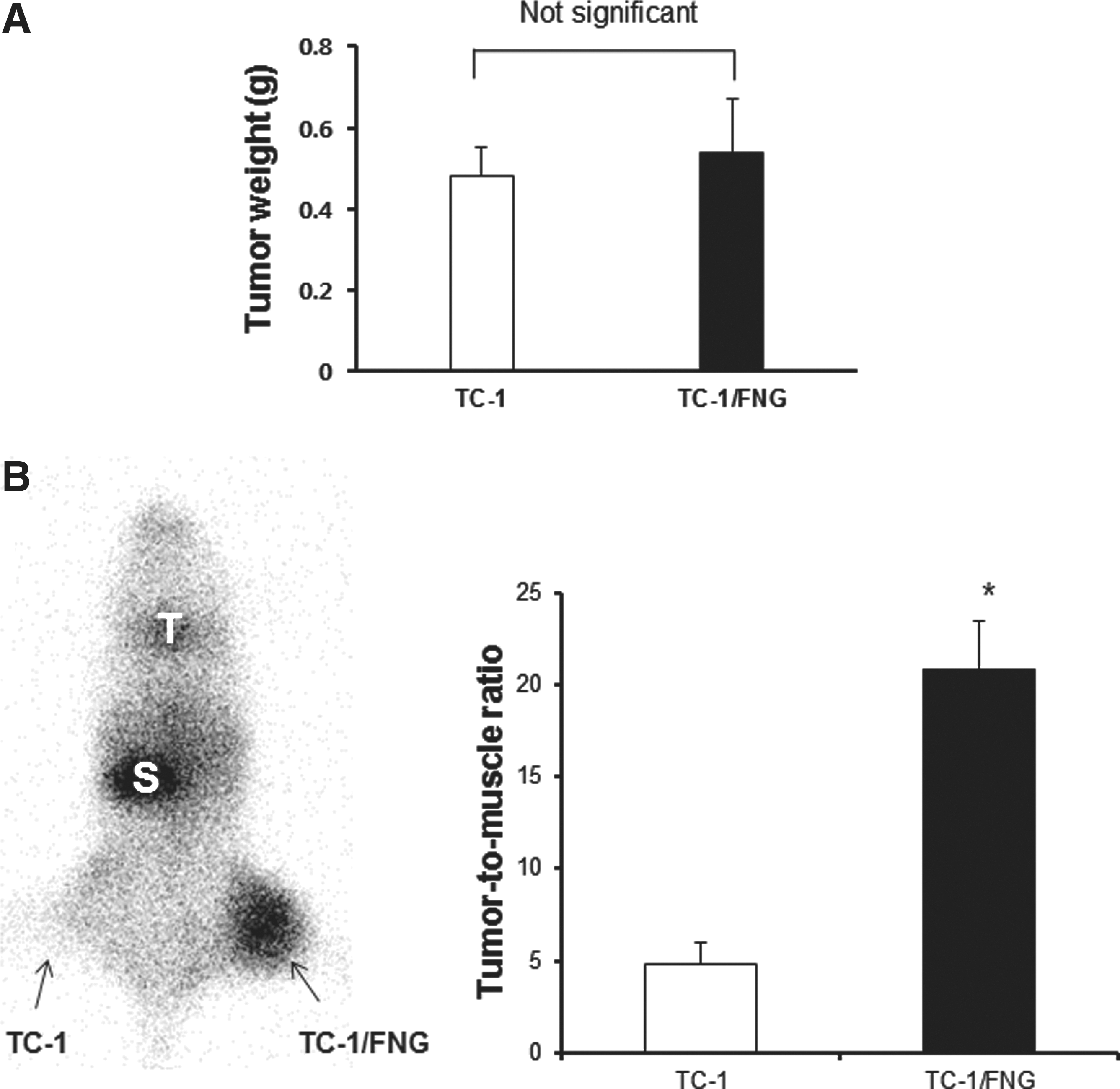
Tumor growth kinetics of TC-1/FNG cells and scintigraphic imaging with Tc-99m pertechnetate in vivo.
In vivo effects of combination therapy on tumor growth and BLI for the evaluation of its therapeutic effects
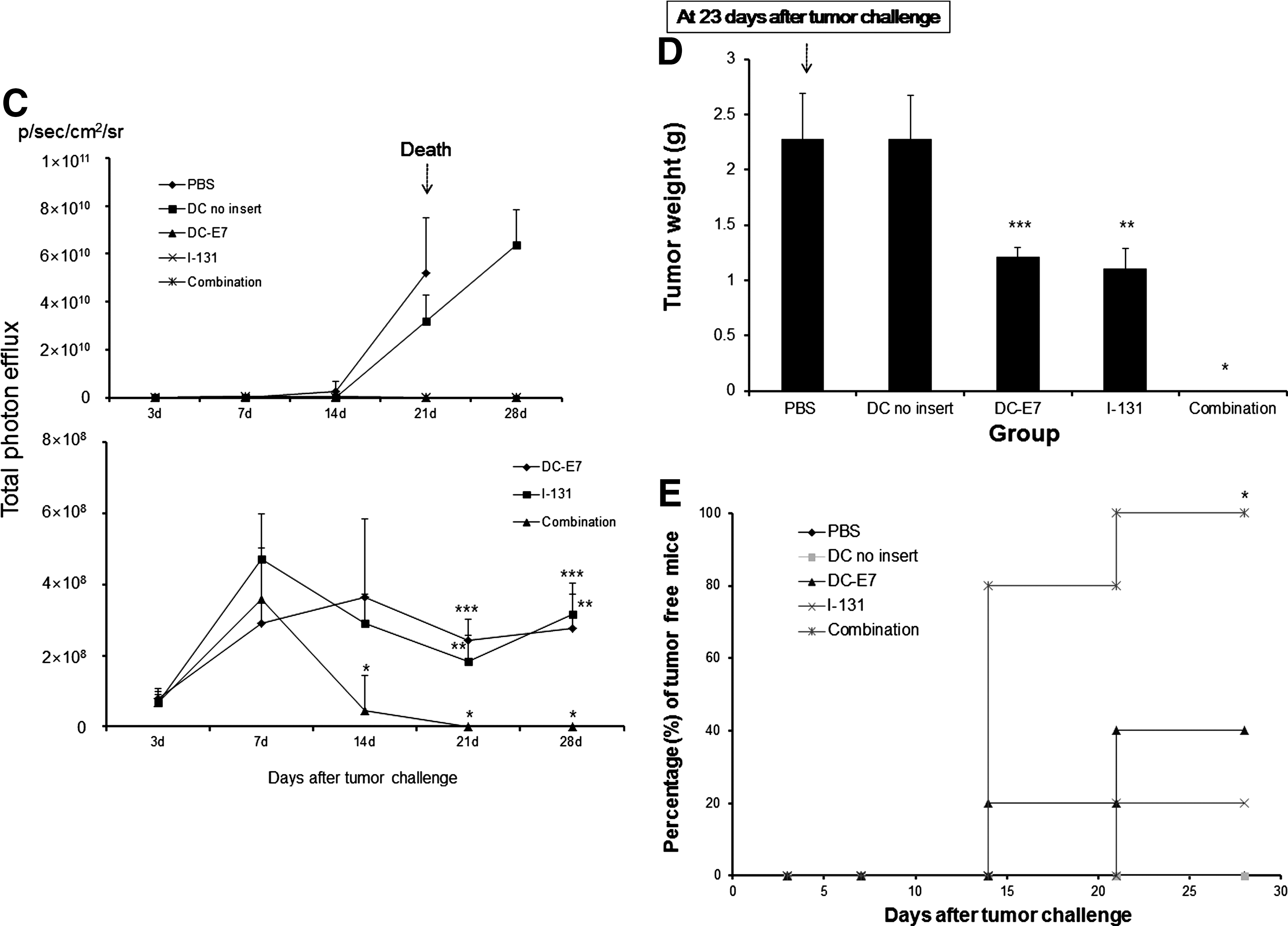
As shown in Figure 5B, aggressive tumor growth was shown in the PBS and DC-no insert groups. Tumor volume of the PBS group reached 10% of the body weight at 23 days, and these mice were not able to survive until 28 days. Thus, BLI of PBS group could not be acquired at 28 days post-tumor challenge. Vaccination with DC-E7 or I-131 treatment alone revealed more effective retardation of tumor growth than both PBS and DC-no insert groups, and a significant difference in tumor growth was shown 21 days after implanatation tumor challenge (Fig. 5B, C; p<0.05, when compared with either the PBS group or DC-no insert group). Two of the 5 mice (40%) in the single DC-E7 vaccination group and 1 of 5 mice (20%) in the I-131 treatment group were determined to be tumor free by palpation and BLI (Fig. 5E). However, an inhibitory effect on tumor growth with either DC-E7 vaccination or I-131 treatment was only monitored 7–21 days after tumor challenge when measured with BLI, and gradual increases of Fluc activity of the tumor were observed until 28 days after implanatation (Fig. 5B, C). Interestingly, a combination of DC-E7 vaccination and I-131 showed a rapid decrease of Fluc activity 14 days after the tumor challenge, resulting in complete inhibition of tumor growth (Fig. 5B, C; p<0.05, when compared with either DC-E7 vaccination or I-131 treatment). Significant difference in tumor growth was observed at 14 days after tumor implantation, and growth inhibition effects of combination therapy were demonstrated seven days earlier than those by either DC-E7 vaccination or I-131 treatment. All mice were tumor free in combination therapy, as determined by both palpation and BLI but not in the other four groups at 28 days (Fig. 5E, p<0.05). As shown in Figure 5D, tumor mass (g) measured after BLI in the DC-E7 or I-131 groups (1.21±0.08 g and 1.10±0.18 g, respectively) was smaller than that for the PBS or DC-no insert groups (2.27±0.41 g and 2.27±0.40 g, respectively, p<0.05, when compared with the PBS or DC-no insert groups). Tumors were not observed by either palpation or BLI in the combination therapy group 28 days after implantation (p<0.05, when compared with the DC-E7 vaccination or I-131 treatment groups). There were no significant decreases in body weight for either single therapy or combination therapy over the 4 weeks of in vivo experimentation (data not shown).
In vivo visualization of tumor growth inhibition effects by combined DC-E7 vaccination and I-131 therapy.
Discussion
Cancer immunotherapy using DCs, irradiated tumor cells, and DNA vaccine-encoding tumor-specific antigen has been widely applied to preclinical cancer models. 4,31,32 Vaccination with DCs provides several advantages over other cancer immunotherapy modalities: (1) DC vaccines have the same effects as the live, attenuated vaccine with regard to their ability to generate a CD8+ T-cell response; and (2) immunotherapy using DCs vaccine was well tolerated with typical adverse effects, such as fever, local injection site reaction, and fatigue. DCs are APCs that recognize, process, and present foreign antigens to T cells in the effector arm of the immune system. 3 Vaccination of DCs loaded with tumor-specific or -associated antigen can control tumor growth by generating a strong cellular immune response in a tumor-bearing host. Although DCs are considered a useful tool for cancer therapy, they are not usually able to induce a robust immune response to tumor antigens, in part because cancer cells are able to suppress and/or escape from normal immune responses. Escape from immune surveillance can result from defects in antigen-processing machinery, the abnormal expression of MHC or costimulatory molecules, and from the production of immunosuppressive factors. 33,34 Thus, we postulated that the weak therapeutic effects of cancer immunotherapy with DC vaccines should be augmented with other therapeutic modalities for further clinical applications.
hNIS radionuclide gene therapy using a therapeutic radioisotope has been introduced as a novel therapeutic method, which induces strong apoptosis and necrosis of irradiated cancer cells. β-rays emitted from I-131 travel 0.2–2.4 mm in tissue, resulting in the death of hNIS-expressing cancer cells via the cross-fire effect. 35 Several research groups have demonstrated that external radiation therapy has the ability to directly kill cancer cells and to generate a radiation-induced immune response; they showed that irradiation to cancer cells induced up-regulation of MHC class I, Fas, ICAM-I, and tumor-specific antigen in various types of cancer, subsequently increasing the killing activity of cytotoxic T cells in vitro and in vivo. 36 –38 For example, Chakraborty et al. 36,37 demonstrated the enhanced phenotype marker and biological effects of irradiated cancer cells on the killing activity of CD8+ T cells.In addition, Garnett et al. reported that when several human cancer cell lines were irradiated with 10 Gy, significant modification of surface markers was found in irradiated human cancer cell lines. 39 We have demonstrated that hNIS radioiodine gene therapy can modulate the phenotype marker (MHC class I and Fas) of cancer cells and increase the killing activity of CTLs against cancer in vitro and in vivo. 23
Based on our recent promising reports, we have presumed that combination of DCs vaccination and hNIS radioiodine gene therapy may further induce strong antitumor effects through both direct killing effects of cancer cells induced by radiation and subsequent generation of increased antitumor immunity. To evaluate the therapeutic potential of the combination therapy, the stable cancer cell line coexpressing E7 antigen (as a tumor-specific antigen), Fluc (as a reporter gene), and hNIS (as a therapeutic gene) gene was first established using both TC-1 cell expressing Fluc and retrovirus coexpressing hNIS and EGFP gene (TC-1/FNG cells). The TC-1 cancer cell proved to be an appropriate target for cancer immunotherapy against E7 antigen-expressing cancer.
24
–26
Kim et al.
29
also visualized the antitumor effect of E7 antigen-specific immunotherapy using Fluc-expressing TC-1 cells.
29
E7-trasnfected DC (DC-E7) was used as a DC vaccine, because it was reported to be an effective DC vaccine for treating the animal model of TC-1 cancer.
24,28
The intramuscular administration route of DC-E7 vaccination was used in this study, because it was reported to generate the most potent E7-specific antitumor immune response.
24
TC-1/FNG cells coexpressing tumor-specific antigen and radionuclide therapeutic genes enable combined DC-based immunotherapy and radionuclide gene therapy within a single-cell element. Further, the use of firefly luciferase gene as a bioluminescent reporter gene in TC-1/FNG allows serial monitoring of the therapeutic effects. Firefly luciferase (Fluc) emits light when it catalyzes its substrate,
Although NIS-based I-131 therapy is noted for the control of certain cancers, there should be other synergic effects of DC vaccination that could be useful in direct radiotherapy. The possible mechanisms responsible for the strong antitumor effects of combination therapy could be suggested as following. First, intramuscular vaccination of DC-E7 may stimulate immature DCs and prime effector T cells, thus prompting the generation of E7-specific immune responses in the tumor-bearing host. Second, I-131 treatment may induce the modification of surface markers including MHC class I, Fas, ICAM-1, and potentially numerous antigenic peptides derived from irradiated cancer cells in the tumor microenvironment. These sequential events may allow E7-antigen loaded DCs to effectively recognize irradiated cancer cells and to cross-present E7 antigenic peptides released from destroyed cancer cells to effector cells. These phenomena may increase the E7-specific antitumor immune response of the combination therapy (DC-E7 vaccination plus I-131) in comparison to single therapy (DC-E7 vaccination or I-131 therapy).
We successfully demonstrated the potent therapeutic effects of combined DC-E7 vaccination and hNIS radioiodine gene therapy in an established animal model of cancer, but some limitations of this study should be addressed before applying the approach in clinics. The hNIS-expressing cancer cell used for radioiodine gene therapy is not a realistic representation of the environment in patients with cancer, and specific delivery of hNIS gene to cancer using vector delivery engineering or a cancer-specific promoter strategy should be studied. Also, as just described, the repertoire of immune response generated by combination therapy should be investigated.
Conclusions
We believe this is the first study to show that a combination of DC-E7 vaccination with hNIS radioiodine gene therapy generates potent antitumor effects (complete tumor growth inhibition) in an established tumor model, and we successfully visualized the therapeutic response to the combination therapy using BLI over time. This combination therapy could be used as a powerful therapeutic strategy for cancer patients with poorly immunogenic cancers or in those patients who do not respond to radiation therapy.
Footnotes
Acknowledgments
This work was supported by the Basic Atomic Energy Research Institute (BAERI, 2010-0017515) and BK21 (2009) and the Korean Ministry of Education, Science and Technology (The Regional Core Research Program/Medical Convergence Technology Development Consortium for Anti-aging and Well-being).
Disclosure Statement
The authors declare that they have no competing interests.