
Abstract
Noxa is an important proapoptotic protein in the intrinsic pathway of cell apoptosis. Experiments were carried out to investigate whether Noxa could, therefore, enhance the cytotoxic effect of gemcitabine in human ovarian cancer cell lines (A2780 and COC1). In this study, the combined treatment of Noxa and gemcitabine, in vitro, significantly inhibited the proliferation of A2780 and COC1 cells, as verified by MTT assay, Hoechst staining, and flow cytometric analysis. Moreover, the combination of Noxa and gemcitabine inhibited tumor growth and prolonged the survival of nude mice in vivo. The combined treatment also inhibited the growth of tumor xenografts through the inhibition of proliferation and the induction of apoptosis, as observed in immunohistochemical anti-PCNA staining and TdT-mediated dUTP-biotin nick-end labeling (TUNEL) assay. Our data suggest that Noxa exhibited potent proapoptotic activity against human ovarian cancer cells, and the combination of Noxa and gemcitabine showed a more significant cytotoxic effect against ovarian cancer cells in comparison with either of these agents alone. To our knowledge, we have provided the first evidence that Noxa can enhance therapeutic responses of ovarian cancer cells to gemcitabine, and that it could be potentially useful as a chemosensitizer in ovarian cancer therapy.
Introduction
Ovarian cancer is the most deadly gynecologic cancer and the fifth leading cause of cancer death in women. 1 Since its symptoms are not specific, it is difficult to detect ovarian cancer in the early stages. 2 Therefore, most patients with ovarian cancer are usually diagnosed in stage III or stage IV of the disease. For women with advanced-stage ovarian cancer, the 5-year survival rate is very poor. 3 Traditional treatment of advanced ovarian cancer is based on surgical resection, followed by chemotherapy. 4 Despite aggressive treatments, patients with ovarian cancer have a high likelihood of recurrence. Moreover, the therapeutic effect is significantly limited by the frequent development of drug resistance in recurrent diseases, which is a major obstacle to successful chemotherapy. 5 Therefore, new strategies are needed to sensitize cancer cells to chemotherapeutic drugs.
Gemcitabine (2V, 2V-difluoro-2V-deoxy-cytidine), a deoxycytidine analog, is structurally related to deoxycytidine, with two fluorine substitutes for the two hydrogen atoms in the 2′ position of the deoxyribose sugar. 6 Gemcitabine enters the cell and is transformed to its difluorodeoxycytidine triphosphate, which has the ability to be incorporated into DNA, where it then terminates DNA-chain elongation. Due to its prolonged retention time in tumor cells, it has high activity against a variety of solid tumors, including ovarian cancer, head and neck cancer, bladder cancer, pancreatic cancer, and non-small-cell lung cancer. 7 –11
Apoptosis is a form of programmed cell death that detects and purges developmentally excess, mutated, and damaged cells. 12 It can be initiated by a variety of stimuli through two distinct pathways. The extrinsic death pathway is initiated by the binding of death ligands to surface receptors and the subsequent formation of the death-inducing signaling complex, which consists of receptor molecules, the adaptor FADD, and CASP8/caspase-8. 13 The intrinsic or mitochondrial apoptotic pathway is regulated mainly by protein–protein interactions of the Bcl-2 protein family members. 14
Noxa is a BH3-only protein that belongs to the Bcl-2 family. It plays an important role in the intrinsic pathway of cell apoptosis. Its expression is transcriptionally regulated through p53, HIF-1 a, and NF-kB by cellular stresses such as DNA damage, hypoxia, and ischemic injury. 15 Induction of the Noxa gene can trigger the activation of proapoptotic BAK and BAX. 16 On activation, BAX and/or BAK oligomerize in the mitochondrial outer membrane, thereby making the mitochondrial outer membrane permeable by generating pores through which factors such as cytochrome C are released to cytosol. 17 In the presence of ATP or dATP, Cyto-c binds to APAF-1, leading to activation of the initiator caspase-9, which, in turn, activates executioner caspases. 16 On the other hand, Noxa also selectively neutralizes Mcl-1, which is an important antiapoptotic protein. 18 Recently, Noxa has been shown to induce apoptosis in some cancer cells, such as the HeLa human epithelial cervical variety. 19 Furthermore, a recent report by Suzuki et al. showed the therapeutic potential of Noxa in treating human breast cancer. However, the role of Noxa in the therapeutic responses of ovarian cancer cells to anticancer drugs such as gemcitabine remains unclear. 20
On the basis of the proapoptotic activities of gemcitabine and the observation that Noxa could induce significant apoptosis when overexpressed in some tumor cells, the present study was designed to investigate whether Noxa could augment the apoptosis of tumor cells induced by gemcitabine in vitro, and whether the induction of apoptosis by Noxa could work co-operatively with gemcitabine to enhance the latter's antitumor efficacy in vivo.
Materials and Methods
Plasmid purification
Both pcDNA3.1-Noxa (pcDNA vector encoding human Noxa gene) expression plasmid (kindly provided by Dr. David Huang) and pcDNA3.1 vector (used as a control) were purified with Endo-Free kits (Qiagen). The purified plasmids were mixed with liposome to form DNA-liposome complex and were then used for subsequent animal experiments.
Cell culture and treatments
Human ovarian cancer A2780 cells and COC1 cells were obtained from the American Type Culture Collection and were grown in DMEM (Gibco) containing 10% fetal bovine serum, 50 units/mL of penicillin, and 50 μg/mL of streptomycin at 37°C with 5% CO2 in a humidified incubator. About 2×105/2×103 cells were grown in each well of 6/96-well plates and incubated overnight to 70% confluence. Transfection was performed using Lipofectamine™ 2000 according to the manufacturer's instructions.
The A2780 cells and the COC1 cells were classified into the following five groups: Control—the cells were left untreated; pcDNA3.1 alone—the cells were transfected with the pcDNA3.1 plasmid; Noxa alone—the cells were transfected with the pcDNA3.1-Noxa plasmid; gemcitabine alone—the cells were treated with 0.3 μg/ml gemcitabine; and Noxa plus gemcitabine (combination)—the cells were transfected with the pcDNA3.1-Noxa plasmid and then treated with 0.3 μg/ml gemcitabine. Forty-eight (48) hours after transfection, the cells were harvested for subsequent experiments.
Detection of Noxa overexpression
To determine the overexpression of Noxa, the total protein of the six-well plates was extracted with 1% Chaps buffer (5 mM MgCl2, 137 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Chaps, 20 mM Tris-HCl [pH 7.5], and protease inhibitors [Complete, Roche]). Western blot analysis was performed using an anti-Noxa antibody (Santa Cruz), and β-actin expression served as a loading control.
Hoechst staining
The cells treated as just described were fixed in 4% paraformaldehyde for 20 minutes. After rinsing twice in phosphate-buffered saline (PBS), these cells were stained with 1 mg/ml Hoechst 33258 for 10 minutes in a dark chamber and then washed with PBS. Finally, apoptosis was visualized with a ZEISS fluorescence microscope (Jena, Inc.).
MTT assay
After various designated treatments, cell proliferation was measured by MTT assay. The data represented the average of three wells, and each assay was replicated thrice. Media-only-treated cells served as the indicator of 100% cell viability.
Flow cytometric analysis
Cells in the six-well plates were harvested, and a quantitative evaluation of cellular apoptosis was carried out by flow cytometric analysis using the propidium iodide (PI) staining method. The cells were stained with PI and were analyzed using a flow cytometer (ESP Elite; Coulter) to measure the percentage of sub-G1 cells.
Western blot analysis of caspase-3 and caspase-9
After being treated with Noxa or/and gemcitabine, A2780 cells in the six-well plates were harvested, and western blot analysis was used to characterize the expression of caspase-3 and caspase-9. Anti-caspase-3, anticleaved caspase-3, anti-caspase-9, and its cleaved form (Cell Signaling Technology) were used, and β-actin expression served as a loading control.
Xenograft mouse models and treatments
Six- to 8-week-old female nude mice were obtained from the Chinese Academy of Medical Science. Each mouse was injected s.c. with A2780 cells or COC1 cells (5×106 cells) in the right flank. Mice were treated on day 8 after inoculation. The mice were randomly divided into the following five groups (6 mice per group): (1) Control: group treated with 100 μL PBS, intravenous (i.v.) by tail vein; (2) pc3.1: group treated with 10 μg pcDNA3.1 plasmid/30 μg liposome complexes in 100 μL PBS; (3) Noxa: group treated with 10 μg pcDNA3.1-Noxa plasmid/30 μg liposome complexes in 100 μL PBS; (4) Gemcitabine: group treated with 100 mg/kg gemcitabine, intraperitoneal (i.p.) weekly for 4 weeks; (5) Noxa+Gem: (3) and (4) combined. The mice were treated with a DNA-liposome complex by i.v. administration via the tail vein twice a week, and received gemcitabine by the i.p. route once a week for 4 weeks. Tumor volume was calculated as length×width 2 ×0.52. 21 At the end of the experiment, the mice were sacrificed, and the tumor tissues were collected for subsequent immunohistochemical analysis and TdT-mediated dUTP-biotin nick-end labeling (TUNEL) experiments. This work was approved by the institute's Animal Care and Use Committee.
Histopathological examinations
The tumors were fixed in 4% formaldehyde, then embedded in paraffin, and cut into 3–5 μm sections. Specimens were stained with hematoxylin and eosin (H&E). The anti-PCNA antibody (Cell Signaling Technology) was used for the immunohistochemistry. TUNEL staining was also performed, using the DeadEnd Fluorometric TUNEL System (Promega). The apoptotic index was calculated by determining the percentage of positively stained cells for all of the nuclei in five different fields. Slides of the apoptosis studies were quantified in a blind manner by two different reviewers.
To observe the potential side-effects, tissues of the heart, kidney, spleen, lungs, and liver were embedded in paraffin. Sections were stained with H&E and examined under a microscope.
Data analysis
SPSS 17.0 was used for statistical analysis. All the values were expressed as means±standard deviation. Data were analyzed by one-way analysis of variance, and then, differences among the means were analyzed using the Tukey–Kramer multiple comparison test. Survival curves were constructed according to the Kaplan–Meier method. 22 Statistical significance was determined by the log-rank test. 23 Differences were considered significant if p<0.05.
Results
Overexpression of Noxa in A2780 and COC1 cells
Western blot analysis was performed to test the elevated expression of Noxa. As shown in Figure 1, much more Noxa protein was expressed in the cells transfected with pcDNA-Noxa than in the cells of the two control groups. These results demonstrated the overexpression of the human Noxa gene in the transfected cells.
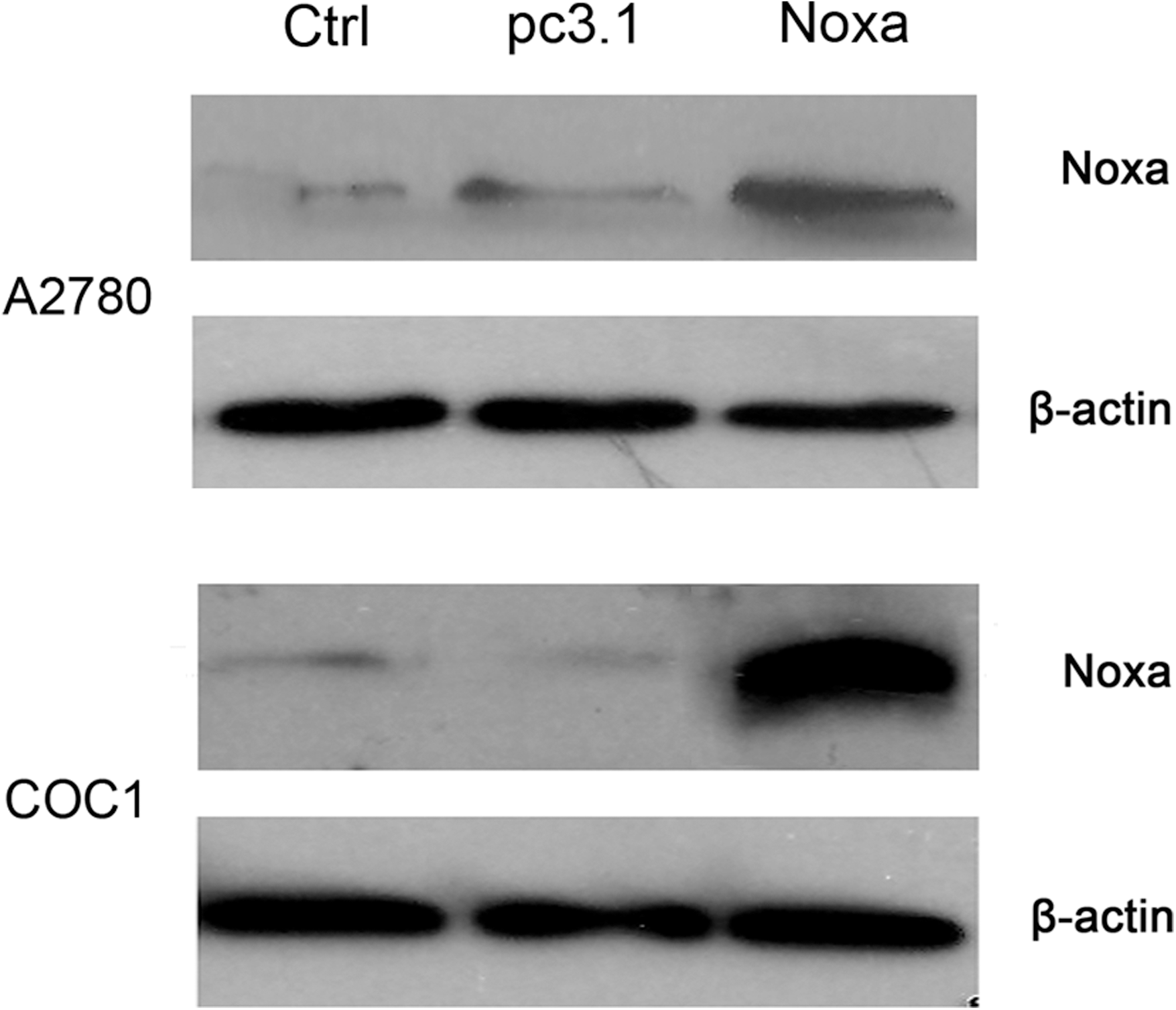
Overexpression of Noxa in ovarian cancer cells. Forty-eight (48) hours after transfection with pcDNA3.1-Noxa, A2780 and COC1 cells were harvested for western blot analysis. A2780 cells and COC1 cells untreated (Ctrl), transfected with pcDNA3.1 (pc3.1), and transfected with pcDNA3.1-Noxa (Noxa) are shown. β-actin expression was used as a loading control.
Noxa enhances the antiproliferative effect of gemcitabine in vitro
The results of the MTT assay showed the cell viability of A2780 or COC1 cells after the treatments. As shown in Figure 2B, the viability of cells decreased significantly in the gemcitabine and Noxa groups in comparison with the two control groups (p<0.05). The combined treatment using Noxa and gemcitabine together significantly decreased the cell viability compared with the single gemcitabine and Noxa groups (p<0.05).
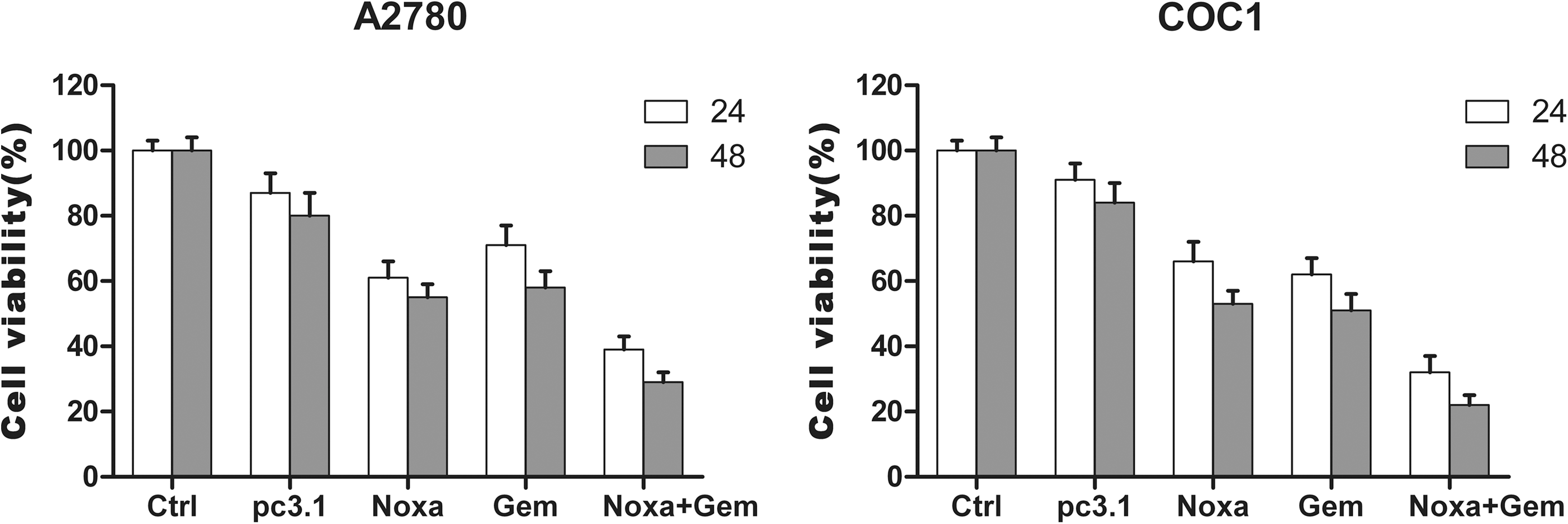
Viability of A2780 and COC1 cells treated with Noxa, gemcitabine, or a combination of both. MTT assay was carried out as described in the Materials and Methods section. The cell viability of the combined group (Noxa and gemcitabine) is significantly decreased in comparison with the group using gemcitabine alone. The statistics showed a significant difference between the two groups (p<0.05). Data represent means±standard deviation (SD) of three wells and experiments performed in triplicate. The untreated cells were set as 100% cell viability in each experiment.
Morphological changes of apoptotic cells, which manifested as more floating and rounded cells, can be seen under a phase-contrast microscope. More floating and rounded A2780 cells were found in the group treated with Noxa and gemcitabine (Fig. 3A). The typical chromatin condensation and fragments of the cell nucleus in the apoptotic cells induced by the treatments were shown by Hoechst staining. More pyknotic and fragmented nuclei of A2780 cells can be seen in the group treated with Noxa and gemcitabine in comparison with the groups treated with either agent alone (Fig. 3B). Similar results were observed in the treated COC1 cells (data not shown).
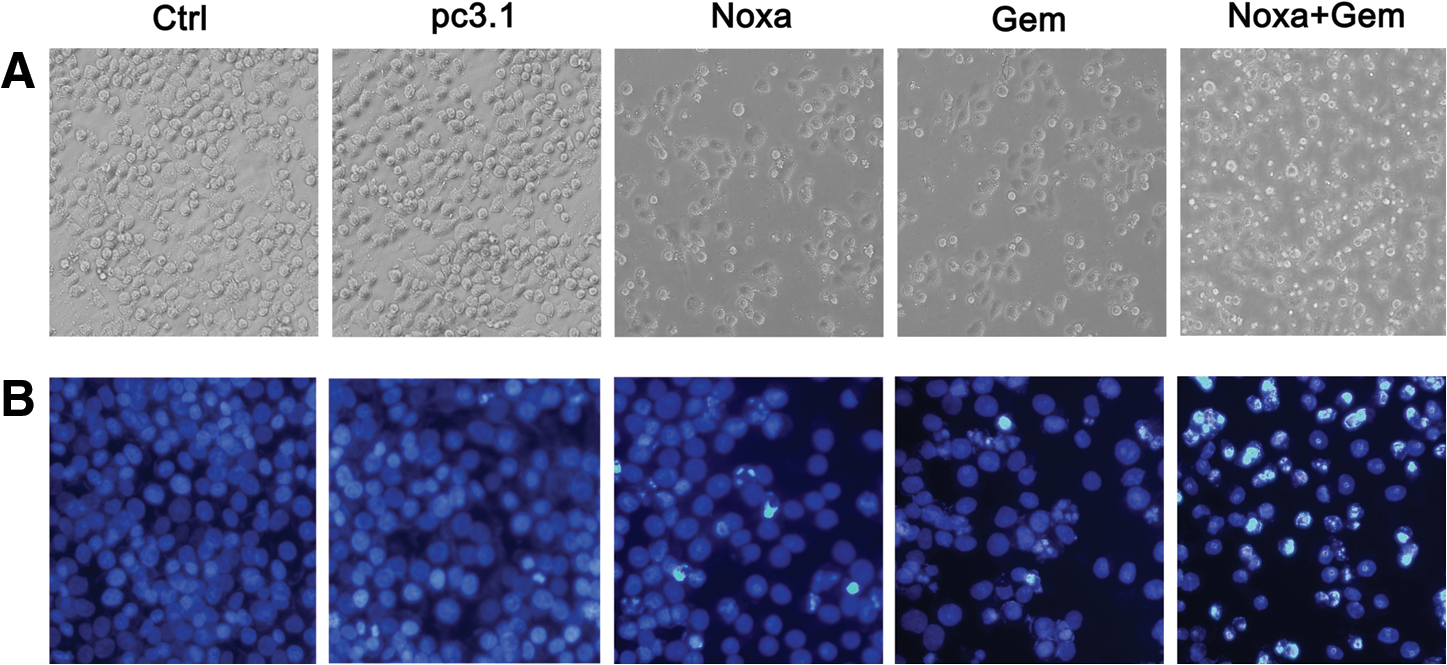
Cytotoxic effects of Noxa, gemcitabine, or a combination of both.
A quantitative assessment of sub-G1 cells by flow cytometry was used to estimate the number of apoptotic A2780 cells, as shown in Figure 4. The percentage of sub-G1 cells was significantly higher in the Noxa and gemcitabine groups than in two control groups (p<0.01). Moreover, the percentage of sub-G1 cells was significantly higher in the combination group than in the gemcitabine group (p<0.01). These data suggest that the cytotoxic effect of gemcitabine was augmented by Noxa.

Noxa and gemcitabine inhibit the growth of A2780 cells via apoptosis induction.
By means of western blot analysis, cleaved caspase-9 and cleaved caspase-3 were observed in A2780 cells that had been treated with Noxa or gemcitabine. However, more cleaved caspase-3 or cleaved caspase-9 was observed in the cells that had received combined treatment with both Noxa and gemcitabine (Fig. 5). These data demonstrate that Noxa enhances the cytotoxic effect of gemcitabine through the activation of caspase-9 and caspase-3.
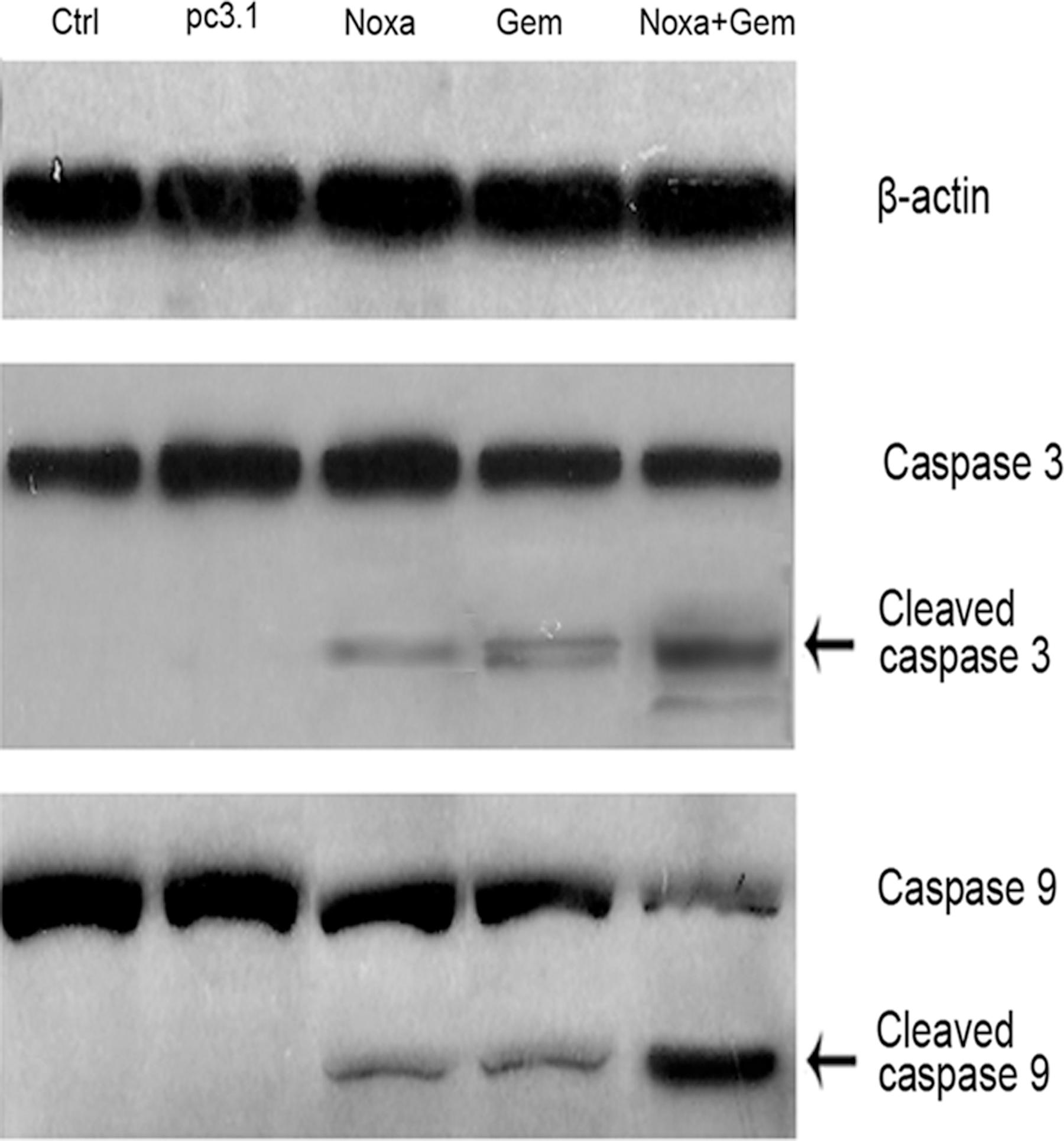
Western blot analysis for caspase-9 and caspase-3 in A2780 cells. Cleaved caspase-9 and cleaved caspase-3 were observed in A2780 cells treated with Noxa or with gemcitabine; a greater amount of cleaved caspase-3 and cleaved caspase-9 was found in the cells treated with a combination of Noxa and gemcitabine. β-actin expression served as a loading control.
The combined treatment with Noxa and gemcitabine inhibited tumor growth and prolonged the survival of nude mice
On the basis of the observed in vitro proapoptotic effects of Noxa and gemcitabine, we carried out a further investigation on the combined antineoplastic effect of Noxa and gemcitabine on xenograft tumors. Either A2780 cells or COC1 cells (5×106 cells) were inoculated s.c. into nude mice. The tumor-bearing nude mice were then either untreated (Control), treated with pcDNA3.1 (pc3.1) or Noxa (Noxa), treated with gemcitabine alone (Gem), or treated with Noxa and gemcitabine (Noxa+Gem). The tumors of the nude mice were then recorded in terms of their volume (Fig. 6A). Treatment with Noxa or gemcitabine singly resulted in a significant inhibition of tumor growth (p<0.01) in comparison with the two control groups. The combined treatment with Noxa and gemcitabine resulted in a significant inhibition of tumor growth in comparison with the gemcitabine group (p<0.05). However, it should be noted that no significant difference in tumor volume was observed between the gemcitabine-treated group and the Noxa-treated group. Survival curve analysis (Fig. 6B) showed that both Noxa and gemcitabine on their own resulted in a significant increase in life span compared with the two control groups (p<0.05). Survival of the tumor-bearing mice that received the combined treatment of Noxa and gemcitabine was further significantly prolonged compared with either the gemcitabine-treated group or the Noxa-treated group (p<0.05); there was no significant difference in survival time between the Noxa-treated mice and the gemcitabine-treated mice. These data suggest that Noxa may work co-operatively to enhance the antitumor efficacy of gemcitabine in the ovarian cancer xenograft model.
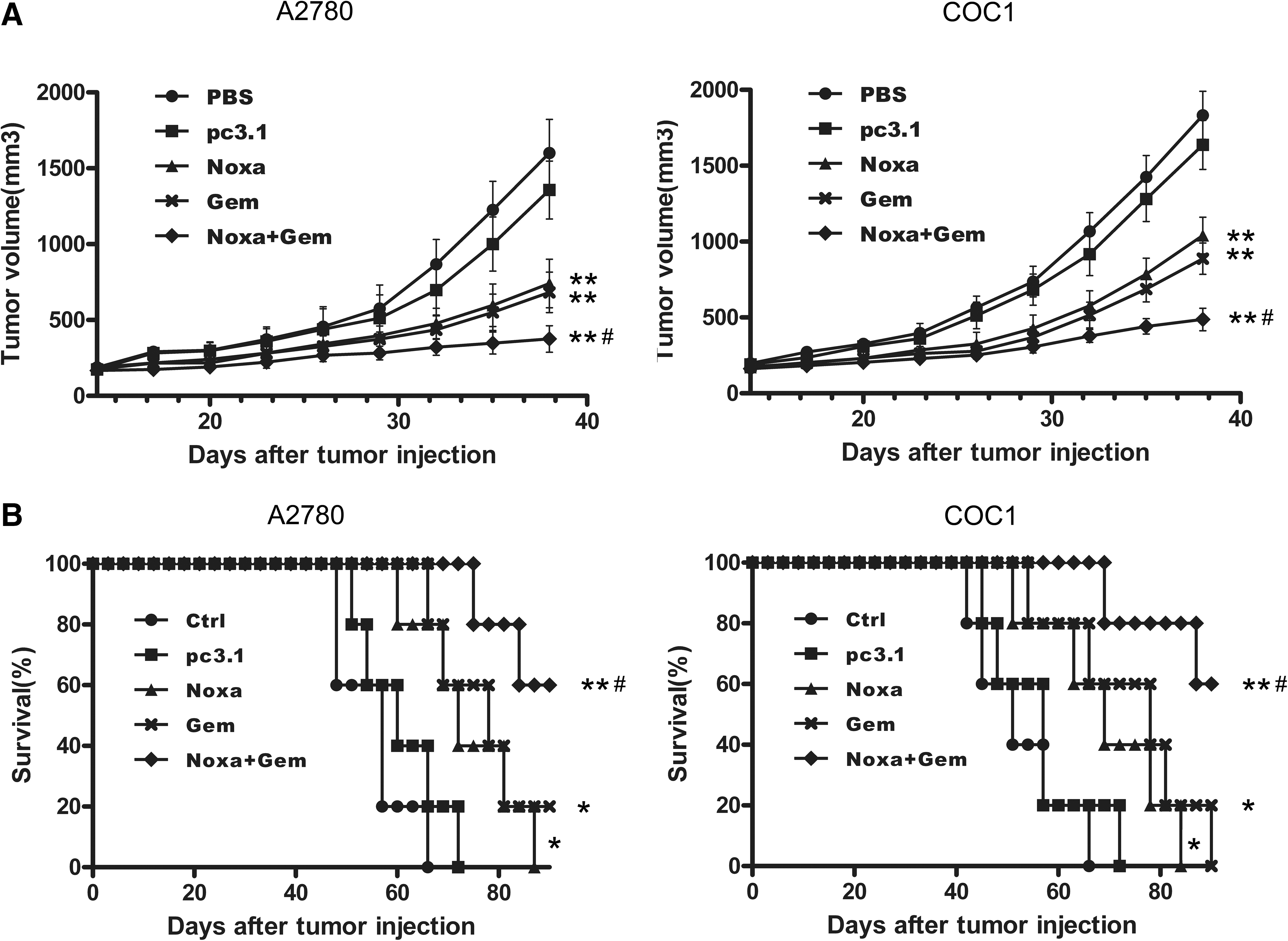
Tumor suppression and survival advantage found in xenograft models. Either A2780 or COC1 cells (5×106 cells) were inoculated s.c. into nude mice. Tumor-bearing nude mice were either untreated (Ctrl), or treated with pcDNA3.1 (pc3.1), with Noxa (Noxa), with gemcitabine alone (Gem), or with Noxa and gemcitabine (Noxa+Gem), respectively. Tumor volume
The treatment with Noxa and gemcitabine inhibited the growth of human ovarian cancer xenografts through the inhibition of proliferation and the induction of apoptosis
Tumor tissues of the tumor-bearing nude mice were excised and stained with H&E. Histologically, the tumors of the Noxa-plus-gemcitabine-treated mice caused visible responses, with larger necrosis/apoptosis regions, compared with the tumors of those treated with gemcitabine (data not shown). Immunohistochemistry of PCNA and TUNEL staining was used, respectively, as markers for proliferation and apoptosis. Representative sections of A2780 tumor tissues from different treatment groups were depicted (Fig. 7A).
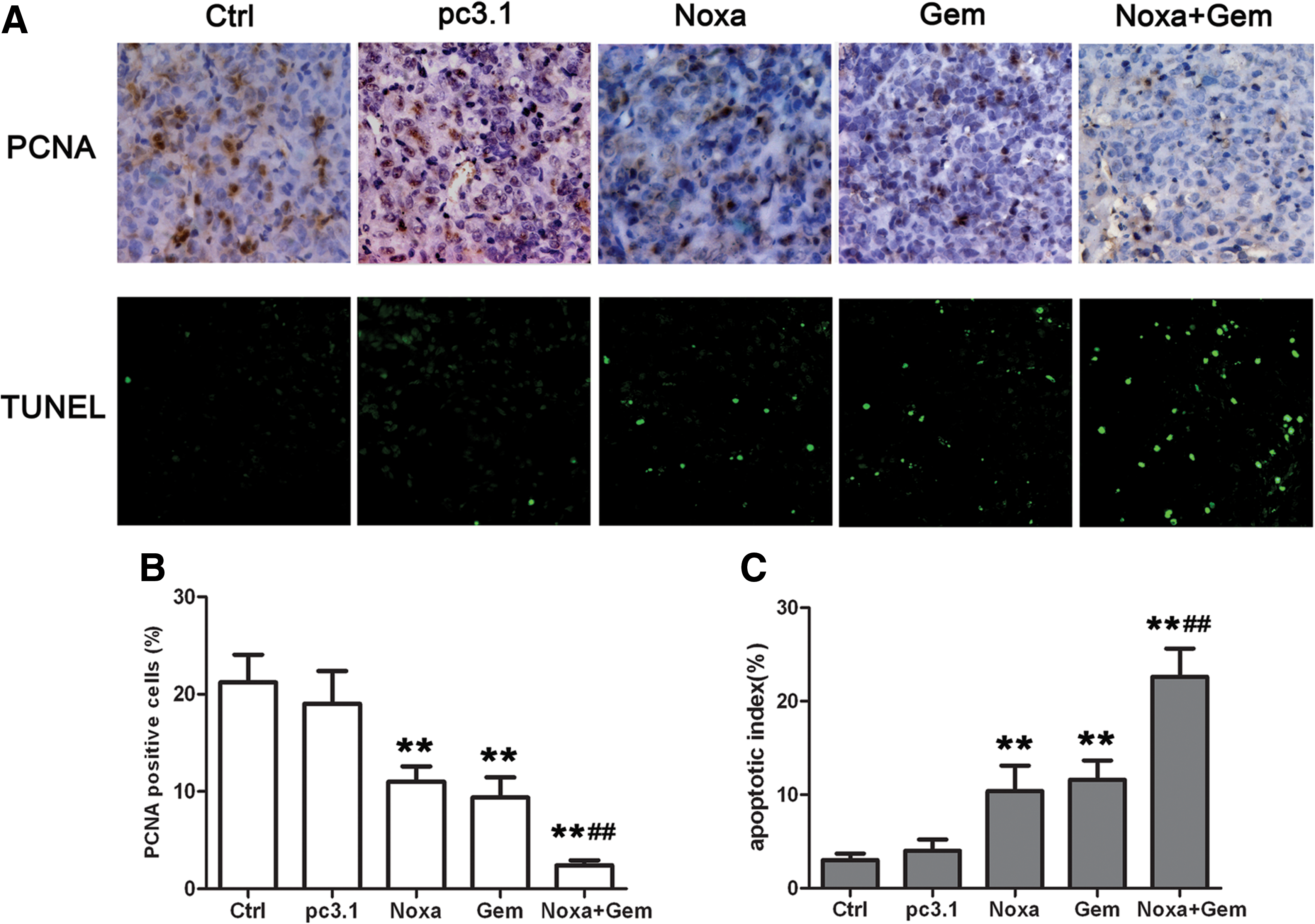
Noxa and gemcitabine inhibited the growth of tumor xenografts through the inhibition of proliferation and the induction of apoptosis.
As shown in Figure 7B, the number of PCNA-positive cells in five random fields was counted. Treatment with Noxa or gemcitabine resulted in significant proliferation inhibition (p<0.01) in comparison with the two control groups. The combined treatment with Noxa and gemcitabine showed a more significant antiproliferation effect in comparison with the gemcitabine group (p<0.01). These results demonstrate that the antiproliferation effect of gemcitabine was augmented when used in combination with Noxa.
TUNEL assay showed that both Noxa and gemcitabine, when singly used, increased the apoptotic rate of tumor cells compared with the two control groups. The number of apoptotic cells in five random fields was counted, and the apoptotic index is shown in Figure 7C. The results suggest that treatment with either Noxa or gemcitabine induced significant apoptosis (p<0.01) in comparison with the two control groups. Moreover, the combined treatment involving Noxa and gemcitabine showed a more significant proapoptotic effect in comparison with the gemcitabine group (p<0.01).
Observation of side-effects
To evaluate the health status of the mice that received the treatments, their weight was recorded once in every 3 days during the course of the experiment. No obvious adverse effects, such as weight loss, loss of appetite, aberrant behavior, or skin ulcerations, were observed in the five groups of mice (data not shown). Furthermore, no toxic pathologic changes in the heart, kidney, spleen, lungs, or liver were detected under microscopic examination (data not shown).
Discussion
Ovarian cancer is the number one cause of death from gynecologic malignancy. Gemcitabine is widely used in ovarian cancer therapy. Although gemcitabine is generally well tolerated, its most common side-effects occurring in patients take the form of myelosuppression, mildly increased hepatic transaminases, lethargy, fever, cutaneous hyperpigmentation, infusion-related maculopapular rash, and radiation recall dermatitis. 24 Both dose-dependent toxicity and the development of drug resistance greatly limit its therapeutic efficacy. Therefore, more efficient molecules and novel strategies are needed to enhance the efficacy of gemcitabine in ovarian cancer therapy.
Gene therapy has recently become a promising new therapeutic option in the treatment of cancer. Progress in the field of cancer gene therapy has advanced significantly, as evidenced by the five products that are now in final phase III clinical development. 25 For example, a form of Ad p53 product has been approved for cancer treatment in China. More than 3000 patients have been treated with gene therapy. 26 Given the continuous progress that has been made in the application of gene therapy for treating cancer, scientists have tried to use it to sensitize cancer cells to chemotherapeutic drugs or radiation to achieve a superior therapeutic effect.
Several kinds of viruses have been used as the vehicles in gene therapy, such as adenoviruses, herpes simplex virus, and new castle disease virus. However, there have been problems associated with their use. Viral vectors are now suspected of integrating with the human genome, as well as of altering metabolic pathways and inducing immunological responses to the virus and/or its gene products. 27 In this regard, scientists have predicted that the future of cancer gene therapy is in nonviral vectors. Nonviral vectors have some advantages over viral vectors. Nonviral delivery systems have fewer adverse immune responses, are easier to manufacture, and can be produced for the pharmaceutical industry in large quantities. 26 In this study, we used cationic liposome-DNA, which is a form of a nonviral delivery system, in ovarian cancer therapy. We have shown that the pcDNA3.1-Noxa plasmid coated with cationic liposome induced apoptosis and enhanced the antitumor effect of gemcitabine on A2780 ovarian cancer cells both in vitro and in vivo.
We carried out several experiments in this study to detect the induction of apoptosis. The in vitro experiments showed that both Noxa and gemcitabine inhibited the proliferation of A2780 cells through the induction of apoptosis, and they further showed that the antiproliferation effects were augmented when Noxa and gemcitabine were combined, as demonstrated by MTT assay (Fig. 2), Hoechst staining (Fig. 3B), and flow cytometric analysis (Fig. 4). In tumor xenograft experiments, we showed that Noxa could enhance the antineoplastic effect of gemcitabine through the enhanced inhibition of proliferation and the induction of apoptosis, as evidenced by immunohistochemical anti-PCNA staining (Fig. 7) and TUNEL assay (Fig. 7).
The p53 pathway plays an important role in gemcitabine-induced apoptosis, 28 and p53, caspase-9, and caspase-3 can be activated by gemcitabine treatment. 29 Since Noxa is an important proapoptotic protein in the p53 pathway, the overexpression of Noxa could reinforce the p53 pathway of apoptosis. And so, the antitumor effect of gemcitabine is enhanced. The induction of the Noxa gene can trigger the activation of proapoptotic BAK and BAX, thereby making the mitochondrial outer membrane permeable, allowing it to release cytochrome C. 16 Cytochrome C then activates caspase-9, which, in turn, activates the executioner caspases. In this study, we observed cleaved caspase-9 and cleaved caspase-3 in the A2780 cells that been treated with Noxa, and found more activated caspase-9 and caspase-3 in the cells that had been treated with a combination of Noxa and gemcitabine (Fig. 5). These results were identical to those of previous reports.
It has been reported by Suzuki et al. 20 that Noxa showed high efficacy against human breast cancer in nude mice. In this study, we found that overexpression of Noxa induced apoptosis in A2780 ovarian cancer cells, and that Noxa could also enhance the antitumor effect of gemcitabine. It is well known that p53 is one of the most widely used agents in current cancer gene therapy. As a transcriptional target of p53 and a potent inducer of apoptosis in various cancer cells, such as the ovarian variety, Noxa may be considered another important target that should be focused on in cancer gene therapy research in the future.
In conclusion, our data suggest that overexpression of Noxa can induce apoptosis in ovarian cancer cells, and that an elevated expression of Noxa can enhance the therapeutic responses of ovarian cancer cells to gemcitabine, both in vitro and in vivo. To our knowledge, we have provided the first evidence demonstrating the potential application of Noxa as a chemosensitizer of gemcitabine in the treatment of ovarian cancer.
Footnotes
Acknowledgments
This work was supported by “A Project of Scientific Research Fund of SiChuan Provincial Education Department (11ZB170).” The authors gratefully acknowledge the technical assistance rendered by Bing Kan and Yong-qiu Mao.
Disclosure Statement
No competing financial interests exist.