
Abstract
Accumulating evidence has indicated that preconditioning chemotherapy could eliminate the suppressive factors in antitumor immune response, thereby leading to the full release of the efficacy of the subsequent immunotherapy. In this study, a single subtoxic dose (5 mg/kg, intraperitoneally) of cisplatin was chosen as the preconditioning chemotherapy in combination with cytokine-induced killer (CIK) cells (4×106, intravenously) to treat the murine B16 melanoma xenografts. It was found that cisplatin pretreatment could enhance the antitumor activity of CIK cells. To explore the potential mechanisms underlying the efficacy-enhancing effect of cisplatin, the in vivo trafficking and distribution of the infused CIK cells were traced. It was found that cisplatin could augment the homing ability of CIK cells into the tumor, tumor-draining lymph nodes (TDLNs), and spleen tissues. The endogenous effector cells, CD3+ T lymphocytes also had an increased accumulation in the tumor and TDLNs after cisplatin precondition. Moreover, cisplatin could also modulate the percentages of myeloid cells, thus encouraging immune responses by increasing the percentages of dendritic cells and relieving the immunosuppression by preferentially eliminating the myeloid-derived suppressor cells. In conclusion, our findings suggested that cisplatin preconditioning chemotherapy could enhance the antitumor activity of CIK cells in a murine melanoma model, and this efficacy-enhancing effect was attributed to the augmented homing ability of exogenous and endogenous effector cells and the modulation of the myeloid cells.
Introduction
Melanoma is one of the most malignant tumors, and its incidence has been steadily increasing in recent years. 1 Local treatment such as surgery has been applied to treat melanoma in early stages, and the prognosis for these patients is generally favorable. 2 However, due to the high metastatic potential of the tumor, the median overall survival is only 6–9 months. 3
Since melanoma is not sensitive to conventional chemotherapeutic agents, great efforts have been made in the investigation of new drugs. For instance, Ipilimumab (Yervoy), approved by Food and Drug Administration for the treatment of unresectable or metastatic melanoma, 4 is a monoclonal antibody (mAb) that activates the host immune system to fight melanoma by inhibiting the cytotoxic T lymphocyte-associated antigen 4 molecules. Vemurafenib, an inhibitor of the constitutively active mutant of B-raf (V600E), is believed to be a promising antimelanoma drug based on the two positive clinical studies (BRIM2 and BRIM3) in patients with BRAF V600 mutation-positive metastatic melanoma. 5 However, the high price remains the most important obstacle restricting the clinical application of these new agents widely across the world. At present, the primary therapeutic strategy for metastatic melanoma is immunotherapy. Adoptive cell therapy (ACT), as one type of passive immunotherapy, is widely used for patients with advanced melanoma, wherein autologous T lymphocytes are activated and expanded ex vivo and then transfused into patients to exert antitumor activity in vivo. In clinics, this technique has been optimized by the introduction of lymphodepletion or the removal of immunosuppressive factors with chemotherapy or radiotherapy. 6,7 Notably, a significant objective response rate was achieved by the combination of nonmyeloablative chemotherapy and tumor-infiltrating lymphocytes (TIL). 8 Limiting factors in this modality include the severe hematological adverse reaction induced by chemotherapy, TIL-associated autoimmunity, the time-consuming procedure of the TIL culture, and the fact that TIL cannot be established in all patients. 8 Moreover, the adoptive transfer of peripheral T cells transfected with specific TCRs isolated from melanoma-reactive human CTL clones is also an important method for melanoma immunotherapy with time-consuming characters. 9 Therefore, a more safe and practical regimen is needed to benefit the patients with advanced melanoma.
Cytokine-induced killer (CIK) cells, a heterogeneous cell population endowed with potent antitumor activity without autoimmunity, have shown a significant effect on hematological diseases and solid tumors. 10 –13 Cisplatin (DDP), a chemotherapeutic agent whose immunomodulating feature has been recognized for decades, is widely used in tumor treatment. 14 However, patients with melanoma benefit little from chemotherapy, including DDP. To the best of our knowledge, there have been no reports on the combination of CIK and DDP treatment in human melanoma.
In our study, DDP serving as the preconditioning chemotherapy was administered at a subtoxic dose in combination with CIK cells in a murine B16 melanoma model, and the antitumor activity and adverse reactions were observed. Our results show that the DDP pretreatment could enhance the antitumor activity of CIK cells and this combined chemo-immunotherapy was safe and effective in treating B16 melanoma. The underlying mechanisms mediating this DDP-induced immunomodulation may involve the augmentation of the intratumoral infiltration of exogenous CIK cells and endogenous T lymphocytes, increase of the dendritic cell (DC) percentages, and depletion of myeloid-derived suppressor cells (MDSCs) in the B16 melanoma-bearing mice.
Materials and Methods
Tumor cells
B16 melanoma cells were purchased from Shanghai Institute of Biochemistry and Cell Biology and were cultured in complete RPMI 1640 medium (Hyclone) supplemented with 10% fetal calf serum, 100 U/mL penicillin and 100 μg/mL streptomycin. Tumor cells were maintained at 37°C in a humidified atmosphere of 5% CO2.
Animals
SPF C57BL/6 female mice were provided by the Academy of Military Medical Science. The mice used in this experiment were 6–8 weeks of age and housed under controlled conditions with moderate humidity and temperature. The animal experiments were in accordance with guidelines as approved by the Institute Animal Care Committee of Jingling Hospital.
Reagents
DDP was produced in Qilu Medicine Corporation. Murine interleukin-2 (IL-2) and interferon-γ (IFN-γ) were purchased from Peprotech. Functional anti-CD3 antibody used in the culture of CIK cells was provided by eBioscience. PE-Cy5-conjugated anti-CD3, PE-conjugated anti-CD11b, FITC-conjugated anti-CD11c and FITC-conjugated anti-Gr-1 antibodies were purchased from eBioscience. Anti-CD3 antibody used in immunohistochemical analysis was obtained from Abcom. Carboxyfluorescein succinimidyl ester (CFSE) was purchased from Dojindo Laboratories. Methyl thiazolyl tetrazolium (MTT) was purchased from Sigma.
Tumor challenge
Mice were s.c. injected with 106 B16 melanoma cells in their shaved right flank. The tumors were detectable (about 5 mm in diameter) 10 days after tumor challenge. Tumor volumes were monitored to evaluate tumor growth. Tumor volumes were calculated as follows: 0.5×length×width 2 in which length and perpendicular width were measured by calipers.
The generation of CIK cells
The method to educate CIK cells was referred to the procedure previously applied. 15 The single-cell suspensions of spleen tissues from C57BL/6 mice were prepared and enriched for mononuclear cells by density gradient centrifugation using Ficoll-Hypaque (Beijing Chemical Reagents Company). The splenic mononuclear cells were cultured in cRPMI-1640 medium with 1000 U/mL murine IFN-γ added on the first day of culture, 500 U/mL IL-2 and 50 ng/mL functional anti-CD3 antibody added on the second day. Thereafter cRPMI-1640 supplemented with 300 U/mL IL-2 was applied every other day for a total 2-week culture, and the harvested cells were designated CIK cells. Before transfusion into the mice, the CIK cells were incubated with CFSE (5 μM) for 10 minutes in 37°C.
In vitro cytotoxicity assay
B16 cells were pretreated with DDP for 48 hours at a range of concentrations: 0, 0.78, 3.125, and 6.25 μg/mL. Then, the cells with or without pretreatment were collected and plated in 96-well plates at 5×103 cells/well in triplicate. Six (6) hours later, the CIK cells were coincubated with the target cells at 5:1, 10:1, and 20:1 effector-to-target (CIK-to-B16) ratios. After 24 hours of incubation, MTT assay was employed to detect the cytotoxicity of the CIK cells against B16 cells by using an enzyme scale meter. Single effector cells and target cells served as controls. The killing rate (%)=[(OD of experiment−OD of effector cells)/(OD of target cells−OD of medium]×100%.
Experiment design
Mice-bearing B16 melanoma were randomly divided into four groups with 5 mice in each group (Fig. 2A): Group normal saline (NS): treated with NS (0.2 mL, i.p., D0) plus NS (0.1 mL, i.v., D3); Group CIK: treated with NS (0.2 mL, i.p., D0) plus CIK cells (4×106, i.v., D3); Group DDP: treated with DDP (5 mg/kg, i.p., D0) plus NS (0.1 mL, i.v., D3); Group DDP-CIK: DDP (5 mg/kg, i.p., D0) plus CIK cells (4×106, i.v., D3).
Immunohistochemical analysis
Tumor tissues from B16 melanoma-bearing mice were isolated and fixed in 10% formalin on D10. All of the fixed tumor samples were embedded in paraffin for a further immunohistochemical study. Sections (10 μm) were made from all tissues for immunostaining with an appropriate dilution of the anti-CD3 antibody (1:500, rat monoclonal; Abcom). All procedures were performed according to commercial instructions. Images of the sections were acquired on an Olympus BX-60 microscope at 400× magnification for the determination of the density of CD3+ cells.
In vivo trafficking of CIK cells
C57BL/6 mice-bearing B16 melanoma were randomly divided into two groups and received treatment with NS (0.2 mL, i.p.) and DDP (5 mg/kg, i.p.), respectively. Three (3) days after the treatment, the mice were i.v. transferred with 5×106 CIK cells previously incubated with CFSE at a concentration of 5 μM for 10 minutes. The peripheral bloods (PB, erythrocytes removed) were collected, and the liver, spleen (erythrocytes removed), tumor, and tumor-draining lymph node (TDLN) tissues were isolated and prepared to single-cell suspensions at the indicated time points: 2, 24, and 48 hours after the transfusion of CIK cells. The single-cell suspensions of all tissues were stained with PE-Cy5-conjugated anti-CD3 antibody and then analyzed by flow cytometry. CFSE served as a tool to trace the trafficking of CIK cells and to determine the proliferation of CIK cells.
Flow cytometry
B16 melanoma-bearing mice were randomly divided into two groups and received treatment with NS and DDP regimen, respectively. The single-cell suspensions of PB (erythrocytes removed), bone marrow (BM, erythrocytes removed), spleen, and TDLN were prepared 3 days after treatment. The cells were stained with the indicated mAbs and incubated for 30 minutes on ice devoid of light. To determine the percentages of T lymphocytes, DC, and MDSC, single-cell suspensions were gated by plotting forward versus side scatter followed by gating on CD3+, CD11b+CD11c+, and Gr-1+CD11b+ cells, respectively. For suspensions from PB, liver, BM, spleen, and TDLN, 10,000 events were collected, while 100,000 were collected for tumor tissues. The data were analyzed using CellQuest software.
Statistical analysis
Numerical data collected from each experiment were expressed as mean±standard error of the mean. Statistical differences between the groups were compared using ANOVA, and LSD was applied for multiple-means comparisons. Differences were considered significant when p was <0.05. Statistical analysis was conducted using the SPSS software13.0.
Results
DDP pretreatment sensitized B16 cells to CIK-induced lysis in vitro
To acknowledge the immunomodulating effect of DDP on B16 cells, the cytotoxicity of CIK cells against B16 cells was detected in vitro. The DDP dose was selected by MTT assay, and the concentration of DDP that resulted in 10%, 20%, and 40% cell death above the background level within 48 hours was adopted in this study. The killing rate of CIK cells against B16 cells left untreated was 11.90±0.34, 35.01±1.28, and 57.19±0.92 with effector-to-target ratio at 5:1, 10:1, and 20:1, respectively (Fig. 1). Interestingly, the effector cells exerted enhanced cytotoxicity on target cells undergoing DDP pretreatment, with the killing rate of 16.18±0.57, 42±0.96, and 65.77±1.33 after 0.78 μg/mL DDP pretreatment and 20.91±0.99, 51.20±1.15, and 76.48±1.15 after 3.125 μg/mL DDP pretreatment, and 17.46±1.12, 45.16±1.48, and 79.21±0.55 after 6.25 μg/mL DDP pretreatment at 5:1, 10:1, and 20:1 effector-to-target ratio, respectively (Fig. 1). The B16 cells pretreated with DDP were more sensitive to the lyses of CIK cells.
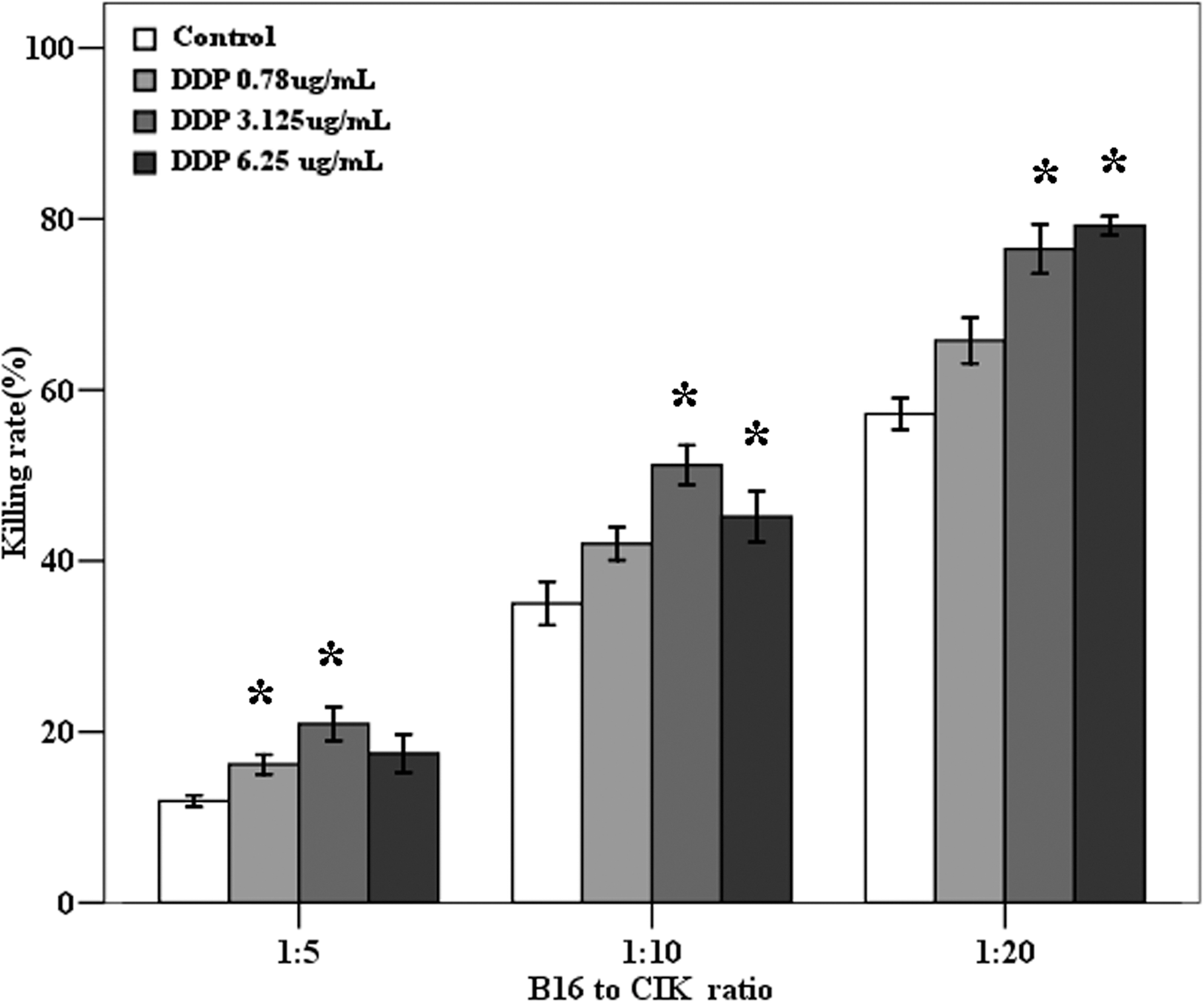
DDP pretreatment sensitized B16 cells to the lyses of cytokine-induced killer (CIK) cells. Five thousand/well B16 cells treated with subtoxic doses of DDP (0.78, 3.125, and 6.25 μg/mL) for 48 hours or without pretreatment (Con) were coincubated with CIK cells at three effector-to-target ratios for 24 hours in 96-well plates. The cytotoxicity of the CIK cells against the B16 cells was determined using methyl thiazolyl tetrazolium (MTT) assay. Columns, mean of triplicated measurements out of three independent experiments; bars, SE. *p<0.05 indicating statistical significance.
Pretreatment with DDP enhanced the antitumor activity of CIK cells
To determine the immunostimulating effect of DDP on adoptive CIK therapy, mice-bearing B16 melanoma were randomly grouped and treated with NS, DDP, CIK cells, and DDP pretreatment plus CIK cells, respectively (Fig. 2A). Tumor volumes were monitored (Fig. 2B) during the whole experiment. In the initial period (D1–D5), there were no obvious differences of the tumor size among the four groups (Fig. 2B, C). However, a combination of DDP and CIK cells induced a significant tumor growth inhibition on D7, and this inhibition was sustained till the end of the study, while single CIK immunotherapy or DDP chemotherapy exerted no effect on tumor growth (Fig. 2B, C).
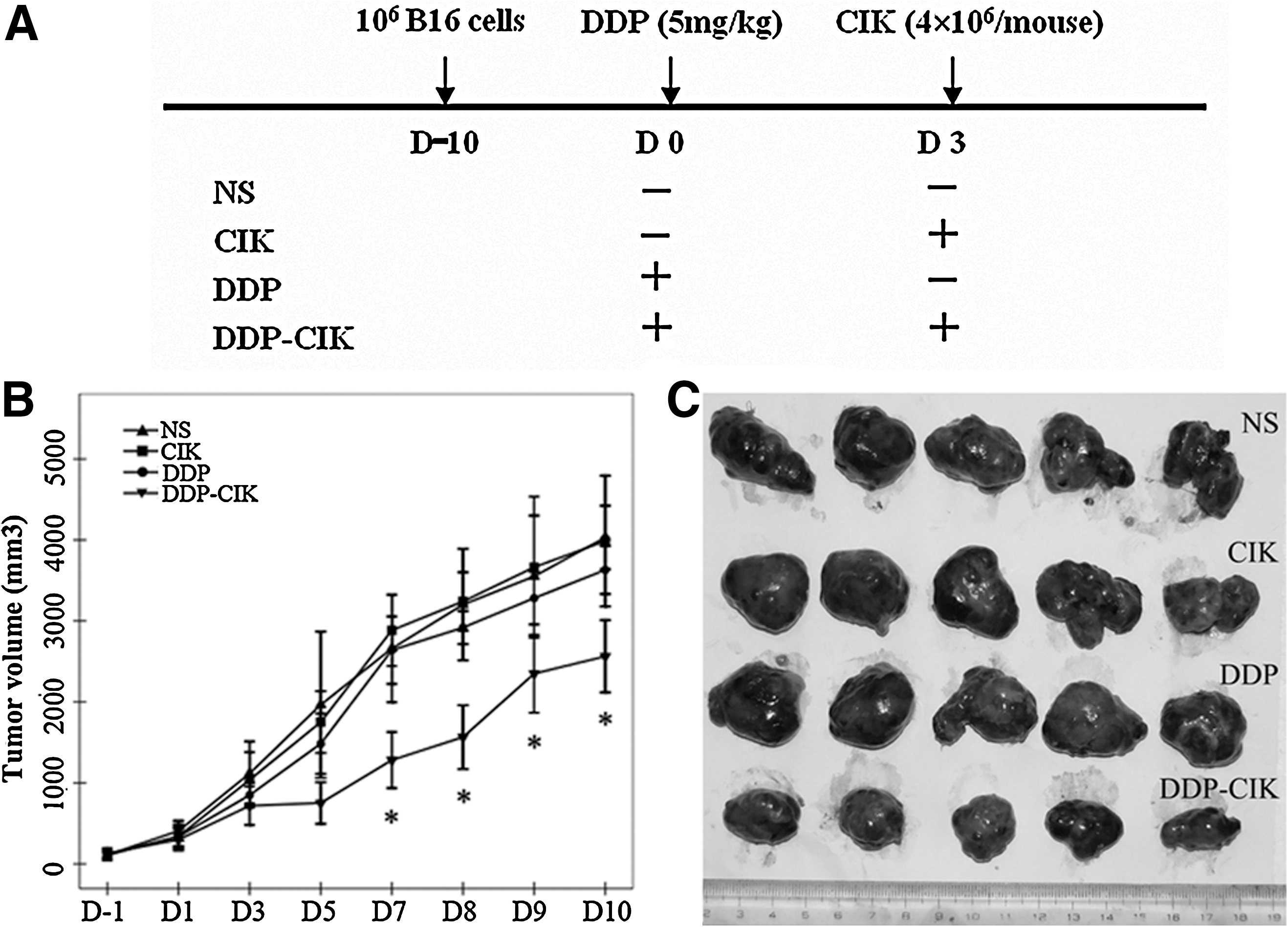
The efficacy-enhancing effect of DDP on CIK cells in the B16 melanoma model. C57BL/6 mice were inoculated with 106 B16 melanoma cells in the right flank. On D0, the tumor-bearing mice were randomly divided into four groups and received treatments according to the treatment schema
DDP pretreatment promoted the infiltration of CD3+ cells into the tumor tissues
Based on the important role of intratumoral effector cells accumulation on the efficacy of immunotherapy, 16,17 immunohistochemistry was applied to evaluate the level of CD3+ cells (exogenous CD3+ CIK cells and endogenous CD3+ T lymphocytes) in tumor tissues isolated on D10. CD3+ cells were nearly unable to infiltrate into the untreated tumor tissues. Single treatment with DDP or CIK cells only induced a mild increase in CD3+ cells accumulation (p>0.05) (Fig. 3A, B). However, tumor tissues after combination therapy were extensively infiltrated with CD3+ cells, with statistical significance compared with other three groups (Fig. 3A, B), which was in accordance with the significant antitumor effect.

The infiltration of CD3+ cells in the tumor microenvironment. C57BL/6 mice-bearing B16 melanoma were randomly divided into four groups and received corresponding treatments (Fig. 2A). The tumor tissues were removed on D10 when the mice were sacrificed and used for immunohistochemical analysis. Sections were prepared and analyzed by CD3 staining (A) and then enumerated the CD3+ cells in ten individual fields (0.04 mm2) at 400× magnification (B). Representative sections are shown from all groups. Scale bar, 25 μm. Columns, means of the numbers of CD3+ cells; bars, SE. *p<0.05, suggesting statistical significance; ns, not significant.
DDP pretreatment augmented the homing ability of CIK cells
To investigate whether pretreatment with DDP could directly influence the in vivo trafficking and distribution of CIK cells, CFSE+ CIK cells were traced by flow cytometry. Compared with the total tissue cells, the numbers of CIK cells were extremely little; it was not feasible to directly determine the percentages of CIK cells over the total tissue cells. Since the majority (about 80%) of CIK cells express the CD3 molecule, CD3+ cells were determined (CD3%) and gated for further determination of the percentages of CFSE+ CIK cells accounting for over-specified CD3+ cells (CFSE%-S) (Fig. 4A,B). The percentages of CFSE+ CIK cells accounting for the total cells were calculated as follows: CFSE0/000-T=CD3%×CFSE%-S.
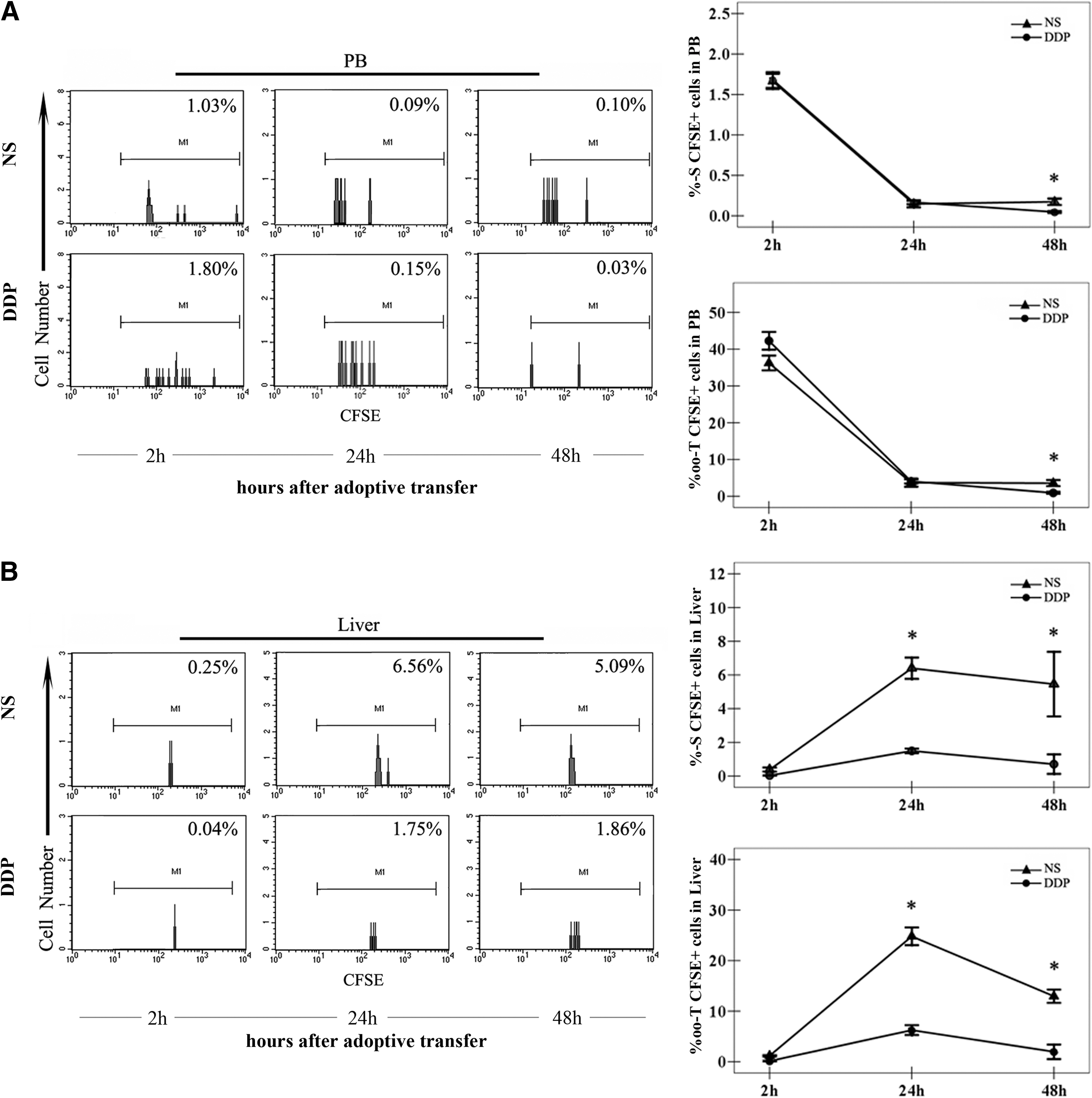
The distribution of CIK cells in peripheral bloods (PB) and liver tissues after transfer. Mice-bearing melanoma (n=3/group) were i.p. injected with NS or DDP (5 mg/kg) and received infusions of 5×106 carboxyfluorescein succinimidyl ester (CFSE+) CIK cells 3 days later. Mice were bled to harvest blood, and the liver tissues were isolated at the indicated time points. According to the protocols, the single-cell suspensions of PB
Two (2) hours after adoptive transfer, the infused CIK cells mainly existed in PB (Fig. 4A) and then, the CIK cells flowed in the circulation and extravasate into various organs. Twenty-four (24) hours after adoptive transfer, CIK cells were detectable in all of the tissues we observed, and the percentages of CIK cells dropped sharply in PB while they peaked in other tissues of interest (Figs. 4 and 5). However, the CIK cells displayed a different distribution pattern in mice treated with NS and DDP (Figs. 4 and 5).
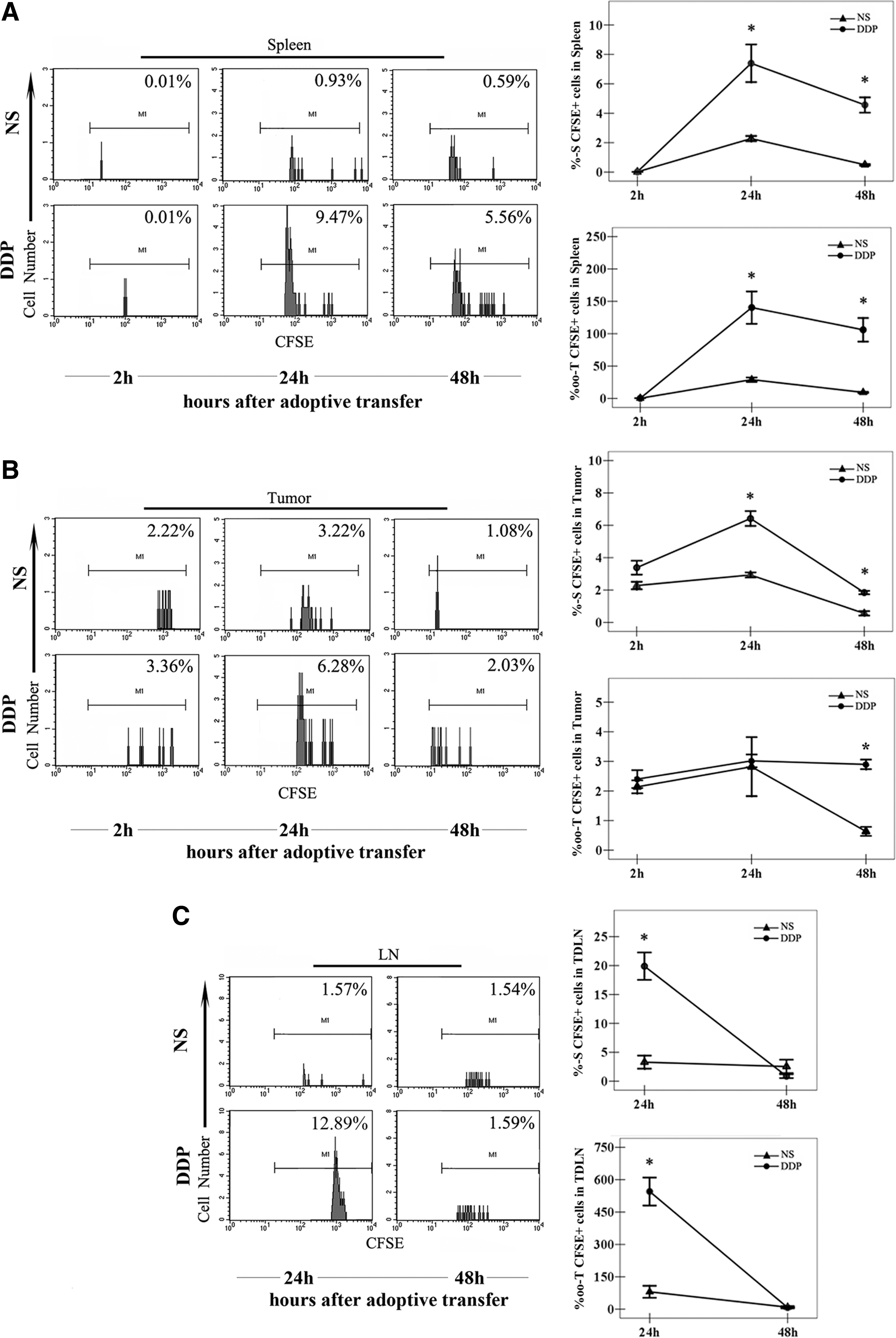
The distribution of the CIK cells in spleen, tumor, and tumor-draining lymph node (TDLN) tissues after transfer. Mice-bearing melanoma (n=3/group) were i.p. injected with NS or DDP (5 mg/kg) and received infusions of 5×106 CFSE+ CIK cells 3 days later. The single-cell suspensions of spleen
The percentages of the CIK cells in PB were not influenced by DDP treatment. CFSE+ CIK cells in both groups flowed in the blood circulation and then declined gradually with a time elapse (Fig. 4A). However, the CIK cells tended to accumulate in the liver of mice treated with NS in comparison with DDP. Although there were no differences in the percentages of CFSE+ CIK cells between the mice treated with DDP and NS 2 hours after transfusion (Fig. 4B), the CIK cells preferentially accumulated in the liver in NS-treated mice at the latter two time points (Fig. 4B).
In spleen tissues, 2 hours after the adoptive transfer, the percentages of the CIK cells in mice treated with DDP appeared comparable to that with NS (Fig. 5A). However, 24 hours after transfusion, there was a surge of the infiltration of CIK cells into the spleens of mice treated with DDP, with a significant difference compared with that with NS (Fig. 5A). This statistical difference lasted even 48 hours after the infusion of the CIK cells (Fig. 5A).
In tumor tissues, the percentages of the CIK cells were comparative between mice treated with DDP and NS in the early stage after the infusion of the CIK cells (Fig. 5B). However, in the late stage after the CIK cell transfer, plenty of CIK cells migrated and infiltrated into the tumor tissue in mice treated with DDP, leading to a significant difference compared with that with NS (Fig. 5B).
In TDLN tissues, the percentages of the CIK cells in the mice treated with DDP were largely higher than that with NS 24 hours after CIK cells transfer, while the levels of the CIK cells were comparable between the two groups 48 hours after the infusion of the CIK cells (Fig. 5C).
DDP pretreatment exerted no effect on the proliferation of CIK cells
Since the chemotherapeutic drugs have been found to enhance the proliferation of effector cells by inducing lymphodepletion, 18 –21 the mean fluorescence index (MFI) of CFSE+ CIK cells was determined to investigate whether DDP featured with this ability. In both groups DDP and NS, the MFI of the CIK cells in all detected tissues attenuated gradually after transfusion (Fig. 6). The MFI of the CIK cells in nonlymph nontarget organs (PB, lung, and liver) were higher than in lymph organs (spleen and TDLN) and target organs (tumor) 48 hours after adoptive transfer (Fig. 6). However, there were no significant differences in CIK cells' MFI between the mice treated with DDP and NS, including lymph organs and nonlymph organs (Fig. 6).
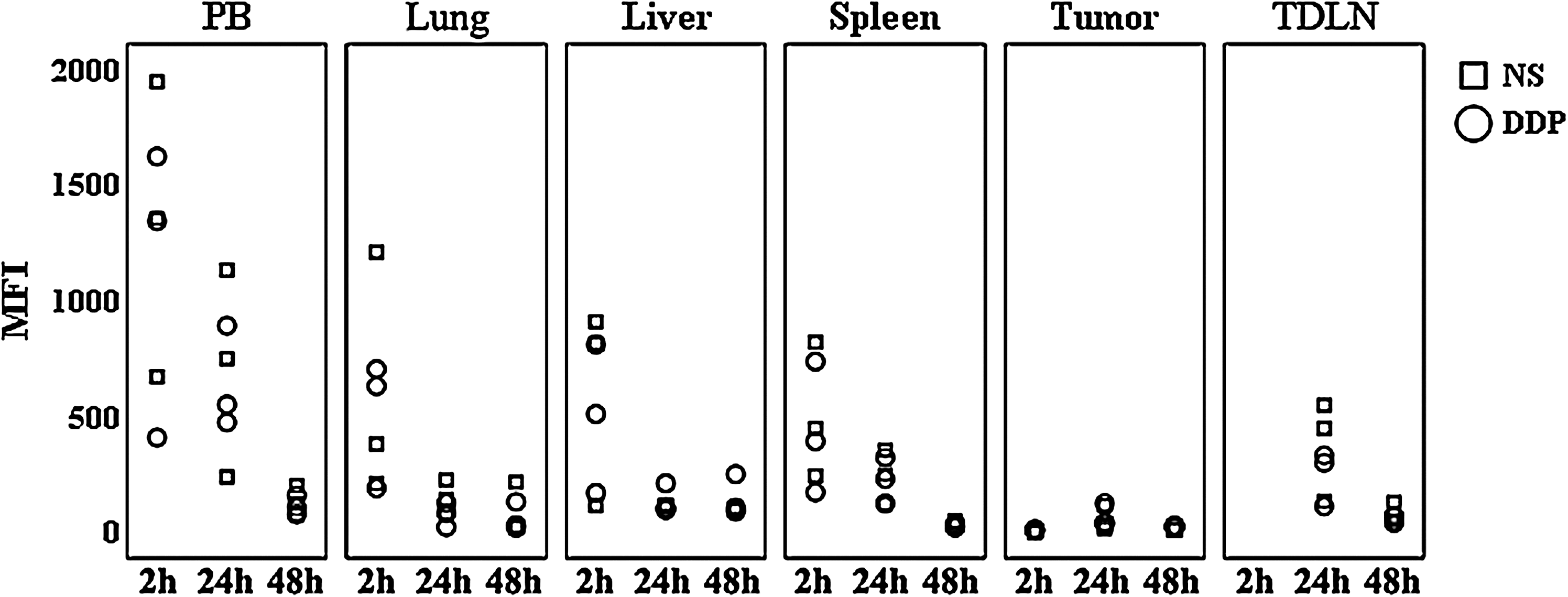
The mean fluorescence index (MFI) of the CIK cells after transfusion. Mice-bearing melanoma (n=3/group) were i.p. injected with NS or DDP (5 mg/kg) and received infusions of 5×106 CFSE+ CIK cells 3 days later. The single-cell suspensions of PB, lung, liver, spleen, tumor, and TDLN tissues were prepared to analyze the MFI of CFSE+ CIK cells by flow cytometry at the indicated time points. Points, value of MFI of CFSE+ CIK cells.
DDP treatment induced a transient increase of CD3+ T lymphocytes in the tumor and TDLN
To investigate the effect of DDP treatment exerted on the endogenous T lymphocytes, the percentages of CD3+ T cells in PB, tumor, and TDLN tissues from B16 melanoma-bearing mice were monitored at indicated time points after DDP treatment. In PB, the percentages of CD3+ T lymphocytes after DDP treatment were comparable with the levels after NS at the most time points (Fig. 7A). Though DDP induced a slight decrease in the percentages of CD3+ T lymphocytes on D3 (3 days after DDP treatment) and a subsequent mild rebound on D6, there was no significance between the mice receiving DDP and those receiving NS (Fig. 7A). In tumor tissues, during the whole observation, the level of CD3+ T lymphocytes after DDP were higher than NS; however, the significant difference only appeared on D6 (Fig. 7B). The CD3+ T cells in TDLN seemed to be more sensitive to DDP treatment compared with tumor tissues. Without DDP treatment, the percentages of CD3+ T lymphocytes in TDLN gradually decreased, reversely related with the increasing tumor burden (Fig. 7C). Nevertheless, owing to the DDP treatment, the levels of CD3+ T cells in TDLN underwent a dramatic course, declining on D1, mounting to the peak on D3 and then dropping to the baseline on D6 (Fig. 7C).

The dynamic changes of endogenous CD3+ T lymphocytes after DDP treatment. B16 melanoma-bearing C57BL/6 mice were treated with NS (0.2 mL, i.p., D0) or DDP (5 mg/kg, i.p., D0), respectively. Mice were sacrificed to harvest PB
DDP treatment increased the percentages of DC in BM and PB
In spleen and TDLN tissues, the percentages of CD11b+CD11c+ DC in mice treated with DDP did not manifest significant differences as opposed to that from NS-treated mice (Fig. 8A, B). In contrast, the percentages of DC in PB from mice treated with DDP were markedly increased in comparison to that from mice treated with NS, demonstrating that a precondition with DDP could induce an increase of DC percentages in PB (Fig. 8A, B). Moreover, DDP treatment also increased the percentages of DC in BM tissues, which suggested that DDP pretreatment could mobilize the BM to produce more DC, possibly contributing to the increase of DC percentages in TDLN tissues.
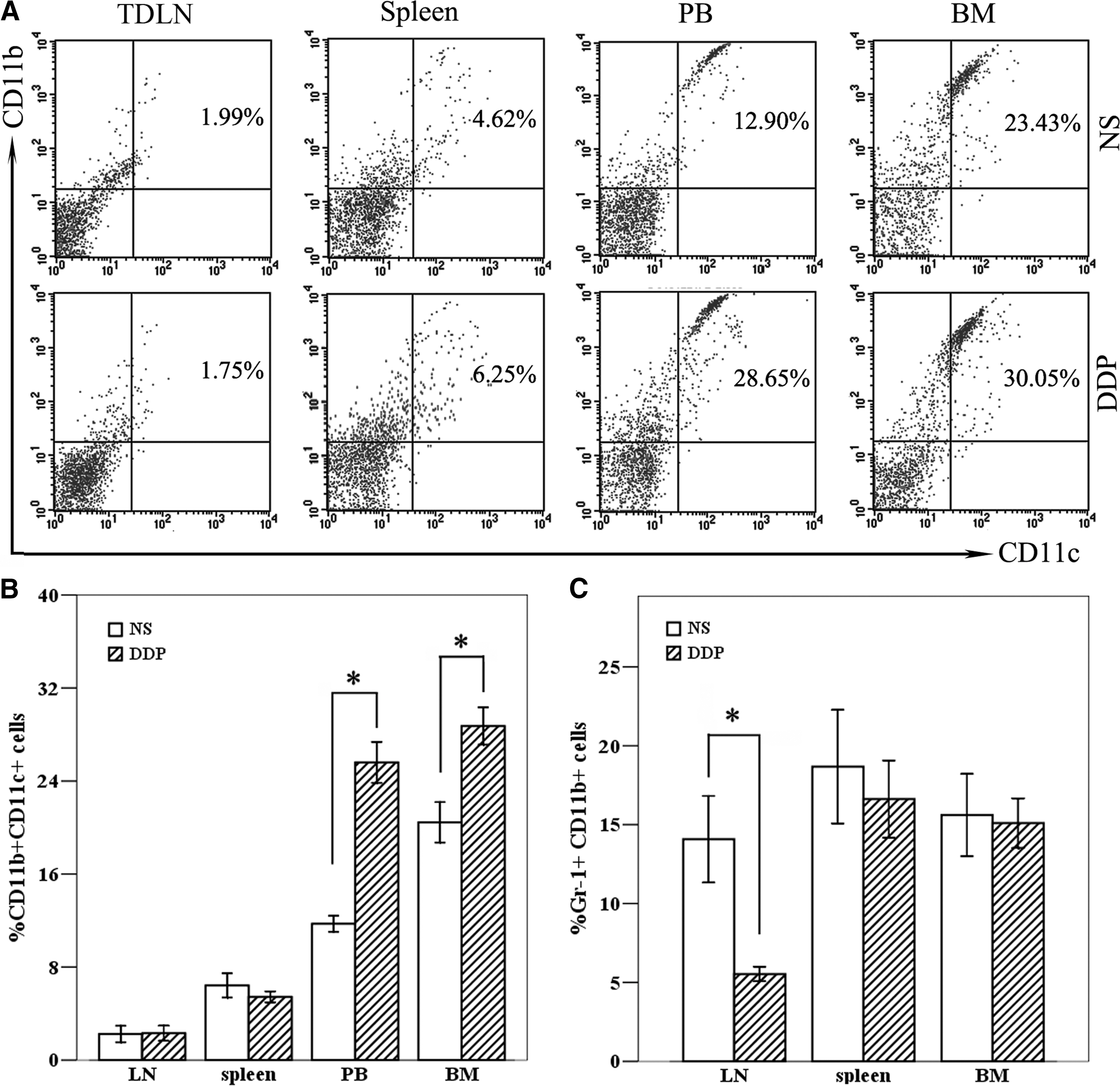
The percentages of dendritic cells (DC) and myeloid-derived suppressor cell (MDSC) after DDP treatment. B16 melanoma-bearing mice were treated with NS (0.2 mL, i.p., D0) and DDP (5 mg/kg, i.p., D0), respectively. Three (3) days after treatment, the single-cell suspensions of PB, bone marrow (BM), spleen, and TDLN tissues were prepared, stained with corresponding mAbs, and the CD11b+CD11c+ cells
DDP treatment depleted the Gr-1+CD11b+ MDSC in TDLN tissues
In this part, we concentrated on the effect of DDP treatment on the percentages of Gr-1+CD11b+ MDSC, a primary immunosuppressive cell population inhibiting the antitumor activity of immunotherapy. 22 The percentages of MDSC in BM, spleen, and TDLN tissues were detected 3 days after DDP treatment (Fig. 8C). DDP treatment seemed to lack the ability to induce the decrease of MDSC percentages in BM and spleen tissues, and a significant depletion of MDSC was only observed in TDLN tissues (Fig. 8C).
Discussion
Recent years, more and more investigators were engaged in the research of immunotherapy, which brings hope for the patients with malignancies. However, due to the immunosuppressions existing in these patients, the effect of applied immunotherapy seems gloomy. 23 Fortunately, great quantities of studies have suggested that some chemotherapeutic agents, when serving as an immunoregulating preconditioning chemotherapy, possess the ability to improve the effect of immunotherapy, including ACT, vaccine, and antibodies. 24 –26 It is, therefore, reasonable that the allied antitumor effect of combination modality of preconditioning chemotherapy and immunotherapy would be exciting and remarkable.
CIK cells, which could be acquired through a simple procedure, have revealed robust in vitro cytotoxicity against a variety of endogenous and exogenous tumor cells. 27 However, in our murine B16 melanoma model, CIK transfer alone failed to induce significant tumor growth inhibition. This inefficiency may be attributed to the immunosuppressive factors existing in these tumor-bearing mice, which hindered both the homing and cytolysis stages of effector cells in vivo. 28 To remove these inhibitory elements and enhance the antitumor activity of the CIK cells, DDP serving as the preconditioning chemotherapy was incorporated into the CIK cell transfer. It was found that preconditioning with a single dose of DDP could significantly enhance the efficacy of CIK cells, and this alliance induced an obvious tumor inhibition against B16 melanoma, intriguing us to make further effort on the mechanisms underlying the efficacy-enhancing effect of DDP.
Actually, a number of studies have shown that the conventional anticancer agent DDP, in addition to its direct cytotoxicity on tumor cells, possesses the ability to modulate antitumor immune responses. It could increase the susceptibility of tumor cells to the lysis by lymphocytes through up-regulating the expression of the NKG2D ligand (NKG2DL) or Fas on tumor cells. 29,30 Our previous study has also demonatrated that DDP could deplete the splenic and intratumoral Treg cells to guarantee the success of CIK therapy in a murine CT-26 colon adenocarcinoma model. 31 However, to the best of our knowledge, there have been no studies characterizing the potential effects of DDP on the homing ability of effector cells.
In this study, DDP pretreatment significantly increased the number of CD3+ cells infiltrating into the tumor tissues on D10, 10 days after DDP administration. As is known, the intratumoral infiltration of lymphocytes is a prerequisite for triggering an effective antitumor immune response. 15,16 Nevertheless, the CD3+ immune cells infiltrating into the tumor tissues and exerting antitumor activity comprised exogenous CD3+ CIK cells (approximately accounting for 80% of total CIK cells) and endogenous CD3+ T lymphocytes. The dynamic changes of these two effector cells after DDP treatment were analyzed in vivo.
Our data also showed that the trafficking and infiltration of the infused CIK cells were regulated by DDP pretreatment. The CIK cells in mice treated with NS preferred to stay in PB and liver tissues, while the CIK cells in mice preconditioned with DDP infiltrated more extensively into the lymph organs (spleen and TDLN) and target organ (tumor), where the CIK cells could more frequently encounter and successfully kill tumor cells rather than get lost in the peripheral circulation. Therefore, it was probable that the augmented homing behavior of the CIK cells after DDP pretreatment was sophisticatedly programmed. Given that the homing of lymphocytes was driven by a variety of chemokines, 32 we proposed that the in vivo trafficking and extravasation of CIK cells may be also mediated by their specific chemokines. In tumor-bearing mice, these chemokines in the tumor microenvironment and lymph organs were set at a fixed level, which may not be adequate to induce large-scale homing of CIK cells. However, the DDP pretreatment could reset this level through modulating the local and systemic immune environment and, thus, lead to the extensive homing of CIK cells. This hypothesis was supported by the research of Bracci. 24
In addition to the augmented homing of CIK cells, the proliferation of the CIK cells may also contribute to their increased percentages. Previous studies have shown that a high-dose chemotherapeutic agent could induce lymphodepletion and then initiate the homeostatic proliferation of the transferred effector cells under the cytokine storm. 18 Therefore, the MFI of CFSE+ CIK cells were determined to evaluate the proliferation of the CIK cells. Our data showed that the MFI of the CIK cells were comparable between mice treated with DDP and NS, suggesting that the single dose of DDP used in this study exerted no effect on the proliferation of CIK cells and that the increased percentages were only attributed to the augmented homing ability of CIK cells. However, the MFI of the CIK cells in nonlymph, nontarget organs were higher than in lymph and target organs, indicating a bigger chance to activate and proliferate in latter organs.
Noteworthy, the infused CIK cells seemed to persist transiently in vivo without the maintenance of high-level cytokine. Forty-eight (48) hours after adoptive transfer, the percentages of the CIK cells rapidly declined to a low level. Especially in tumor tissues, the CIK cells accounted for approximately only 2% of CD3+ cells, even less than the level 2 hours after adoptive transfusion. Therefore, to pursuit effective and sustained antitumor activity, repeated infusion of CIK cells and additional administration of cytokines were needed to induce the eradication of tumor cells.
The kinetics of endogenous CD3+ T lymphocytes after DDP was also monitored. It was found that DDP treatment induced a transient increase of CD3+ T lymphocytes in tumor and TDLN tissues. In our previous study, it was demonstrated that the endogenous CD3+ T lymphocytes were essential for the efficacy-enhancing effect of DDP, 33 possibly acting as assistant cells co-operating with CIK cells to successfully lyse tumor cells. In tumor tissues, the significant difference of endogenous CD3+ T lymphocytes between mice treated with DDP and those teated with NS appeared on D6, which was later than that in TDLN. The CIK cells transferred on D6 were supposed to acquire the optimal assistance of endogenous CD3+ T lymphocytes and, thus, possess maximal antitumor activity. However, the CIK cells in this study were transfused on D3, when the endogenous CD3+ T lymphocytes in TDLN reached the peak. It was meaningful to perform further studies to compare the efficacy-enhancing effect of DDP preconditioned 3 and 6 days before the CIK cells.
To thoroughly illuminate the immunoregulating effect of DDP, the changes of DC and MDSC after DDP pretreatment were also observed in our study. As is known, both DC and MDSC were derived from myeloid precursor cells. Nevertheless, they differentiated differently in the later stage and functioned distinctly in the battle with tumor cells. DC was the primary antigen-presenting cell in the immune system and has been reported as being able to enhance the antitumor activity and proliferation potential of the CIK cells through depleting the CD4+CD25hi cells in heterogeneous CIK populations, 19 thus usually used to be cocultured with CIK cells in vitro (named DC-CIK) to enhance the specificity and capacity of CIK cells' cytotoxicity, and this culture system was applied in clinics. 33 MDSC were induced by the tumor-derived factors by blocking cell differentiation in the myeloid lineage, 34 with potent immunosuppressive activity to inhibit both adaptive and innate immunity through multiple mechanisms. 21 Interestingly, MDSC would differentiate into mature DC under normal differentiation conditions. 35 Overall, the increase in DC level and the decrease in MDSC level may benefit the antitumor activity of effector cells. Recent studies have shown that some chemotherapeutic drugs could modulate the level of these two immune cells. 36 –38 CTX was found to induce a rapid activation of DC during the lymphopenic phase and a later expansion of DC during the recovery phase. 36 Gemcitabine and 5-fluorouracil were reported to selectively deplete MDSC, and the elimination of MDSC promoted T-cell-dependent antitumor responses in vivo. 37,38 In accordance, DDP in this study was also found to possess the ability to regulate the DC and MDSC levels with tissue specificity. The changes in DC were mainly manifested in a systemic immune environment, while the changes in MDSC were restricted in TDLN, a relatively local immune environment and more closely related with tumor tissues. This phenomenon may be due to the different sensitivity of these two cells to DDP and the distinct role they play in the immune system. However, the modulation of myeloid lineage induced by DDP pretreatment may contribute to the enhanced antitumor activity of the CIK cells.
Conclusions
Our study suggested that the DDP pretreatment could enhance the antitumor activity of the CIK cells, and this combined chemo-immunotherapy was safe and effective in treating B16 melanoma. The underlying mechanisms mediating this DDP-induced immunomodulation may involve the augmentation of the intratumoral infiltration of exogenous CIK cells and endogenous T lymphocytes, an increase in the DC percentages, and the depletion of MDSC in the mice-bearing B16 melanoma.
Footnotes
Acknowledgments
This study was supported by a grant from the National Natural Science Foundation of China (NSFC 30872979). This article was reviewed by Dr. Jun Yi and Dr. Bing Feng.
Disclosure Statement
The authors report no conflicts of interest. The authors are responsible for the content and writing of this article.